IL METABOLISMO ENERGETICO
BIOCHIMICA. GLICOLISI, FERMENTAZIONE E RESPIRAZIONE CELLULARE
© GSCATULLO
(
Metabolismo energetico
Introduzione
Funzione
Il metabolismo energetico è un’attività coordinata cui partecipano milioni di reazioni che svolgono tre
funzioni principali:
Ricavare energia mediante la trasformazione di energia luminosa in energia chimica, è quanto
avviene negli autotrofi (piante e vegetali); o demolendo molecole di nutrienti, negli eterotrofi.
Idrolizzare i polimeri biologici (polisaccaridi, proteine, acidi nucleici) in monomeri (monosaccaridi,
amminoacidi, nucleotidi), e trasformare i trigliceridi in glicerolo e a acidi grassi.
Sintetizzare i polisaccaridi, le proteine e gli acidi nucleici a partire dai monomeri che li costituiscono,
e i trigliceridi a partire da glicerolo e acidi grassi.
Queste trasformazioni si svolgono grazie ad una serie di enzimi, che sono presenti ognuno in una diversa
tappa e catalizzano le reazioni che trasformano il substrato in prodotti intermedi, e quelli nel prodotto finale.
L’insieme delle reazioni coinvolte nello stesso processo metabolico si chiama via metabolica.
Numerose vie metaboliche sono comuni a tutti gli organismi, altre invece sono peculiari di alcuni gruppi. Negli
eucarioti molte vie metaboliche sono compartimentate, avvengono cioè all’interno di organuli specifici come
i mitocondri o i cloroplasti.
Le Redox
Sappiamo che le reazioni metaboliche si dividono in cataboliche, esoergoniche che producono energia libera
utile per la sintesi di ATP, e anaboliche, endoergoniche che utilizzano l’energia dell’ATP per i processi cellulari.
Nella cellula la maggior parte delle reazioni anaboliche e cataboliche sono reazioni di ossidoriduzione (o
reazioni redox), in cui uno o più elettroni vengono trasferiti da una specie chimica (un composto) ad un’altra.
Una reazione redox è costituita da due semireazioni:
La semireazione di ossidazione (OX), che consiste nella perdita di uno o più elettroni;
La semireazione di riduzione (RED) che consiste nell’acquisto di uno o più elettroni.
La specie chimica che perde elettroni è detta riducente, quella che li acquista è chiamata ossidante.
L’ossidazione e la riduzione avvengono sempre assieme: il trasferimento di elettroni richiede la presenza
contemporanea di un donatore e di un accettore di elettroni. Il riducente, che perde elettroni, si ossida,
mentre l’ossidante, che li acquista, si riduce.
Nei sistemi biologici le semireazioni di ossidazione e di riduzione avvengono in genere attraverso la perdita e
l’acquisto di atomi di idrogeno: il loro trasferimento infatti comporta anche il trasferimento di un elettrone.
𝐻 = 𝐻+ + 𝑒 −
Per questo motivo in biologia, ossidazione è sinonimo di deidrogenazione, così gli enzimi che catalizzano
reazioni di ossidazione, e che fanno parte della famiglia delle ossidoreduttasi, sono detti anche deidrogenasi.
Coenzimi
Le deidrogenasi cellulari catalizzano l’ossidazione di centinaia di composti diversi assieme a specifici coenzimi
chiamati NAD, NADP e FAD. Questi coenzimi possono andare incontro a ossidazioni e riduzioni reversibili,
comportandosi di fatto come “navette” di idrogeno (e quindi di elettroni) che passano da una via metabolica
ad un’altra.
Il NAD
Il nicotinammide-adenin-dinucleotide (NAD) e il nicotinammide-adenin-dinucleotide-fosfato (NADP) sono
coenzimi costituiti da due nucleotidi uniti da due gruppi fosfato. In genere il NAD opera nelle reazioni
cataboliche (es. ossidazione del glucosio), mentre il NADP in quelle anaboliche e nella protezione dai radicali
liberi prodotti dall’ossigeno.
Entrambi i coenzimi possono esistere in due forme distinte: quella ossidata e quella ridotta. Quando in una
via metabolica una molecola di substrato va incontro a ossidazione perdendo due atomi di idrogeno, essi
possono essere accettati dalla forma ossidata del coenzima (NAD+), che si trasforma in questo modo nella
forma ridotta (NADH). È importante questo processo perché in questo modo l’idrogeno (H), acido, non si
trova libero nella cellula, il che ne altererebbe il PH. La reazione avviene tra il NAD e una molecola di idrogeno
(2H).
𝑁𝐴𝐷 + + 2𝐻 → 𝑁𝐴𝐷𝐻 + 𝐻 +
All’opposto il NADH può essere ossidato grazie all’ossigeno: ciò comporta una riduzione dell’ossigeno
molecolare e la trasformazione del coenzima nella forma ossidata NAD+.
1
𝑁𝐴𝐷𝐻 + 𝐻 + → 𝑂2 → 𝑁𝐴𝐷 + + 𝐻2 𝑂
2
Questa reazione è fortemente esoergonica e porta ad una variazione di energia libera di -52,4 kcal/mole. Ciò
significa che il NADH trasporta una grande quantità di energia: basti pensare che l’ATP ne trasporta solo 7,3
kcal/mole. Il NAD deriva dalla vitamina B, o niacina, che è sintetizzata a sua volta a partire dall’amminoacido
triptofano.
Il FAD
Il flavin-adenin-dinucleotide (FAD) è il coenzima tipico delle flavoproteine, una classe di enzimi che catalizza
reazioni redox. Anche il FAD si presenta in una forma ossidata (FAD) ed in una ridotta (FADH2). La forma
ossidata può accettare due atomi di idrogeno e trasformarsi in quella ridotta. Questo coenzima è coinvolto
nelle reazioni cataboliche dell’ossidazione del glucosio e deriva dalla vitamina B2 o riboflavina, presente nel
fegato e nei muscoli.
𝐹𝐴𝐷 + 2𝐻 → 𝐹𝐴𝐷𝐻2
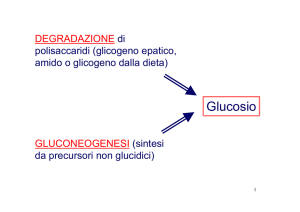
Il catabolismo del glucosio
Alcune vie metaboliche sono comuni a tutti gli organismi, in particolare il metabolismo del glucosio ricopre
un ruolo centrale per le piante, gli animali e molti microrganismi eucarioti e procarioti. Questo esoso è ricco
di energia potenziale che viene liberata grazie alla combustione (catabolismo), questo processo avviene nelle
cellule: i legami del glucosio vengono rotti e liberano l’energia che li formavano, questa energia viene
immagazzinata nelle molecole di ATP e può essere trasportata per tutta la cellula.
Questa estrazione di energia dai legami di glucosio (catabolismo del glucosio) avviene tramite tre grandi
processi catabolici:
La glicolisi, via iniziale del catabolismo del glucosio, che lo converte con una serie di reazioni in una
molecola a tre atomi di carbonio, il piruvato. L’energia che si è liberata è accumulata sotto forma di
ATP e NADH. La glicolisi è un processo anaerobico perché non richiede ossigeno: costituisce per alcuni
organi, come il cervello, la sola fonte di energia!
In presenza di ossigeno alla glicolisi segue la respirazione cellulare, che ossida ulteriormente il
piruvato formando acqua e anidride carbonica. Questo processo libera una grande quantità di
energia immagazzinata nell’ATP, per avvenire richiede ossigeno, per tanto è un processo aerobico.
In assenza di ossigeno alla glicolisi segue la fermentazione: un processo anaerobico come la glicolisi,
che consiste nella trasformazione del piruvato in lattato o etanolo. Questo processo libera un’energia
assai minore rispetto la respirazione cellulare.
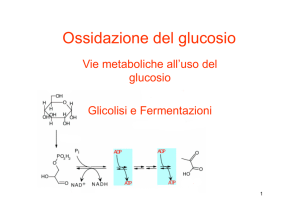
La glicolisi
La glicolisi si svolge nel citoplasma di tutte le cellule e comprende dieci reazioni, ognuna catalizzata da uno
specifico enzima. Durante le reazioni della glicolisi si verifica l’ossidazione incompleta della molecola di
glucosio con formazione di due molecole di piruvato e liberazione di energia chimica: l’energia liberata è
impiegata per la sintesi di ATP e per la riduzione di NAD+.
Le dieci tappe del processo si possono raggruppare in due grandi fasi di cinque ciascuna:
La fase endoergonica, in cui il glucosio (6 atomi di C) viene scisso tramite una serie di reazioni in due
molecole a 3 atomi di C. Questa fase è detta endoergonica perché è necessario fornire energia, sotto
forma di due molecole di ATP, per avviarla. Ogni ATP fornisce al glucosio un gruppo fosfato (processo
che prende nome di fosforilazione) che rende la molecola più instabile e reattiva e le impedisce di
uscire dalla membrana plasmatica della cellula, grazie alla carica negativa.
La fase esoergonica in cui le due molecole di gliceraldeide 3-fosfato (G3P, una ottenuta come
isomero: diidrossiacetone fosfato), vengono trasformate in piruvato sciogliendone i legami esteri con
il fosfato. I gruppi fosfato vengono accettati da molecole di ATP. Si ottengono alla fine del processo:
2 molecole di piruvato, 4 ATP e 2 NADH. La seconda fase deve essere considerata “doppia” per ogni
tappa in quanto il processo si ripete per ognuna delle due molecole di G3P.
La reazione della glicolisi ha per bilancio:
𝑔𝑙𝑢𝑐𝑜𝑠𝑖𝑜 + 2𝐴𝑇𝑃 + 2𝑁𝐴𝐷 + + 4𝐴𝐷𝑃 + 2𝑃𝑖 → 2 𝑝𝑖𝑟𝑢𝑣𝑎𝑡𝑜 + 2 𝐴𝐷𝑃 + 2𝑁𝐴𝐷𝐻 + 2𝐻 + + 2𝐴𝑇𝑃 + 2𝐻2 𝑂
Semplificando i termini comuni, otteniamo
𝑔𝑙𝑢𝑐𝑜𝑠𝑖𝑜 + 2𝑁𝐴𝐷 + + 2𝐴𝐷𝑃 + 2𝑃𝑖 → 2 𝑝𝑖𝑟𝑢𝑣𝑎𝑡𝑜 + 2𝑁𝐴𝐷𝐻 + 2𝐻 + + 2𝐴𝑇𝑃 + 2 𝐻2 𝑂
La fermentazione
L’obbiettivo della fermentazione è ritrasformare il NADH prodotto nella glicolisi (2NADH) in 2NAD++2H
riossidandolo in assenza di ossigeno (via anaerobica): in assenza di NAD+ infatti il processo di glicolisi si
bloccherebbe e la cellula morirebbe per mancanza di energia! Il substrato che le cellule usano per riossidare
il NADH per via anaerobica è ovviamente il piruvato, prodotto della glicolisi.
La fermentazione lattica
In alcune condizioni le cellule non dispongono di ossigeno sufficiente per riossidare il NADH prodotto dalla
glicolisi attraverso la respirazione cellulare, per rigenerare il NAD+ usano allora un processo detto
fermentazione lattica, che riduce il piruvato al lattato per mezzo dell’enzima lattato deidrogenasi.
Il legame centrale del carbonio con l’ossigeno “si apre” per legarsi ai due idrogeni del NADH + H. Questa
fermentazione avviene tipicamente nelle cellule muscolari durante un’attività intensa: il circolo sanguineo
non fornisce abbastanza ossigeno e le cellule non riescono ad ossidare il piruvato e il NADH prodotto dalla
glicolisi. Il muscolo utilizza le riserve di glucosio per produrre ATP: la glicolisi ne produce 2 molecole e il
piruvato viene smaltito in lattato con la fermentazione. Quando i livelli di lattato nel sangue sono alti i tessuti
possono acidificarsi e si avverte lo sforzo: il lattato deve allora essere ritrasformato in glucosio dal fegato.
Questo processo prende nome di ciclo di Cori, dal nome degli studiosi che chiarirono questa via metabolica
e la sua funzione. La fermentazione lattica è impiegata anche da batteri come Lactobacillus casei, che si trova
nell’intestino umano ma usato anche per produrre lo yogurt.
La fermentazione alcolica
Il lievito e altri microorganismi riossidano il NADH in NAD+ attraverso un processo chiamato fermentazione
alcolica. È suddiviso in due fasi: la prima il piruvato è decarbossilato, perde cioè un carbonio che viene
trasformato in CO2, grazie all’enzima piruvato decarbossilasi e trasformato in acetaldeide; la seconda fase
avviene con l’alcol deidrogenasi e prevede l’ossidazione del NADH + H in NAD+ e trasformando l’acetaldeide
in etanolo.
La respirazione cellulare
Per tutti gli organismi che non svolgono la fermentazione la glicolisi è solo la prima fase dell’ossidazione del
glucosio. In presenza di ossigeno il piruvato prodotto dalla glicolisi viene ulteriormente ossidato a H2O e CO2;
questa fase aerobica costituisce la respirazione cellulare.
La respirazione cellulare avviene nei mitocondri, organuli avvolti da una doppia membrana:
la membrana mitocondriale esterna, liscia e permeabile, a piccole molecole e a ioni, grazie a canali
transmembrana formati da proteine chiamate porine;
la membrana mitocondriale interna, estesa e ripiegata a formare le creste, impermeabile alle
molecole di piccole dimensioni e agli ioni.
Grazie alla presenza di due membrane i mitocondri risultano divisi in due compartimenti: lo spazio
intermembrana, posto tra la membrana esterna e quella interna, e la matrice mitocondriale, delimitata dalla
membrana interna.
La respirazione cellulare comprende tre vie metaboliche che avvengono in sequenza:
1. la decarbossilazione ossidativa del piruvato, nella matrice mitocondriale ossida il piruvato a gruppo
acetile;
2. il ciclo di Krebs, serie ciclica di reazioni che ossidano il gruppo acetile a CO2, anche questa nella
matrice mitocondriale;
3. la fosforilazione ossidativa, che riossida i coenzimi ridotti NADH e FADH2 liberando elettroni e
protoni, i primi sono trasferiti all’ossigeno tramite una catena di trasporto o catena respiratoria,
lungo il processo l’energia liberata è accumulata sotto forma di ATP.
La decarbossilazione del piruvato
Prima di entrare nel Ciclo di Krebs il piruvato deve essere trasformato in acetil-CoA, nei procarioti questa
conversione avviene nel citoplasma, mentre negli eucarioti il piruvato deve entrare nella matrice
mitocondriale grazie ad una proteina di trasporto. La reazione, catalizzata dalla piruvato deidrogenasi,
comprende due eventi:
1. la decarbossilazione e l’ossidazione del piruvato (3 atomi di C) ad acetile (2 atomi di C) con liberazione
di una molecola di CO2 e la riduzione di una molecola di NAD+ a NADH.
2. La formazione di un legame tra il gruppo acetile e il coenzima A (CoA) per produrre acetil-CoA.
Complessivamente, quindi, per ogni molecola di glucosio si otterranno due CO2, due acetil-CoA e due NADH.
Per il suo ruolo centrale nel collegamento tra la glicolisi e la respirazione cellulare, la piruvato deidrogenasi
costituisce un enzima chiave del nostro metabolismo e le sue alterazioni risultano essere molto pericolose.
Il Ciclo di Krebs
Il ciclo di Krebs o ciclo dell’acido citrico è una via metabolica costituita da otto reazioni, ciascuna catalizzata
da uno specifico enzima. Il ciclo inizia con il legame del gruppo acetile dell’acetil-CoA (2 atomi di C)
all’ossalacetato (4 C), per formare il citrato (6 C), questa reazione costruttiva (anabolica) necessita di energia
per essere attivata e si ricava dal legame con il CoA che viene perso. Il citrato diventa isocitrato (sempre 6 C)
grazie ad un enzima isomerasi, e l’isocitrato viene decarbossilato (CO2 + NADH + H+) per formare l’αchetoglutrato (5C). Quest’ultimo è ancora decarbossilato, diventa succinil-CoA (il CoA è uno dei coenzimi
utilizzati, ma lo perde), e quindi succinato. Il succinato è ossidato (FADH2) in fumerato, che con l’acqua
reagisce in malato, che è di nuovo ossidato (NADH + H+) per ottenere l’ossalacetato.
L’energia impiegata nel Ciclo di Krebs è impiegata per la riduzione di tre molecole di NAD+ e di una di FAD,
oltre che per la sintesi di una molecola di ATP. Questi risultati, se considerati per ogni molecola di glucosio
avviata alla glicolisi, devono essere raddoppiati: al ciclo di Krebs giungono due acetil-CoA, uno per ogni
piruvato decarbossilato. Nel complesso le reazioni del ciclo di Krebs portano all’ossidazione completa dei
due atomi di carboni del gruppo acetile.
2 𝑎𝑐𝑒𝑡𝑖𝑙𝐶𝑜𝐴 + 6𝑁𝐴𝐷 + + 2𝐹𝐴𝐷 + 2𝐴𝐷𝑃 + 2𝑃𝑖 + 4𝐻2 𝑂
→ 4𝐶𝑂2 + 6𝑁𝐴𝐷𝐻 + 6𝐻 + + 2𝐹𝐴𝐷𝐻2 + 2𝐴𝑇𝑃 + 2𝐶𝑜𝐴
La fosforilazione ossidativa
L’ossidazione del glucosio, del piruvato e del gruppo acetile producono molte molecole di NADH e FADH2:
queste molecole contengono un’elevata quantità di energia che viene usata nella fosforilazione ossidativa
per sintetizzare ATP.
Tale processo ha inizio con l’ingresso nella catena respiratoria mitocondriale, che si trova nella membrana
interna del mitocondrio, degli elettroni che si trovano nei coenzimi NAD e FAD (sotto forma di H). NADH e
FADH2 passano attraverso una serie di trasportatori di elettroni fino ad arrivare all’ossigeno con cui formano
una molecola d’acqua. Durante questo percorso gli elettroni prendono parte ad una serie di reazioni redox
rilasciando gradualmente la loro energia.
I trasportatori di energia hanno natura diversa, le flavoproteine, l’ubichinone o coenzima Q (ponte tra i
complessi), i citocromi (proteine), e le proteine ferro-zolfo; e sono organizzati in quattro grandi complessi
proteici di membrana distinti in:
Complesso I, NADH deidrogenasi che si occupa di trasferire due elettroni del NADH all’ubichinone, e
quattro protoni dalla matrice allo spazio intermembrana
Complesso II, succinato deidrogenasi trasferisce elettroni dal FADH2 all’ubichinone.
Complesso III, o ubichinone-citocromo c ossidoreduttasi
Complesso IV, o citocromo c ossidasi che trasferisce quattro elettroni dal citocromo C ad una mezza
mole di molecola di ossigeno per formare l’acqua, secondo la reazione:
𝑂2 + 4𝑒 − + 4𝐻 + → 2𝐻2 𝑂
La chemiosmosi
Il trasferimento di elettroni lungo la catena respiratoria è accompagnato da un trasferimento di protoni dalla
matrice verso lo spazio intermembrana, questo provoca uno scompenso chimico, per l’alta concentrazione
di H+ nello spazio intermembrana a differenza di quanto presente nella matrice, e elettrico per la differenza
di cariche.
Questo scompenso, o gradiente, elettrochimico costituisce una fonte di energia potenziale detta forza
proton-motrice. Poiché la membrana interna è impermeabile ai protoni per attraversarla devono passare
attraverso un complesso proteico chiamato ATP sintasi che: canalizza i protoni permettendo di diffondersi di
nuovo nella matrice e utilizza l’energia di questa diffusione per formare ATP a partire da ADP e Pi.
L’accoppiamento tra la forza proton-motrice e la sintesi dell’ATP è detto chemiosmosi e avviene in tutte le
cellule che respirano. La reazione che produce ATP è
𝐴𝑇𝑃 ⇋ 𝐴𝐷𝑃 + 𝑃𝑖 + 𝑒𝑛𝑒𝑟𝑔𝑖𝑎 𝑙𝑖𝑏𝑒𝑟𝑎
Bilancio energetico
Bilancio glicolisi e fermentazione: 𝐶6 𝐻12 𝑂6 → 2 𝑙𝑎𝑡𝑡𝑎𝑡𝑜 (𝑜 2 𝑒𝑡𝑎𝑛𝑜𝑙𝑜 + 2𝐶𝑂2 ) + 2𝐴𝑇𝑃
Bilancio glicolisi e respirazione cellulare: 𝐶6 𝐻12 𝑂6 + 6𝑂2 → 6𝐶𝑂2 + 6𝐻2 𝑂 + 32𝐴𝑇𝑃
Realizzato da Paolo Franchi, 5°BC A.S. 2015/2016 il 02/05/2016. AMDG





![10. Glicolisi [modalità compatibilità]](http://s1.studylibit.com/store/data/001348757_1-a541d29faa26ffecd55b52cbf7ba8f98-300x300.png)