Dr.ssa Mauro
ricevimento studenti
il lunedì dalle ore 12.00 alle ore 13.30
o previo appuntamento telefonico
0861.266888
E-mail: [email protected]
N.d.A.: Il materiale contenuto nel presente file è inteso ad
esclusivo utilizzo didattico da parte degli studenti del I°
anno di Biotecnologie, che sono invitati a non ridistribuire
suddetto materiale, parzialmente coperto da copyright.
MEMBRANA PLASMATICA
Sottile membrana di circa 6-8 nm che
avvolge la cellula. Non è visibile al MO
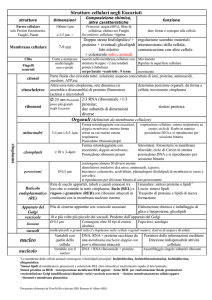
La membrana plasmatica o membrana cellulare o plasmalemma è un sottile
involucro che avvolge la cellula separandola dall’esterno e regolando lo
scambio di sostanze trofiche.
Evoluzione dei modelli di membrana
Struttura delle membrane biologiche
lipidi
proteine
per lo più fosfolipidi, glicolipidi
e steroli
integrali o intrinseche
estrinseche o periferiche
oligosaccaridi confinati alla sola
glucidi
superficie esterna dove svolgono
una funzione recettoriale e di
riconoscimento
La membrana è una struttura ASIMMETRICA
Film sottile di lipidi e proteine, tenute unite
da interazioni non covalenti
Modello di membrana a mosaico fluido
Fosfolipidi
Fosfato (-)
glicerolo
acidi grassi
saturi
insaturi
Testa
Polare
Coda
Apolare
I lipidi di membrana sono molecole anfipatiche: polo idrofilo e coda idrofoba.
Preciso orientamento spaziale nella costituzione di tutte le membrane
costituzione del doppio strato molecolare (bilayer lipidico)
Lipidi: 50% della massa della
membrana: circa 109 mol/cellula
animale.
Fosfolipidi:
a) fosfogliceridi derivati dal
glicerolo (fosfatidilcolina…)
b) sfingolipidi derivati dalla
sfingosina (sfingomielina…)
Colestero:
ha effetti complessi sulla
fluidità delle membrane
Gli acidi grassi nei lipidi di membrana:
le code degli idrocarburi formano una barriera
idrofobica per la diffusione dei soluti polari.
Grassi insaturi: aumentano la fluidità di membr.
ostacolando l’impacchettamento delle altra
molecole
Lunghezza delle catene: 12-20 atomi di C, in media
16-18.
Lunghezza ottimale per la formazione di un doppio
strato di spessore 6-8 nm.
Legami semplici
Legami doppi
Tra le catene degli acidi grassi: legami deboli di
Van der Waals.
LIPIDI
I lipidi di membrana: la componente
“fluida” del modello di membrana a
mosaico fluido.
Sono molecole anfipatiche che
formano spontaneamente un doppio
strato.
Il colesterolo
Effetti sulla fluidità:
1- a t relativamente alta (37°C) stabilizza le membrane
riducendo il movimento dei fosfolipidi
2- a t relativamente bassa impedisce l’impacchettamento
stretto dei fosfolipidi
Fluidità di membrana
Flip-flop: un evento raro
E’ invece sostenuta la rotazione e
la diffusione laterale.
I lipidi di membrana sono un solvente
bidimensionale per le proteine di membrana e
regolano con la loro composizione la funzione delle
proteine di membrana.
LE PROTEINE DI MEMBRANA
Svolgono la maggior parte delle funzioni specifiche della
membrana plasmatica
Possono essere glicosilate sulla faccia esterna (glicocalice o
rivestimento cellulare)
Circa il 30% delle proteine codificate sono proteine di
membrana
LE PROTEINE DI MEMBRANA
Per studiarle: criofrattura della membrana lungo lo strato idrofobico
LE PROTEINE DI MEMBRANA
Periferiche
Integrali
monopasso
Ancorate ai lipidi
multipasso
LE PROTEINE DI MEMBRANA
Porzione esterna (glicosilata)
con ponti disolfuro
Alfa elica destrogira
Dominio citoplasmatico
LE PROTEINE DI MEMBRANA
LE PROTEINE DI MEMBRANA
Le membrane sono MOSAICI
strutturali e funzionali
Le proteine di membrana sono di diversi tipi e
quindi hanno funzioni diverse
Recettori
Enzimi
Trasportatori
Come le tessere di un mosaico danno figure diverse nel loro complesso
Il modello strutturale della membrana è uno, ma
le membrane sono tutte diverse per composizione e assortimento delle
singole “tessere”
MODELLO A MOSAICO FLUIDO
Fluidità delle membrane
sistema dinamico
movimento attraverso
le membrane
CARATTERISTICHE FUNZIONALI E STRUTTURALI
MEMBRANA DINAMICA
Glicocalice: funzioni
glicoproteine
glicolipidi
Il rivestimento è composto da
catene polisaccaridiche legate
covalentemente alle proteine
transmembrana o ai
fosfolipidi di membrana del
foglietto esterno.
Lo spessore è vario, può
arrivare fino a 50 nm in
alcune cellule di rivestimento,
come nel tratto digestivo
• evitare alla cellula contatti con
proteine indesiderate,
chimici e fisici
le
con aggressivi
• Riconoscimento cellula-cellula
Es. coagulazione del sangue, risposte
immunitarie
In conseguenza della sua composizione e della sua
architettura molecolare la membrana plasmatica è
SEMIPERMEABILE
costituisce il dispositivo di regolazione degli
scambi fra l’ambiente extra ed intra
cellulare: si ha cioè un flusso bidirezionale
ininterrotto di molecole con caratteristiche
diverse.
I componenti idrofobici della membrana limitano enormemente il passaggio di
molecole idrofiliche attraverso il doppio strato lipidico.
Solo la presenza e la particolare attività di proteine transmembrana specializzate
rendono possibile il passaggio di alcune molecole attraverso questa barriera
Funzioni delle membrane cellulari
1) Supporto strutturale
Preservare l’individualità della cellula
Delimitazione: cellula-ambiente; organulo citoplasma
Mantenimento della forma, plasticità: rapporti con il citoscheletro
2) Scambio di molecole
Mantenere una permeabilità altamente selettiva
Trasporto regolato in modo attivo
3) Scambio di informazione
Controllare il flusso di informazione
Rapporti cellula-cellula e cellula-matrice extracellulare
Riconoscimento e “trasduzione” di segnali chimici
Funzioni di membrane specializzate
Es. membrana nucleare, mitocondriale, del golgi e RER, delle vescicole, dei lisosomi ecc…
MEMBRANA CELLULARE: SELETTIVITA
PERMEABILITÀ SELETTIVA –Membrana
Trasporto di membrana
molecole piccole
molecole grosse
endocitosi
Trasporto passivo
Diffusione semplice
Trasporto attivo
(con pompe o cotrasportatori)
Diffusione facilitata
(da canali o permeasi)
esocitosi
MECCANISMI DI TRASPORTO –Membrana
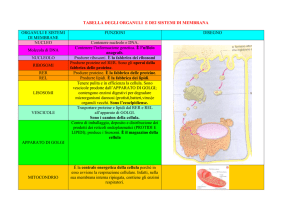
CITOPLASMA E ORGANULI CELLULARI
Cellula pancreas esocrino
al ME
ORGANULI CELLULARI
Nucleo
Reticolo Endoplasmatico
Apparato del Golgi
Mitocondri
Lisosomi
Sistema vacuolare di
membrana
RETICOLO ENDOPLASMATICO
LISCIO (REL)
RUGOSO (RER)
microsomi ottenuti per centrifugazione al ME
Reticolo endoplasmatico
FUNZIONI:
•Sintesi e rimaneggiamento delle proteine
(RER)
•Sintesi di lipidi e steroidi (REL)
•Degradazione dei lipidi (REL)
•Detossificazione di alcuni composti (REL)
•Rimaneggiamento di tutte le membrane
della cellula (RER)
Reticolo endoplasmatico liscio
Il REL, in forma di piccole vescicole o sacculi
indipendenti è particolarmente abbondante nelle cellule
epiteliali epatiche (detossificazione dei matariali tossici)
e in tutte le cellule che producono ormoni steroidei.
Reticolo endoplasmatico liscio
assume caratteristiche di specializzazione in alcune
cellule (fibre muscolari scheletriche, cellule
miocardiche dove è presente in forma di tubuli
- reticolo sarcoplasmatico-)
Funzioni del REL
Deposito di Calcio
Funzioni del REL
Deposito di Calcio
Reticolo endoplasmatico rugoso
La cavità sono in continuità con la cisterna perinucleare
(spazio esistente tra la membrana interna ed esterna
dell’involucro nucleare)
Reticolo endoplasmatico rugoso
La superficie esterna delle
membrane puo’ essere rivestita di
granuli di 15-20 nm:
i RIBOSOMI
RIBOSOMI
Ogni ribosoma è costituito da una subunità grande e
da una piccola entrambe prodotte nel nucleolo
Funzionano da base strutturale per la
sintesi proteica
RIBOSOMI : Meccanismo sintesi proteica
proteine liberate nel lume del RER
Modalità di sintesi e maturazione proteica
Proteine integrali con NH3 terminale citosolico
Proteine integrali con COOH terminale citosolico
Proteine integrali con estremità terminali citosoliche
Maturazione proteica nel lume del RER: es. Glicosilazione
RER
Zolle di Nissl
Apparato del Golgi
È costituito da una o più serie di cisterne appiattite e
leggermente incurvate (pile del Golgi) e vescicole
collegate tra loro
Apparato del Golgi
Le funzioni dell’apparato
di Golgi consistono:
1) Nel completamento della
sintesi di glicoproteine e
glicolipidi
2) Condensazione dei
prodotti di sintesi e
segregazione in organuli
delimitati da membrana.
Apparato del Golgi
Le proteine
presentano un
particolare segnale
che le dirige verso il
Golgi
Durante questi
trasferimenti all’interno
del Golgi le proteine
vengono modificate
(es. glicosilate,
acetilate, fosforilate)
Apparato del Golgi
Le proteine che escono dall’apparato del Golgi
vengono racchiuse in vescicole e possono avere
diversi destini
Destino delle vescicole
ENDOCITOSI
ESOCITOSI
GEMMAZIONE
Endocitosi
Macrofaco che fagocita un
globulo rosso (GR), img
ME
Esocitosi
Vacuolo gemmato
dall’apparato del Golgi
Spazio extracellulare
Le proteine prodotte nel RER gemmano come vescicole di
transizione, si portano al Golgi e si staccano come vescicole di
condensazione; quindi come vescicole di secrezione migrano nel
citoplasma apicale dove le membrane si fondono rivestendone il
contenuto all’esterno
Transcitosi
Esempio: transcitosi di anticorpi materni
verso epitelio intestinale del neonato
Fusione delle vescicole
La fusione delle vescicole è regolata
dalla presenza di specifiche proteine dette SNARE
Destino delle vescicole
lisosomi
Sono vescicole
rotondeggianti
contenenti idrolasi
acide e
costituiscono il
sistema “digestivo”
della cellula
Lisosomi
Intervengono nella degradazione di:
• macromolecole
• microrganismi
• frammenti cellulari fagocitati
• organuli cellulari invecchiati
I vari enzimi digeriscono il materiale fagocitato
degradandolo in prodotti terminali di basso peso molecolare
che, trasportati da particolari proteine di trasporto,
fuoriescono dai lisosomi e possono essere riversati nello
spazio extracellulare oppure riusati dalla cellula
Lisosomi
Le idrolasi in essi contenute non vengono mai in
contatto con il citoplasma
M = membrano
C = cuore elettron-denso
Lisosomi
Nelle cellule che sono destinate a morte
programmata (apoptosi) i lisosomi riversano il loro
contenuto all’interno della cellula, provocando
AUTOLISI
Mitocondri
I mitocondri sono organuli citolpasmatici presenti in tutte le cellule
eucariotiche, ad eccezione dei globuli rossi dei Mammiferi.
Il numero dei mitocondri/cellula varia da 1.000 a 2.000,
fino a 30.000 negli ovociti.
Mitocondri
La forma più comune è quella bastoncellare con lunghezze di 1 - 6 µm x 0,2-1
µm. Più rari sono mitocondri ovali o sferoidali. Le forme sferoidali possono
divenire filamentose e viceversa.
I mitocondri filamentosi possono gemmare rami collaterali destinati a
costituire un mitocondrio indipendente.
L’insieme di tutti i mitocondri della cellula, cioè il loro volume, e’ detto
condrioma.
E’ possibile determinarlo attraverso una centrifugazione frazionata.
Mitocondri
La membrana interna e quella esterna
sono di natura lipoproteica anche se le
due frazioni sono differentemente
ripartite.
La frazione proteica è costituita da una
varietà di circa 60 tipi di proteine
idrofobe, trasportatrici e della catena
respiratoria.
Mitocondri
**
*
Tra le due membrane
mitocondriali rimane compreso
un sottile spazio di circa 10 nm,
la camera mitocondriale
esterna *.
La camera mitocondriale
interna ** è variabile a seconda
dell’estensione delle creste.
Il contenuto della camera interna corrisponde alla matrice
mitocondriale.
Essa contiene dei ribosomi (mitoribosomi)
DNA Mitocondriale !!!
Nella camera interna sono contenute 5 10 molecole di mDNA
mitocondriale circolare e non associato a proteine . (vd cellula
epatica di ratto foto)
Mitocondri=Centrale Energetica