I GENI ASSOCIATI
Bateson , Saunders e Punnett: incrocio diibrido in Lathirus odoratus (1905)
• In F2 non viene rispettato il rapporto di segregazione 9 : 3 : 3 : 1
• Anche nella F2 dell’incrocio in trans sono i parentali ad essere
nettamente prevalenti sui ricombinanti
• Bateson e coll. non seppero spiegare i risultati ottenuti.
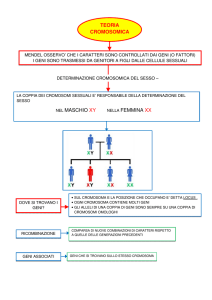
TEORIA CROMOSOMICA dell’ EREDITA’ (Sutton e Boveri 1904)
“La associazione di cromosomi materni e paterni in un numero prefissato di coppie
per ciascun organismo vivente e la loro successiva separazione alla meiosi
costituiscono la base fisica dell’ eredità mendeliana”
Il comportamento dei cromosomi alla meiosi può spiegare le leggi di Mendel
supponendo che i ‘fattori’ siano portati dai cromosomi.
Sutton e Boveri arrivarono a questa teoria grazie ai loro studi citologici al microscopio
ottico ma anche grazie agli studi sulla meiosi e la fecondazione di Hertwig, Fleming,
Strasburger e van Beneden alla fine dell’800, studi successivi al lavoro di Mendel del
1865.
La diffusione di queste scoperte ha permesso a Morgan
• di provare che i geni sono portati dai cromosomi
• di scoprire la trasmissione dei geni associati
Thomas Hunt Morgan
Premio Nobel per la medicina 1933
Drosophila melanogaster
Drosophila melanogaster
Femmina (a sinistra) e maschio (a destra) di
Drosophila melanogaster
La femmina ha nelle cellule somatiche quattro
paia di cromosomi, compreso un paio di
cromosomi X.
Nel maschio si osservano un paio di cromosomi
sessuali XY.
Morgan analizzò diversi caratteri e chiamò:
selvatico il fenotipo più diffuso nella popolazione
mutante il fenotipo alternativo (raro) dello stesso
carattere
Reincrocio per due geni indipendenti:
BC1
liscio, giallo x rugoso, verde
gameti
SsGg
ssgg
25% SG
100% sg
per reincrocio (BC) si
intende l’incrocio di un
ibrido con uno dei due
parentali, generalmente
quello recessivo.
25% S g
25% s G
25% s g
Risultato:
25% SsGg
liscio, giallo
PARENTALI
25% Ssgg
liscio, verde
RICOMBINANTI
25% ssGg
rugoso, giallo
RICOMBINANTI
25% ssgg
rugoso, verde
PARENTALI
50%
Si definiscono parentali i fenotipi che presentano le caratteristiche di uno dei due parentali e ricombinanti
quelli che presentano caratteristiche fenotipiche di entrambi i genitori
Morgan e coll. : reincrocio diibrido in Drophila melanogaster (1911)
P
F1
b+b+ vg+vg+ x
b b vg vg
b+ b vg+ vg
b+ corpo grigio
b corpo nero
reincrocio
vg+ ali normali
vg ali vestigiali
(in cis)
Ottennero: 83% parentali
17% ricombinanti
Con il reincrocio in trans si ottennero gli stessi risultati
Risultati molto diversi furono ottenuti utilizzando come individuo F1 un maschio
in CIS
P
grigio
normali
x
nero
vestigiali
F1
maschio grigio
normali
BC1
50% grigio
normali
100% PARENTALI
x femmina nero
vestigiali
50% nero
vestigiali
0% RICOMBINANTI
Risultati degli esperimenti di Morgan e collaboratori:
• i due geni b e vg non sono a trasmissione indipendente, sono associati o
concatenati, tendono a essere trasmessi insieme
• l’associazione non è completa perché si trovano comunque dei ricombinanti (17%)
• L’associazione può essere completa nel maschio di Drosophila
Con il
reincrocio in
trans si
ottennero gli
stessi risultati
Interpretazione dei risultati degli esperimenti di Morgan e coll.
1. I geni analizzati erano associati (cioè tendevano a rimanere insieme durante la segregazione
degli alleli alla meiosi) perché erano situati sullo stesso cromosoma
Esperimenti di Bridges su ceppi di Drosophila con non disgiunzione del cromosoma X
e di Morgan sui caratteri legati al cromosoma X
fornirono la dimostrazione che i geni sono localizzati sui cromosomi e quindi della validità
della teoria cromosomica della ereditarietà
Eredità legata al cromosoma X:
in Drosophila (Morgan, 1910)
occhi rossi, selvatico
occhi bianchi, mutante
Interpretazione dei risultati degli esperimenti di Morgan e coll.
2.
Spiegazione della ricombinazione tra geni associati: ipotesi del crossing over in base ai
dati citologici di Jansenns 1909 al microscopio elettronico
Durante la meiosi se avviene il crossing-over si ha
uno scambio fisico reciproco di parti tra i due
cromosomi omologhi (tra cromatidi non fratelli)
con la formazione di combinazioni alleliche non parentali,
ricombinanti, dei geni associati
A
a
Aa
B
b
b B
Interpretazione dei risultati degli esperimenti di Morgan e coll.
3.
Spiegazione della assenza di ricombinazione nel maschio di Drosophila
Nel maschio di Drosophila non avviene il crossing over, ma è una eccezione.
4.
L’entità della concatenazione fra geni si può determinare :
calcolando la frequenza di ricombinazione, cioè il rapporto fra individui (gameti)
ricombinanti e individui totali in un reincrocio.
Come si indicano i geni concatenati:
A
a
B
b
A
a
B
b
oppure
a+ b+
a b
eterozigote
in CIS
oppure +
a
+
a
b
+
+
b
in Drosophila
eterozigote
in TRANS
Interpretazione della ricombinazione tra geni indipendenti e tra geni associati
MEIOTICA
MEIOTICA
MEIOTICA
MAPPE GENETICHE
Morgan: “Gli elementi fisici responsabili dei caratteri ereditari di un organismo (i geni) si
trovano ordinati linearmente nei cromosomi e le percentuali di ricombinazione osservate
negli incroci fra caratteri concatenati sono proporzionali alla distanza, sul cromosoma, fra
i geni corrispondenti”
Su questo presupposto si basa il metodo messo a punto da
Sturtevant (1913-15) per la costruzione di mappe genetiche
Una mappa genetica attribuisce i geni ai
gruppi di associazione (i cromosomi) e,
all’interno di ogni gruppo di associazione,
ordina i singoli geni secondo la loro posizione
l’uno rispetto all’altro.
Mappatura di 4 geni con incroci a due punti (incroci diibridi e reincrocio):
Esempio in Drosophila: i geni dichaete (d), pink (p), hairless (h) e rough (ro) si trovano sullo stesso cromosoma.
P
DP
DP
F1
DP
dp
x
dp
dp
DP
dp
x
dp
dp
parentali (93%)
dp
dp
Dp
dp
Test Cross1
(BC1)
dP
dp
ricombinanti (7%)
La distanza fra d e p è di 7 cM
• L’unità di misura è il centimorgan cM: la distanza di 1cM è la distanza
per la quale si ottiene l’1% di ricombinanti
Geni Frequenza
di ricombinazione
d-h
h-p
d-p
h - ro
d - ro
p - ro
23,4%
21,1%
7,0%
21,9%
36,0%
34,0%
d -7- p ----21,1---- h
d ------23,4------ h ----21,9---- ro
d ------------36------------ ro
p ---------34--------- ro
La distanza tra d e h è minore della somma della distanza tra d e p e tra p ed h !?
Perché?
• I Doppi Crossing-over portano ad una sottostima della
distanza tra i geni in esame
Se la frequenza di ricombinazione fra due geni è maggiore del
15% è più conveniente fare un incrocio a tre punti
(perché maggiore è la distanza, maggiore è la probabilità che
avvengano i DCO) (15% dato sperimentale)
INCROCIO A TRE PUNTI
st occhio scarlatto
e corpo ebano
ss setole corte
P
F1
st+ occhio rosso
e+ corpo grigio
ss+ setole normali
st+ e+ ss+
st+ e+ ss+
x
st+e+ss+
st e ss
st e ss
st e ss
x
st e ss
st e ss
Back Cross
INCROCIO A TRE PUNTI
• 8 tipi di gameti con diverse frequenze
• 6 tipi di gameti ricombinanti e 2 parentali
• I doppi ricombinanti sono i meno frequenti e
indicano quale è il gene centrale
• Calcolo delle distanze tra i geni:
Distanza st – e
Distanza e--ss
st
50+52+5+3 = 110 = 0,1456
755
755
43+41+5+3 = 92 = 0,1218
755
755
14,56 cM
e 12,18 cM
ss
14,56 %
12,18 %
INTERFERENZA DA CHIASMA
Il verificarsi di un crossing over in una regione puo’ influenzare il verificarsi di un altro
crossing over in una regione vicina
Interferenza positiva : la frequenza di doppi crossing over è inferiore rispetto all’atteso
Interferenza negativa : la frequenza di doppi crossing over è maggiore rispetto all’atteso
COEFFICIENTE DI COINCIDENZA (C)
Frequenza di doppi ricombinanti osservata
C=
Frequenza di doppi ricombinanti attesa
INTERFERENZA = 1 - C
Se C < 1 interferenza positiva
Se C > 1 interferenza negativa
Se C = 1 non c’è interferenza
C= coefficiente di coincidenza
C= Freq dco OSS
Freq dco ATT
Nell’esempio
I = Interferenza
I = 1-C
1-0,55= 0,45
8/ 755
0,146x 0,122
C = 0,55
C<1
I = 0,45
• L’interferenza si ha se i geni si trovano sullo stesso braccio cromosomico e se le distanze sono
molto piccole
• Il grado di interferenza varia in funzione dell’organismo e del cromosoma
• Nelle piante si sono osservate spesso interferenze negative: il verificarsi di un c.o. in una regione
aumenta la probabilità che si verifichi un altro c.o. nella regione adiacente
Mappa genetica di Drosophila
2n = 8
Mappa genetica di pomodoro
2n = 24
Mappe genetiche e mappe fisiche
•
Le distanze genetiche, calcolate
in base alla frequenza di
ricombinazione, non sempre
corrispondono alle distanze fisiche
calcolate in pb
• Il crossing over non avviene lungo
il cromosoma in modo casuale:
esistono gli “hot spots” regioni ad
alta frequenza di ricombinazione e
regioni in cui i crossing over sono
rari (es. i telomeri)
Drosophila
S. cerevisiae
• Oggi le mappe genetiche si costruiscono con i marcatori molecolari
• Sono disponibili mappe genetico-molecolari in cui i singoli cromosomi
sono saturati con un alto numero di loci
• Es mais (2001) 10 gruppi di associazione con 1772 loci mappati che
coprono una grandezza di 1800 cM (B&F 2005)
Evoluzione storica delle mappe genetiche in mais
Mappa di associazione di tre cromosomi di mais
24
Ricapitolando…..
D’Amato: Una mappa genetica è la rappresentazione di ogni “gruppo di associazione”
(cioè di ogni cromosoma del corredo aploide) di una specie in forma di un segmento
lungo il quale sono indicati in sequenza i geni con la loro distanza di mappa rispetto a
un gene terminale (punto 0).
Per costruire una mappa genetica occorre :
• scegliere geni che siano associati, cioè appartengano allo stesso gruppo di
associazione
• definire tanti gruppi di associazione quanti sono i cromosomi del corredo aploide
• calcolare le distanze tra i geni in base alle frequenze di ricombinazione in appositi
Reincroci a due punti (con due marcatori) o a tre punti (con tre marcatori)
Dalla analisi della progenie di un icrocio a tre punti si può
determinare:
il gene centrale
la distanza tra il gene centrale e gli altri due geni
l’ interferenza
• Localizzare un numero elevato di geni marcatori per ogni cromosoma che coprano
l’intera grandezza del cromosoma
Esercitazione n. 4
1) In mais, l’epidermide del seme colorata (R) è dominante sull’ epidermide incolore (r) e la pianta verde (T ) è dominante
sulla pianta gialla (t) . Reincroci di individui F1 eterozigoti dettero la seguente progenie:
Epid colorata pianta verde
46
Epid colorata pianta gialla
340
Epid incolore pianta verde 380
Epid incolore pianta gialla
34
totale
800
Nella F1, i due geni associati erano in (cis) o(trans)? Calcolate la percentuale di ricombinazione.
Si considerino tre geni, d (ali dichaete ) p (occhio pink) e (corpo ebony) di Drosophila. Dall’incrocio di un triplo
eterozigote con il triplo recessivo si ottiene la seguente progenie:
ali normali, occhio pink, corpo ebony (+pe) = 824; ali dichaete, occhio rosso, corpo grigio (d++ )= 878;
ali normali, occhio pink, corpo grigio (+p+) = 214; ali dichaete, occhio rosso, corpo ebony (d+e) = 187;
ali normali, occhio rosso, corpo ebony (++e) = 11; ali dichaete, occhio pink, corpo grigio (dp+) = 18;
ali dichaete, occhio pink,corpo ebony (dpe) = 28;
ali normali, occhio rosso, corpo grigio (+++) = 31.
Determinare: a) l’ordine dei geni sul cromosoma; b) le distanze di mappa; c) il coefficiente di coincidenza.
2)
3) Nel mais le tre coppie alleliche + c (aleurone colorato a. incolore) + sh (endosperma turgido e. raggrinzito)
+ wx (endosperma amiloso e. ceroso) sono localizzate sullo stesso cromosoma.
Dall’incrocio tra il triplo eterozigote e il triplo recessivo è stata osservata la seguente progenie:
c + sh 113, +++ 4, c wx + 2708, c + + 626, c wx sh 2, + wx + 116, + + sh 2538, + wx sh 601.
Indicare l’ordine dei geni e le distanze di mappa.
5) Nel cromosoma 2 di mais i 3 loci a, r e d presentano questa associazione:
a
15
r
25
d
Quali classi fenotipiche e con quale frequenza si troveranno nella progenie dell’incrocio +++/ard x ard/ard su
un totale di 1000 piante, in presenza di una interferenza positiva del 33% ?
Determinazione genetica del sesso negli animali
Primi esperimenti sul controllo del sesso negli animali
McClung; Wilson; Henking primi anni 900 su diverse specie di insetti
Insetti: presenza di cromosomi responsabili della differenziazione sessuale.
Eterocromosomi (detti X e Y)
Autosomi
Digametia maschile
Costituzione cromosomica e sesso in Drosophila
Il sesso è determinato dal rapporto fra numero di cromosomi X e numero di corredi
cromosomici aploidi autosomici.
N° cromosomi X
N° corredi autosomici
aploidi
Costituzione cromosomica
2A
2A
2A
3A
4A
3A
3A
4A
2A
2A
2A
4A
3A
XXX
XX
XXY
XXX
XXXX
XX
XXY
XXX
X
XY
XYY
XX
X
= X/A = 1
femmina
Rapporto X / A
N° cromosomi X
N° corredi autosomici
aploidi
Sesso
1,5
1,0
Superfemmina
Femmina
0,67
Intersesso
0,75
0,50
Intersesso
Maschio
0,33
Supermaschio
= X/A = 0,5
maschio
N° cromosomi X
N° corredi autosomici
aploidi
= X/A = 1
femmina
N° cromosomi X
N° corredi autosomici
aploidi
= X/A = 0,5
maschio
Negli uccelli e in alcuni insetti la digametia è femminile
e gli eterocromosomi si indicano con Z e W
UOMO
44 cromosomi autosomici
2 cromosomi sessuali X e Y
Digametia maschile
Il cromosoma X contiene 1098 geni di cui solo 54 hanno l’omologo sull’ Y (Nature 2005)
29 localizzati nelle regioni PAR1e PAR2, gli altri 25 fuori da queste regioni
PAR = Regione Pseudo Autosomica
Eredità legata al sesso: in Drosophila occhi rossi: selvatico; occhi bianchi: mutante
(Morgan, 1910)
Conclusioni di Morgan :
•
Occhio bianco è recessivo rispetto
all’ occhio rosso
•
Il gene per il colore degli occhi è
portato dal cromosoma X
•
Il fenotipo bianco si manifesta anche con un solo
allele nei maschi che hanno un solo cromosoma
X (emizigosi)
Il carattere “colore degli occhi” in Drosophila è legato al sesso
Y
X
XX
Schema di eredità
“Criss-cross”
di un carattere
legato al sesso
Albero genealogico della Regina Vittoria di Inghilterra
Eredità legata al sesso: eredità della emofilia nell’uomo
Albero genealogico della Regina Vittoria e dei suoi discendenti, che illustra la trasmissione della emofilia.
La Regina Vittoria era eterozigote per l’allele recessivo dell’emofilia legato al cromosoma X e non
essendovi casi di emofilia nei suoi antenati, è plausibile che la mutazione sia insorta nelle cellule germinali
della madre
P
Vittoria XEXe
XEXE
1/4
XEXe
1/4
XE XE
P
XEY Alberto
x
XEY
1/4
XeY
x
Alice(portatrice) XEXe
1/2
P
Alice(portatrice) XEXe
XEXE
1/4
XEXe
1/4
X eY
1/4
XEY
1/4
Leopoldo
(emofiliaco)
Leopoldo
(emofiliaco)
XE Y
1/2
x
XEY
XeY Rupert (nipote di Leopoldo
1/4
emofiliaco)
DETERMINAZIONE GENETICA DEL SESSO NELLE PIANTE
Specie monosessuali o dioiche:
fiori maschili e femminili su individi diversi
es. kiwi, pioppo, salice, cannabis sativa e indica
luppolo, papaia, palma da datteri, asparago
Specie bisessuali:
monoiche
fiori maschili e femminili sulla stessa pianta
es. mais, sorgo
ermafrodite
Specie subdioiche
Specie intersessi
organi maschili e femminili sullo stesso fiore
piante a sessi separati, con alcun individui bisessuali
piante a sessi separati con alcuni individui ermafroditi e sterili
Cromosomi sessuali eteromorfici nelle piante (specie dioiche, piante a sessi separati)
Tipo Melandrium = digametia maschile XY (come nell’uomo)
22A +X Y maschio
22 A XX femmina
Tipo Rumex acetosa = digametia maschile: maschio 2n=15 (12A+X+Y1+Y2)
femmina 2n=14 (12A+X+X)
X/A come Drosophila però Y è
indispensabile per produrre polline fertile
Tipo Fragaria = digametia femminile nelle varietà poliploidi
le varietà diploidi sono ermafrodite
Melandrium album (Sinapis alba) 22A+XX 22A+XY
Disegno schematico dei cromosomi del sesso in Melandrium.
I-II e III: segmenti differenziali del cromosoma Y;
V: segmento differenziale del cromosoma X;
IV: segmenti omologhi, a livello dei quali avviene l’appaiamento di
X con Y.
Il segmento I reca il gene soppressore del sesso femminile
il segmento II contiene il gene che controlla l’inizio dello sviluppo
delle antere
il segmento III controlla gli ultimi stadi dello sviluppo delle antere;
il segmento V reca i determinanti del sesso femminile.
Oggi sappiamo che sull’X e sull’Y sono presenti due
geni strettamente associati:
SuF suF che sopprime lo sviluppo organi femminili
M m che ha forte effetto mascolinizzante
Sugli autosomi è presente un gene che potenzia SuF
Modello simile anche in pioppo e salice
FISH sui cromosomi di Melandrium album
Melandrium album: schema di eredità “criss-cross o “zig-zag” (gene angustifolia)
Femmina
Maschio
P
XAXA
x
XaY
F1
XAXa
e
XAY
F2
XAXA
1/4
XAXa
1/4
XAY
1/4
angustifolia
XaY angustifolia
1/4
Esercitazione n. 5
1) Caratteri legati al sesso in Drosophila sono: occhio bianco (w); corpo giallo (y); occhio rosso vermiglio
(vermilion, v); occhio a sbarra (Bar, dominante, B). Bar determina una riduzione della superficie dell’occhio. Il
carattere si manifesta in modo estremo (narrow bar) nel maschio emizigote e nelle femmine omozigoti, in modo
intermedio (wide bar) nelle femmine eterozigoti. Dare la spiegazione genetica del fatto che se una femmina con
occhi rossi è incrociata con un maschio a occhi bianchi dà una progenie metà con occhi rossi e metà con occhi
bianchi. Quali sono le F1 e F2 dell’incrocio fra femmina a corpo giallo e maschio ad ali vestigiali (carattere
autosomico)? Qual è il risultato dell’incrocio fra femmina wide bar e maschio bar?
2) Nei polli di razza Plymouth Rock barrata, il carattere piumaggio bianco legato al sesso è recessivo rispetto al
piumaggio nero barrato (rispettivamente b e B). Se maschi bianchi sono incrociati con femmine nere barrate,
quali saranno i fenotipi F2? Se una delle femmine barrate F2 è incrociata con un maschio barrato F2, che
proporzione di figli bianchi nascerà?
3) Nell’uomo i più noti caratteri legati al sesso sono: il daltonismo (recessivo), per cui gli individui affetti non
riescono a distinguere certi colori (di solito il rosso dal verde); l’emofilia (recessivo), per cui gli individui sono
soggetti a emorragie.
Se un uomo (figlio di un emofiliaco) che presenta sangue con tempo di coagulazione normale sposa una donna
normale, qual è la probabilità che abbiano figli maschi emofiliaci?
4)Dare genotipi e fenotipi dei seguenti incroci: 1) madre a occhi blu x padre a occhi bruni e daltonico, figlio di
padre a occhi blu e madre a occhi bruni, dei quali non si sa se uno dei due fosse daltonico (si ricorda che occhi
bruni è dominante su occhi blu e non è un carattere legato al sesso); 2) madre, figlia di emofiliaco e con occhi
blu e padre a occhi bruni (eterozigote) normale, ma anch’egli figlio di emofiliaco.
FUNZIONE DEL GENE
Beadle e Tatum 1941: Teoria un gene-un enzima
esperimenti con i mutanti nutrizionali di Neurospora
Via biosintetica dell’ arginina
FUNZIONE DEL GENE
Esistono proteine non enzimatiche allora la teoria un gene - un enzima viene riformulata
Un gene - una proteina
caratteristiche del fenotipo
Però esistono proteine formate da più subunità proteiche come l’emoglobina, allora
UN GENE--UNA CATENA POLIPEPTIDICA
Oggi sappiamo che il prodotto di un gene può essere anche una molecola di RNA
Esistono geni che codificano diversi tipi di RNA
FINE