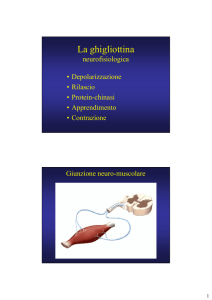
MICROFILAMENTI
● Mantenimento della forma della cellula
● Movimento ameboide
● Locomozione cellulare
● Contrazione cellulare
● Divisione cellulare
● Motilità intracellulare
MICROFILAMENTI
Struttura
Due catene di actina filamentosa
(F ACTINA)
Diametro
8 nm
Subunità
actina G (globulare)
Lo studio della funzione dell’actina si vale di diverse sostanze che ne
inibiscono la funzione:
CITOCALASINA induce depolarizzazione dell’actina
FALLOIDINA blocca la depolarizzazione
LATRUNCULINA blocca i monomeri liberi
b. Esposizione a
CITOCALASINA (5 min.)
Poiché ogni monomero di actina ha una polarità e tutte le
subunità sono orientate nel filamento allo stesso modo, il
filamento di actina F risulta polarizzato.
Ogni monomero è ruotato di 166° nel filamento, che quindi appare
come una doppia elica
L’actina è una proteina che lega e idrolizza ATP.
L’estremità + ha una affinità per l’actina/ATP dieci
volte maggiore dell’estremità -.
Quindi…l’estremità + cresce dieci volte più velocemente
della -
L’ATP è idrolizzato in un momento successivo
all’incorporazione del monomero nel polimero.
In teoria non serve l’ATP per far polimerizzare l’actina, ma
l’actina-ATP polimerizza più facilmente di actina-ADP, mentre
l’actina-ADP depolimerizza più facilmente.
(A mano a mano che procede la polimerizzazione l’actina
idrolizza l’ATP).
Come per i microtubuli anche per i microfilamenti è
molto importante l’assemblaggio ed il disassemblaggio
TREADMILLING
It occurs when one end of a
filament grows in length while the
other end shrinks resulting in a
section of filament seemingly
"moving" across a stratum or the
cytosol. This is due to the constant
removal of the protein subunits
from these filaments at one end of
the filament while protein subunits
are constantly added at the other
end
Esiste una concentrazione critica dei monomeri alla quale la velocità
del loro assemblaggio è uguale alla velocità di disassemblaggio:
filamenti in apparente equilibrio
Quindi….
1) La concentrazione critica di monomeri per l’aggiunta alla
estremità – è più alta di quella richiesta per la crescita
dell’estremità +
2) Dato che l’ATP e idrolizzato poco dopo l’incorporazione, a
concentrazioni critiche per l’estremità - è più facile trovare
actina-ATP all’estremità + e actina-ADP all’estremità –
3) Visto che l’actina-ADP depolimerizza più facilmente dell’actinaATP, al di sotto di una certa concentrazione di monomeri
l’estremità + cresce e la – depolimerizza.
Come per i microtubuli, le cellule mantengono un
equilibrio dinamico tra le forme monomerica e polimerica
dell’actina.
Cambiamenti delle condizioni locali della cellula
(influenzati da parecchie proteine accessorie diffusibili)
possono spostare l’equilibrio verso l’assemblaggio o il
disassemblaggio.
Questa riorganizzazione è necessaria per processi
dinamici come la locomozione cellulare, i cambiamenti di
forma e la citochinesi.
La miosina rappresenta il motore molecolare
dei filamenti di actina
(miosine: superfamiglia che comprende almeno 18 membri)
Miosini convenzionali coinvolte nelle
contrazioni muscolari, nella
citodieresi, nel mantenimento della
tensione alle adesioni focali
La Miosina II è formata da sei catene polipeptidiche : 2 catene
pesanti e 2 coppie di catene leggere.
Si distinguono: due teste globulari (siti catalitici), due colli (alfa
elica della catena pesante più due catene leggere), una coda
(avvolgimento di due alfa eliche delle catene pesanti)
MLCK: Myosin light chain kinase (Ser Threo kinase) fosforila la catena
Leggera della miosina, attivandola
La testa miosinica ha attività ATPasica. Due delle quattro catene leggere,
dette “regolatrici”, hanno siti di fosforilazione con cui controllano l’attività
delle teste e il dispiegamento delle code, lequali possono così interagire tra di
loro in modo parallelo o antiparallelo per formare aggregati di filamenti
bipolari.
Il ciclo ATPasico regola la contrazione della testa
miosinica sul filamento di actina
Il legame con l’ATP, l’idrolisi in ADP+Pi e il rilascio
dell’ADP provocano diverse modificazioni
conformazionali responsabili dello scatto della testa
della miosina legata al filamento di actina che viene
così spostato
MODELLO SCHEMATICO DELLA CONTRAZIONE ACTINA/MIOSINA
Actina e miosina II si
organizzano nell’unità
contrattile del muscolo
scheletrico, il
sarcomero. Due grandi
proteine, titina e
nebulina, contribuiscono
alla struttura e stabilità
del sarcomero.
La titina è una specie di
molla che tiene “aperto”
il sarcomero in
condizione di riposo.
La nebulina regola la
lunghezza e
l’assemblaggio dei
filamenti di actina.
Altre proteine
incappucciano
l’estremità+ del
filamento di actina.
Nel disco Z è contenuta
anche α-actinina.
Lo stimolo elettrico
determina un accumulo di
calcio intracellulare.
Il Ca++ agisce su due
proteine accessorie legate
all’actina, la troponina e la
tropomiosina.
A bassa concentrazione di
Ca++ la tropomiosina
interferisce con il legame
della miosina all’actina,
mentre alte concentrazioni
di Ca++ provocano uno
slittamento del complesso
troponina-tropomiosina,
che libera il sito di attacco
per la miosina sull’actina.
Miosine non convenzionali:
collaborano con i microtubuli nel
trasporto di diversi tipi di
vescicole
Rab27a regulates the localization and exocytosis of lysosome-related organelles in different
cell types through distinct effector proteins.
* identified disease mutations
Olkkonen V M , and Ikonen E J Cell Sci 2006;119:5031-5045
©2006 by The Company of Biologists Ltd
Assenza di gene normale per la miosina Va o per Rab27:
Sindrome di GRISCELLI
(autosomica recessiva), caratterizzata da ipopigmentazione,
Immunodeficienza (deficit dell’attività citotossica dei linfociti T,
Cellule natural killer)
Proteine associate ai microfilamenti sono
responsabili della conversione dei microfilamenti da una forma
all’altra
- Alcune controllano la polimerizzazione legandosi ai monomeri
e impedendo ai microfilamenti di aggregarsi
- Alcune li stabilizzano
- Alcune li frammentano
- Alcune raggruppano i microfilamenti in fasci paralleli
- Alcune li organizzano in reti tridimensionali
- Alcune si associano ai microfilamenti per formare fasci
contrattili
1. iniziazione: formina, complesso Arp2-3
8. Proteine che legano l’actina alla membrana ERM (ezrina, radixina moesina)
e membri delle spettrine (tra cui la DISTROFINA, proteina (se alterata)
responsabile della distrofia muscolare
La polimerizzazione dei microfilamenti di actina comincia
in un centro di nucleazione in cui la proteina dimerica
Formina assembla i primi monomeri di actina G. Anche la
più rapida polimerizzazione successiva è guidata dalla
formina che mantiene i monomeri di actina nella
conformazione corretta per una più efficiente
polimerizzazione
Il complesso proteico Arp2/3 dirige la
ramificazione (BRANCHING) dei microfilamenti
actinici, legandosi all’estremità+
WAS
Wiskott–Aldrich syndrome (WAS) is a rare X-linked recessive
disease characterized by eczema, thrombocytopenia (low platelet
count), immune deficiency, and bloody diarrhea (secondary to the
thrombocytopenia).
The main function of WASp is to activate actin polymerization by
binding to the Arp2/3 complex. In T-cells, WASp is important because it
is known to be activated via T-cell receptor (TCR) signaling pathways
to induce cortical actin cytoskeleton rearrangements that are
responsible for forming the immunological synapse leading to
IMMUNODEFICIENCY
Wiskott–Aldrich syndrome (WAS)
La Cofilina agisce:
a) come fattore
depolimerizzante
legandosi all’estremità-e
aumentando la velocità di
dissociazione dei
monomeri (ai quali resta
legata impedendone il
riassemblaggio)
b) spezzando i filamenti di
actina
La profilina stimola lo scambio tra ADP e
ATP dando luogo alla formazione di actina
G/ATP. Il complesso profilinaactinaG/ATP si lega più efficacemente del
singolo monomero all’estremità +
accelerando la polimerizzazione, ma in
assenza di un filamento da accrescere il
complesso blocca l’inizio della
polimerizzazone.
Fascio di filamenti paralleli
Fimbrina
Fascio contrattile
Α-actinina
Filamenti a rete
La filamina è una proteina dimerica a forma di V
che lega i filamenti di actina in reti ortogonali
LA FILAMINA FA ANCHE DA PONTE TRA
INTEGRINE E CITOSCHELETRO DI ACTINA
(vedi dopo)
Fasci di filamenti di actina formano la struttura centrale dei
microvilli
Associazione del citoscheletro di actina alla
membrana plasmatica: Adesioni focali integrine
matrice extracellulare
Talina fa da ponte
Tra le integrine e il
Citoscheletro di actina
Nelle adesioni focali.
Associazione del citoscheletro di actina alla
membrana plasmatica: Giunzioni aderenti-Ecaderina (adesione cellula cellula)
CATENINE
CADERINE (giunzioni dellula/cellula)
La dinamica dei riarrangiamenti dei filamenti di Actina è controllata
dalle piccole GTPasi della famiglia Rho: Rho, Rac, Cdc42
Disposizione dei microfilamenti all’interno della
cellula
Citodieresi (dipendente da Rho)
I microfilamenti (filamenti di actina) sono particolarmente numerosi al
di sotto della membrana plasmatica (ACTINA CORTICALE)
Organizzazione di complessi diversi di actina in una cellula
MATRICE EXTRACELLULARE (ECM)
E’ una rete organizzata di materiale extracellulare presente nelle vicinanze
della membrana plasmatica
MEMBRANA BASALE è uno strato sottile che si interpone tra l’epitelio e la
parte stromale di un organo.
MATRICE INTERSTIZIALE costituisce uno spazio che fa da connessione
e protezione delle strutture parenchimali, attraversato da vasi, nervi, che può
Essere attraversato da diversi tipi cellulari.
E’ prodotta principalmente da fibroblasti. E’ ricca in Collagene, Elastina,
Fibronectina, laminine
COLLAGENE
PROTEOGLICANI
FIBRONECTINA
RGD=arg gly asp
è riconosciuto dalle integrine beta-1
Anche la proteina TAT del virus HIV
Contiene un dominio RGD che può
Portare ad una attivazione di integrin
MEMBRANA BASALE
MATRIGEL
E’ una matrice cellulare prodotta da un
sarcoma utilizzata in laboratorio come
substrato di colture cellulari per mimare la
matrice cellulare. Contiene laminina,
collageni, proteine adesive e fattori di
crescita
INTEGRINE
• Svolgono un ruolo chiave nell’interazione tra
l’ambiente intracellulare e quello extracellulare.
• Sono conservati in tutti gli organismi eucarioti a
partire dal lievito.
• 2 catene polipeptidiche (alfa e beta).
• Proteine transmembrana con un piccolo dominio
intracellulare. Non hanno attività chinasica
• Legano cationi bivalenti (Ca2+, Mg2+, Mn2+,)
importante per la loro attivazione.
FUNZIONI DELLE INTEGRINE
• 1. Adesione cellula/cellula o cellula/ECM
• 2. Trasduzione del segnale al nucleo per
indurre espressione genica. Cross talk tra
Integrine e fattori di crescita
Ruolo delle integrine nella aggregazione piastrinica: interazione di alfa2b/beta3
Con il motivo RGD del fibrinogeno
PATOLOGIE CAUSATE DA MUTAZIONI GENETICHE DELL’ESPRESSIONE
DI INTEGRINE
Deficit di integrina beta-1 sono incompatibili con la vita (nel topo l’embrione -/muore al giorno 4.5 di gestazione per difetto di impianto
Deficit di integrina beta2:
LEUKOCYTE ADHESION DEFICIENCY (LAD)
È una rara patologia autosomica recessiva. E’ caratterizzata da
immunodeficienza che porta ad infezioni ricorrenti. I granulociti sono sono in
grado di migrare nei siti di infezione
Alterazione genetiche integrine di alfa6-beta4 ed altre molecole connesse nella
adesione dei cheratinociti alla membrana basale: EPIDERMOLISI BOLLOSA
INTERAZIONE INTEGRINE/MATRICE EXTRACELLULARE:
LE ADESIONI FOCALI
Control of actin organized cell structures by small Rho GTPases
Induced by Integrin Activation
Stress fibers are organized by Rho
Lamellipodia are organized by Rac
Filopodia are organized by Cdc42
EXTRACELLULAR MATRIX RIGIDITY
INTEGRINS
FOCAL ADHESIONS
RhoA
Rho Kinase
Myosin II phosphorylation
Increased acto-myosin contractility /Stress fibers
Vie di trasduzione del segnale indotte dalle GTPasi Rac e Cdc42 che portano
Alla polimerizzazione dell’actina
Migrazione mesenchimale (lamellipodio) tipica del miofibroblasto
Migrazione ameboide tipica del leucocita o varie cellule tumorali
Questa è una visione classica basata su esperimenti condotti in 2D. Cellule
possono adottare diversi tipi di migrazione a seconda del substrato più o meno
rigido, della possibilità di degradare il substrato tramite proteasi
Migrazione mesenchimale/migrazione ameboide
©2012 by The Company of Biologists Ltd
Migrazione in 2D
Falloidina
(actina)
Direzione della
migrazione
Beta1 integrine
(focal adhesions)
Lamellipodio (Rac)
Coda
Migrazione in 3D
fibronectina
Falloidina
(actina)
paxillina
(focal adhesion)
Proteasi degradano la matrice
Attività di remodeling e forza
per farsi strada attraverso
i fasci di matrice
lobopodio
Forma della cellula = funzione cellulare è totalmente diversa tra
2D e 3D
3D fibroblasti
cresciuti su matrice
extracellulare
2D fibroblasti
cresciuti su plastica
Generazione di scaffold sintetici in materiale biocompatibile per
favorire la crescita cellulare 3D. Scaffod di tessuto osseo, generazione di
organi tridimensionali