Risposte e breve discussione
1)
2)
b. L'acidità di una soluzione acquosa è espressa come pH ed è uguale al logaritmo negativo
in base 10 della concentrazione degli ioni idrogeno ([H+]). Nell'acqua pura vi è una
concentrazione di H+ pari a 10-7 M ed il pH = -log 10-7 = -(-7) = 7,0
c. Il pH di una soluzione è la misura della sua concentrazione idrogenionica. Per un acido
debole l'equilibrio di dissociazione è descritto da HA ⇔ H+ + A- ed il pKa è definito come
pKa = log 1/Ka
Ka è la costante di dissociazione acida ed è pari a:
[H+] [A-]
Ka = ————
[HA]
Si può dimostrare che il pKa di un acido è quel valore di pH al quale l'acido è dissociato per
metà e quindi, [A-] = [HA]. La relazione tra pH ed il rapporto di concentrazioni tra acido e
base coniugata è data dall'equazione di Henderson-Hasselbalch:
pH = pKa + log [base]/[acido]
Quindi per una soluzione contenente acido acetico 0,2 M e ione acetato 2M, sapendo che il
pKa dell'acido acetico è 4,8;
pH = 4,8 + log 2/0,2
= 4,8 + log 10
= 4,8 + 1
= 5,8
3)
4)
5)
6)
a. La maggior parte degli amminoacidi in soluzione a pH 7 sono in forma di doppio ione, il
cosiddetto "zwitterione". Il gruppo amminico legato al carbonio α è protonato (NH3 +) e
quello carbossilico è ionizzato (COO-). Solo cinque dei venti amminoacidi che fanno parte
delle proteine possiedono anche un gruppo ionizzabile nella catena laterale. Questi, pur
presentando una carica elettrica addizionale, rappresentano una minoranza delle molecole.
a. Arginina, lisina e istidina possiedono nella catena laterale dei gruppi chimici che in
soluzione acquosa si comportano come basi, acquistando idrogenioni dal solvente. L'acido
aspartico si dissocia invece come acido cedendo idrogenioni al solvente. La leucina possiede
una catena laterale idrocarburica ramificata e quindi non va incontro a dissociazione.
b. A parte la treonina che possiede un gruppo ossidrile nella catena laterale, gli altri
amminoacidi contengono zolfo. Tra questi soltanto l'omocisteina non si ritrova nella
costituzione delle proteine. E' invece un metabolita intermedio nella formazione della
cisteina a partire dalla metionina. Omocisteina e metionina sono anche intermedi del ciclo di
rigenerazione dell'S-adenosilmetionina, la molecola di grande importanza fisiologica
implicata nelle reazioni di metilazione. La metionina è un amminoacido essenziale che deve
essere introdotto con la dieta.
c. Il legame peptidico è il legame che si genera tra l'atomo di carbonio carbossilico di un
amminoacido e quello di azoto amminico di un secondo amminoacido. E' un legame rigido e
planare, non possiede libertà di rotazione e l'atomo di ossigeno carbonilico e quello di
idrogeno amminico sono sempre in configurazione trans (opposta). A causa di questa
conformazione, tuttavia, i legami formati dagli atomi di carbonio α dei residui
amminoacidici che partecipano alla formazione del legame peptidico possiedono una grande
libertà rotazionale.
7)
8)
9)
10)
11)
12)
13)
b. La struttura primaria di una proteina consiste semplicemente nella sua sequenza di
amminoacidi. Solo quei procedimenti che perseguano la determinazione della sequenza
amminoacidica hanno quindi rilievo a questo proposito. Ad esempio, hanno importanza la
separazione delle catene polipeptidiche e dei frammenti originati dalla rottura proteolitica
della catena polipeptidica. A tale proposito, hanno anche importanza la sequenziazione dei
frammenti ottenuti dalla digestione della catena polipeptidica originaria ed il confronto per
sovrapposizione delle sequenze dei frammenti peptidici ottenuti in condizioni sperimentali
differenti. α-eliche e foglietti- β sono invece esempi di struttura secondaria.
b. La struttura ad α-elica è un tipo comune di struttura secondaria delle proteine. Questa
consiste in un ripiegamento a spirale dello scheletro covalente polipeptidico con le catene
laterali degli amminoacidi che si estendono verso l'esterno, perpendicolarmente all'asse
dell'elica. Ogni atomo di ossigeno carbonilico del legame peptidico forma un ponte a
idrogeno con un legame peptidico posto a quattro residui di distanza nella catena
polipeptidica. Questo determina un numero di 3,6 amminoacidi per giro di elica. Poiché l'αelica è spesso presente all'interno delle proteine, essa è formata preferenzialmente da
amminoacidi apolari. Al contrario, gli amminoacidi con catene laterali ionizzabili o
ingombranti interferiscono con la formazione della struttura ad α-elica. La prolina e
l'idrossiprolina sono totalmente incompatibili con la spirale destrorsa dell'α-elica. L'alanina
è tra gli amminoacidi che possiedono la maggior propensione ad essere inseriti in una
struttura ad α-elica.
d. L'elettroforesi su gel di poliacrilammide in presenza di SDS viene comunemente
impiegata per separare le proteine in base alla loro massa. Con questa tecnica, le proteine
sono disciolte in una soluzione del detergente dodecilsolfato di sodio (SDS). L'SDS è un
detergente ionico che distrugge le interazioni non covalenti delle proteine native legandosi a
queste in un rapporto di una molecola di SDS ogni due residui amminoacidici. Ciò fa si che
la proteina denaturata possieda una quantità di carica negativa che è approssimativamente
proporzionale alla sua massa. La carica elettrica originaria della proteina non gioca quindi
alcun ruolo nella corsa elettroforetica. L'applicazione di una corrente elettrica alle estremità
del gel determina una migrazione più rapida per le proteine più piccole mentre quelle più
voluminose rimangono vicine al punto di applicazione.
e. Il termine struttura quaternaria si riferisce all'organizzazione strutturale di proteine
costituite da più catene polipeptidiche dette subunità. Solo alcune proteine possiedono
questo livello di organizzazione strutturale.
b. La sequenza e la composizione delle fibre di collageno sono uniche. Circa un trezo degli
amminoacidi è rappresentato dalla glicina. Prolina e idrossiprolina, o una combinazione
delle due, si trovano spesso comprese tra i residui di glicina che si ripetono nella sequenza.
Questi due amminoacidi costituiscono circa il 20 % del collageno. Si pensa che i residui di
prolina siano determinanti per la formazione dell'elica del collageno mentre quelli di glicina
ne permettano il superavvolgimento. I residui di prolina e, in minor proporzione, quelli di
lisina possono essere idrossilati in presenza di acido ascorbico (vitamina C), un agente
riducente. Idrossiprolina e idrossilisina sono perciò residui caratteristici del collageno e sono
raramente presenti in altre proteine. L'isoleucina è invece presente in tutte le proteine.
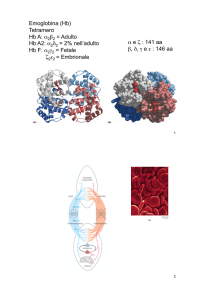
e. L'emoglobina dell'adulto, o emoglobina A, è costituita da quattro catene polipeptidiche, di
cui due α e due β. Le catene sono tenute assieme da interazioni non covalenti. Il tetramero di
emoglobina può essere correttamente rappresentato dall'associazione di due dimeri, ognuno
dei quali contiene le due diverse catene polipeptidiche. Quindi la schematizzazione (α 1 -β1 )
(α 2-β2 ), che si riferisce rispettivamente ai dimeri 1 e 2, è il modo migliore per rappresentare
la struttura quaternaria dell'emoglobina dell'adulto. Si pensa che le interazioni idrofobiche
siano le interazioni non covalenti più importanti per la stabilizzazione della struttura
quaternaria.
b. Nella curva di dissociazione dell'ossigeno si riporta la percentuale di saturazione
dell'emoglobina con l'ossigeno in funzione della quantità di ossigeno presente in soluzione,
14)
15)
16)
17)
18)
19)
20)
espressa come pO2 (pressione parziale di ossigeno). La saturazione dell'emoglobina varia da
0 a 100 %. La pO2 può variare dall'assenza di ossigeno in soluzione fino a valori più alti. Il
2,3-bifofoglicerato, o BPG, è presente all'interno dei globuli rossi in concentrazioni simili a
quelle dell'emoglobina. Il BPG si lega all'emoglobina non ossigenata abbassandone l'affinità
per l'ossigeno e favorendo quindi il rilascio di ossigeno a livello dei capillari. Ne consegue
che l'aumento dei livelli di BPG sposta la curva di dissociazione dell'ossigeno verso destra.
Allo stesso modo, l'aumento degli ioni H+ e dei livelli di CO2 inducono il rilascio dell'O2
dall'emoglobina, spostando la curva verso destra. Viceversa, l'abbassamento dei livelli di H+
e della CO2 spostano la curva verso sinistra. L'emoglobina fetale ha una maggior affinità per
l'ossigeno in tutte le condizioni. Il mescolamento di emoglobina fetale con quella di adulto
comporta un aumento dell'affinità per l'O2 con uno spostamento della curva verso sinistra.
d. L'emoglobina è una proteina tetramerica la cui curva di saturazione dell'ossigeno ha un
caratteristico andamento sigmoidale. Ciò è conseguenza delle interazioni cooperative tra i
quattro siti di legame. L'ossigeno si lega all'emoglobina senza che si abbiano cambiamenti
dello stato di ossidazione dello ione ferroso presente nell'eme. Il monossido di carbonio e il
cianuro si legano all'emoglobina con maggiore affinità rispetto all'ossigeno. L'O2 è rilasciato
ai tessuti e scambiato con la CO2 in quanto l'aumento dei livelli di CO2 nei capillari
comporta una diminuzione dell'affinità dell'emoglobina per l'ossigeno.
e. Gli enzimi accelerano le reazioni diminuendone l'energia libera di attivazione ovvero
abbassandone la barriera di attivazione. Attraverso la combinazione con il substrato, un
enzima altera il meccanismo della reazione favorendo la formazione di uno stato di
transizione la cui energia è inferiore a quella dello stato di transizione della reazione non
catalizzata. In questo modo, gli enzimi accelerano le reazioni fino a milioni di volte. I livelli
energetici dei reagenti non sono influenzati dalla presenza degli enzimi. Gli enzimi non
modificano gli equilibri delle reazioni.
d. Gli isoenzimi, forme diverse dello stesso enzima, sono costituiti dall'associazione di
subunità diverse in varie proporzioni. Ad esempio, la lattato deidrogenasi è un tetramero di
quattro subunità. Le quattro subunità possono essere uguali tra loro ovvero essere una
miscela di due tipi di subunità (H e M). Le combinazioni possibili delle due subunità
possono originare cinque differenti isoenzimi: H4 , H3 M, H2 M2 , HM3 , M4 . Questi hanno
simile specificità di substrato ma diverse affinità e mobilità elettroforetica. Il tipo H è
presente soprattutto nel muscolo cardiaco mentre il tipo M è più abbondante nel fegato e nel
muscolo scheletrico.
b. La Km, costante di Michaelis-Menten, è definita come la concentrazione di substrato in
presenza della quale la reazione catalizzata da un enzima procede ad una velocità pari a metà
della velocità massima.
c. Gli enzimi allosterici, a differenza degli enzimi comuni, non seguono la cinetica di
Michaelis-Menten. Un sito attivo di un enzima allosterico spesso influenza positivamente un
altro sito attivo della stessa molecola. Questo fenomeno, noto come cooperatività, determina
un andamento sigmoidale della curva di V in funzione di [S]. I termini inibizione competitiva
e non competitiva si riferiscono alle cinetiche di Michaelis-Menten ma non agli enzimi
allosterici.
c. Gli inibitori non competitivi, contrariamente agli inibitori competitivi, non sono analoghi
strutturali del substrato. Di conseguenza, gli inibitori non competitivi si legano agli enzimi
in posizioni distanti dal sito attivo. Per questo motivo, il grado di inibizione è legato
esclusivamente alla concentrazione di inibitore e l'aumento della concentrazione del
substrato non potrà rimuovere l'inibizione. In questo caso quindi avremo una diminuzione
della Vmax senza che la Km sia apprezzabilmente modificata. Nel caso dell'inibizione
competitiva avremo invece un sensibile aumento del valore della Km.
a. Il legame di un effettore alla subunità regolatrice di un enzima allosterico causa un
cambiamento conformazionale che può comportare un aumento o una diminuzione
dell'attività enzimatica. La cooperatività positiva è propria di alcune molecole, ad esempio
21)
22)
l'emoglobina. Un effettore positivo aumenta l'affinità per il substrato. Questo è il caso della
protein chinasi AMP ciclico -dipendente. L'AMP ciclico si lega alla subunità regolatrice che,
di conseguenza, si dissocia dalla subunità catalitica, attivandola. In assenza di AMP ciclico,
la subunità regolatrice è saldamente legata alla subunità catalitica rendendo l'enzima
inattivo. Molti enzimi allosterici catalizzano spesso la prima tappa, ovvero la tappa
regolatrice, di una via metabolica. Il prodotto finale della via agisce, quindi, come effettore
negativo dell'enzima. Questo fenomeno è noto come inibizione retrograda (inibizione a
feedback).
b. Gli enzimi possono essere regolati a lungo o a breve termine. La regolazione a breve
termine, in genere rapida, avviene attraverso inibizione da prodotto terminale, controllo
allosterico, disponibilità di substrato e modificazioni covalenti. In questo tipo di regolazione
il prodotto, i substrati o un altro enzima regolatore modificheranno la Vmax o la Km
dell'enzima in modo rapido. Al contrario, la regolazione a lungo termine, che avviene nel
giro di ore o addirittura giorni, è effettuata attraverso un aumento della velocità di
degradazione o della velocità di sintesi di un enzima. I cambiamenti dei livelli quantitativi di
enzima che conseguono a tali meccanismi sono in genere frutto della presenza di ormoni o
metaboliti che svolgono azioni di repressione o derepressione genica.
d. La pepsina è secreta dalle cellule della mucosa gastrica in forma di precursore. A
differenza della maggior parte dei proenzimi digestivi, questa non è attivata da un'idrolisi
catalizzata da proteasi. Il pepsinogeno è convertito a pepsina mediante idrolisi acida
spontanea a pH 2. L'acido cloridrico secreto dalle cellule gastriche è responsabile di questa
acidità. Tutti gli enzimi pancreatici sono attivati contemporaneamente non appena riversati
nel duodeno. Ciò avviene per idrolisi del tripsinogeno, chimotripsinogeno, proelastasi e
procarbossipeptidasi da parte della tripsina stessa. Quantità catalitiche di tripsina originano
inizialmente dall'azione dell'enteropeptidasi duodenale sul tripsinogeno.