")
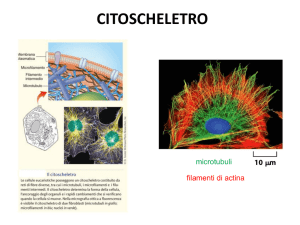
Citoscheletro
Per funzionare correttamente le cellule devono
organizzarsi nello spazio ed interagire
meccanicamente con il loro ambiente; esse
devono avere una forma corretta ed essere
strutturate internamente in modo appropriato;
molte di esse devono essere in grado di cambiare
forma e di muoversi; tutte devono essere in grado
di riorganizzare i loro componenti interni mentre
crescono e si dividono. Tutte queste funzioni sia
spaziali che meccaniche dipendono da un
notevole sistema di filamenti chiamato
citoscheletro
Alcune funzioni del citoscheletro
-Esercita una trazione sui cromosomi allontanandoli durante
la mitosi e quindi taglia la cellula in divisione in due cellule
-Spinge e guida il traffico intracellulare di organelli e
vescicole trasportando materiali da un punto all’altro della
cellula
-Sostiene la fragile membrana plasmatica e fornisce
collegamenti meccanici che permettono alla cellula di
sopportare stress e tensioni senza lacerarsi
-Permette ad alcune cellule di “nuotare” (spermatozoi) e ad
altre (come fibroblasti e globuli bianchi del sangue) di
strisciare su superfici
-Fornisce il macchinario alla cellula muscolare per la
contrazione e al neurone per estendere assoni e dendriti
-Guida la crescita della parete cellulare vegetale
-Controlla la diversità delle forme delle cellule eucariotiche
Il Citoscheletro
1)Il citoscheletro della cellula eucariotica è una
intelaiatura interna di filamenti che comprende
microtubuli, filamenti di actina
(microfilamenti), filamenti intermedi. Questi
filamenti creano una struttura interna più
rigida che stabilisce la forma della cellula; ad
esempio le cellule epiteliali possono avere una
forma cubica, mentre i neuroni possono avere
molti assoni sottili di notevole lunghezza
Quindi possiamo dire che la forma di una
cellula eucariotica è determinata dal suo
citoscheletro
*
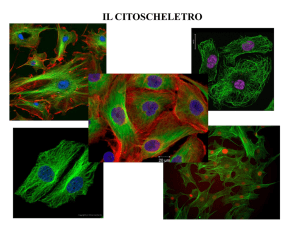
Cellula in coltura fissata e colorata per evidenziare due
dei suoi principali componenti citoscheletrici, i microtubuli
(verdi) ed i filamenti di actina (rossi); il DNA è colorato in
blu
Figure 16-1 Molecular Biology of the Cell (© Garland Science 2008)
Consideriamo i microtubuli in cellule
estremamente diverse come i fibroblasti
ed i neuroni (vedi immagini successive)
*
I fibroblasti
sono cellule
mobili che
si spostano
all’interno
dell’organi
smo. Da un
singolo
punto vicino
al nucleo i
microtubuli
si irradiano
con una
configurazio
ne a stella
I lunghi processi
rappresentati da
assoni e dendriti
che emergono
dal corpo
cellulare di un
neurone sono
estesi per mezzo
di fasci paralleli di
microtubuli che
decorrono per
l’intera lunghezza
Essi sono sia
elementi
strutturali sia
elementi utilizzati
come binari lungo
i quali i motori
molecolari
trasportano
proteine ed altri
materiali
dinamicità
Cambiamenti nell’organizzazione
del citoscheletro sono in
relazione a cambiamenti nella
forma della
Cellula
La formazione di filamenti proteici
da subunità proteiche molto più
piccole permette l’assemblaggio e
il disassemblaggio regolato per
dare nuove forme al citoscheletro
Figure 16-7 Molecular Biology of the Cell (© Garland Science 2008)
Quando la cellula si divide si ha un
cambiamento della sua morfologia dovuto
soprattutto al fatto che i microtubuli
subiscono un cambiamento drastico
dell’organizzazione, con una completa
riorganizzazione
(vedi immagine successiva): nel corso della
divisione cellulare (mitosi) la rete di microtubuli
posseduti da una cellula si dissocia
completamente e si riorganizza nella costituzione
del fuso mitotico, che è la struttura che permette
durante la divisione cellulare la segregazione dei
cromatidi
*
microtubuli cromosomi centrioli
*
Cellula prima
della divisione
Quando la cellula si divide non si
ha un riarrangiamento solo dei
microtubuli (verdi), ma anche dei
microfilamenti (filamenti di
actina) (rossi); questi formano
un anello contrattile al centro
della cellula, che divide la cellula
in due dopo che i cromosomi si
sono separati. Una volta divise
le due cellule figlie riorganizzano
sia il citoscheletro di actina che
quello dei microtubuli
Fuso mitotico
Anello
contrattile
cromosomi
Cellule figlie
Figure 16-2 Molecular Biology of the Cell (© Garland Science 2008)
Organizzazione del
citoscheletro in cellule
epiteliali polarizzate
Tutti i componenti del citoscheletro
cooperano per produrre le
caratteristiche forme delle cellule
specializzate, incluse le cellule
epiteliali che rivestono l’intestino
tenue. Alla superficie apicale
(superiore), che si affaccia al lume
intestinale, fasci di filamenti di actina
(rossi) formano microvilli; appena
sotto ai microvilli una banda di
filamenti di actina che corre lungo la
circonferenza della cellula
contribuisce a formare giunzioni
cellula-cellula. Filamenti intermedi
(blu) sono ancorati ad altri tipi di
strutture adesive (desmosomi ed
emidesmosomi) che connettono le
cellule epiteliali in modo da formare un
robusto foglio e le attaccano alla
matrice extracellulare sottostante al
lato basale della cellula. I microtubuli
(verdi) corrono verticalmente
dall’apice alla base della cellula e
forniscono un sistema coordinato che
permette alla cellula di dirigere
componenti di nuova sintesi verso le
loro porzioni appropriate.
Figure 16-5 Molecular Biology of the Cell (© Garland Science 2008)
2)
Nella cellula il mantenimento della
localizzazione delle strutture cellulari è
importante così come il loro spostamento
verso specifici siti intracellulari ; ciò è garantito
dal citoscheletro, un insieme di proteine che
formano dei binari del sistema intracellulare di
trasporto e che agiscono insieme ai motori
proteici che si spostano lungo di essi
Possiamo quindi dire che il citoscheletro non
determina solo la forma della cellula ma
anche organizza la sua struttura interna
stabilendo l’organizzazione generale del
citoplasma
*
Alcuni esempi di spostamenti di organelli, vescicole,
cromosomi (mediate dal citoscheletro), che devono avvenire
per permettere il funzionamento della cellula:
1) Le vescicole secretorie devono lasciare l’apparato del
Golgi, che si trova in prossimità del centro della cellula, e
venire poi trasportate verso la membrana plasmatica dove
svuotano il loro contenuto nello spazio extracellulare (vedi
immagini precedenti su apparato di Golgi).
2) D’altro canto le vescicole portate all’interno della cellula a
livello della membrana plasmatica devono venire
trasportate verso gli endosomi (vedi argomento svolto
precedentemente)
3) I mitocondri si spostano costantemente nella cellula
4) Nel corso della vita di una cellula il reticolo endoplasmatico
si distende e si riorganizza
5) Durante la divisione cellulare i cromosomi si allineano e poi
si muovono verso i poli opposti della cellula
Lo spostamento dei cromosomi, delle vescicole e degli organelli viene effettuato
al momento giusto dal citoscheletro
*
3)
Il citoscheletro permette inoltre il
movimento delle cellule
Molti tipi di cellule si muovono o all’interno
del corpo (cellule animali) o attraverso
l’ambiente (organismi unicellulari ed alcuni
gameti)
Ad esempio i globuli bianchi che inseguono
e distruggono i batteri patogeni strisciando
lungo una superficie
Altre cellule, come gli spermatozoi nuotano
attraverso un fluido per raggiungere la loro
destinazione.
Il citoscheletro guida queste forme di
motilità cellulare
*
Il movimento è in relazione ai filamenti di
actina o microfilamenti, i quali, oltre ad
attraversare la cellula, si estendono in
strutture specializzate che protrudono
dalla superficie e permettono alla cellula
di muoversi
*
Filamenti di actina sul bordo di un fibroblasto permettono al
fibroblasto di muoversi
Per riassumere
La cellula eucariote ha un complesso di strutture
fibrillari che costituiscono una sorta di “ossatura”
interna (citoscheletro)
IL citoscheletro permette alla cellula di:
Mantenere o modificare la propria forma
Migrare
Mantenere gli organelli nelle sedi appropriate
Muovere gli organelli e le vescicoleverso siti specifici
della cellula
Dividersi e segregare in maniera corretta i cromosomi
alla mitosi
Come fa il citoscheletro a garantire queste funzioni?
E’ necessario analizzare la sua struttura per capire la
funzione
*
Le principali strutture del Citoscheletro
Microtubuli
Filamenti di Actina
Filamenti intermedi
*
*
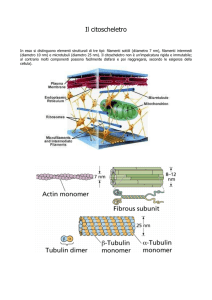
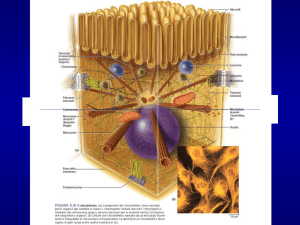
Microtubuli
Strutture tubulari cave (diam.25 nm)
Polimeri del dimero α-tubulina+β tubulina
Strutture rigide
Si organizzano attorno ad un MTOC (microtubules
organizing center) nel centrosoma
Costituiscono le fibre del fuso mitotico, i neurotubuli
dell’assone dei neuroni, ciglia e flagelli (9 coppie di
microtubuli esterni più una coppia centrale, 9+2)
Si degradano e si ricostituiscono velocemente
(INSTABILITA’ DINAMICA)
Ogni microtubulo presenta una estremità positiva, che si
ritiene porti la tubulina β ed una negativa con la tubulina
α. Il microtubulo possiede quindi una polarità
Un singolo microtubulo può contenere
decine o centinaia di migliaia di molecole di
tubulina e può estendersi per molti
micrometri occupando più della metà della
lunghezza di una cellula
Le cellule in interfase contengono centinaia
di lunghi microtubuli che attraversano tutto il
citoplasma
*
In particolare i microtubuli sono polimeri
polari di α e β tubulina
Tredici catene lineari di
subunità,chiamate protofilamenti, si
associano lateralmente formando un
microtubulo
I microtubuli sono polimeri polarizzati:
l’estremità + è formata da subunità di β
tubulina e si assembla più velocemente
mentre la subunità – è costituita da subunità
di α tubulina e si assembla più lentamente
*
I filamenti formati da protofilamenti multipli
hanno proprietà vantaggiose (vedi immagine
successiva)
Figure 16-8 (part 1 of 2) Molecular Biology of the Cell (© Garland Science 2008)
Questa organizzazione del microtubulo
conferisce al microtubulo stesso due
proprietà
1) Le due estremità di un microtubulo
sono strutturalmente diverse e
possono comportarsi in modo diverso
2) Ciascun microtubulo ha una polarità
ossia una direzione e può essere
considerato rivolto verso un punto e
verso un altro, pertanto i microtubuli
rappresentano delle corsie direzionali
per i motori proteici (vedi più avanti);
tale polarità è quindi fondamentale per
organizzare l’ambiente interno cellulare *
L’assemblaggio e il disassemblaggio dei
microtubuli avvengono attraverso un
processo unico chiamato instabilità
dinamica
I microtubuli si alternano costantemente tra
la fase di allungamento e quella di
accorciamento; una estremità del
microtubulo subisce episodi alternati di
crescita e di accorciamento
A seconda delle necessità le cellule
possono rendere i microtubuli più o meno
dinamici
*
tutti i microtubuli sono disposti con le
loro estremità (-) situate in prossimità del
centro della cellula e le loro estremità (+)
poste vicino alla periferia
i microtubuli sono orientati in modo parallelo e
decorrono dalla parte superiore della cellula a
quella inferiore tutti con le estremità (-) rivolte verso
l’alto e le estremità (+) rivolte verso il basso
tutti i microtubuli di un assone puntano in una
stessa direzione
Nell’ambito di ogni specifica cellula tutti i microtubuli hanno la
stessa polarità, così la polarità di un microtubulo può essere
utilizzata come un preciso indicatore citoplasmatico di direzione *
La disposizione dei microtubuli in vari tipi di cellule
In un fibroblasto (o altri tipi di cellule a disposizione
radiale dei microtubuli) tutti i microtubuli sono disposti
con le loro estremità (-) situate in prossimità del centro
della cellula e le loro estremità (+) poste vicino alla
periferia (vedi immagine precedente)
Nelle cellule epiteliali i microtubuli sono orientati in
modo parallelo e decorrono dalla parte superiore della
cellula a quella inferiore tutti con le estremità (-) rivolte
verso l’alto e le estremità (+) rivolte verso il basso (vedi
immagine precedente). In modo analogo tutti i
microtubuli di un assone puntano in una stessa
direzione (vedi immagine precedente)
*
La polarità dei microtubuli è essenziale per
l’organizzazione e funzione della cellula.
Se prendo in considerazione un punto qualsiasi
della superficie di un microtubulo, è possibile
stabilire quale direzione porta all’estremità (+) e
quale invece conduce all’estremità (-). I
microtubuli rappresentano così delle corsie
direzionali per i motori proteici (vedi avanti)
*
Quasi sempre i microtubuli funzionano
insieme ai motori proteici, che generano forze
e spostano vescicole e altri complessi lungo la
superficie dei microtubuli
Le ciglia ed i flagelli sono organelli specializzati,
formati da microtubuli e motori proteici, che fanno
avanzare la cellula attraverso un fluido o
muovono un fluido sulla superficie cellulare
*
I motori proteici associati ai microtubuli
-Quasi tutte le funzioni cellulari che dipendono dai microtubuli
richiedono dei motori proteici ad essi associati
-I motori proteici sono enzimi che generano una forza e
“camminano” lungo i microtubuli verso le estremità (+) o (–) del
microtubulo.
-I motori proteici hanno il dominio della “testa” che si lega al
microtubulo e genera una forza ed il dominio della “coda” che si
lega alla membrana di una vescicola o di un organello o ad un altro
carico di trasporto
Tra i motori proteici ricordiamo:
le chinesine che “camminano” verso l’estremità (+) dei microtubuli
le dineine che “camminano” verso l’estremità (-) dei microtubuli
Il movimento è orientato quindi in una direzione precisa
Esistono anche motori proteici che si associano ai filamenti di actina,
un altro componente del citoscheletro dotato di polarità(vedi più
avanti); i filamenti intermedi, invece, (terzo componente del
citoscheletro), sono privi di polarità e non sono stati identificati motori
proteici che utilizzano i filamenti intermedi come piste per il proprio
spostamento
I motori proteici interagiscono con i
microtubuli, (ma anche con i microfilamenti
, vedi piu’ avanti) utilizzando ATP. Essi
traducono l’energia derivata dall’idrolisi di
ATP in una forza che permette il movimento
*
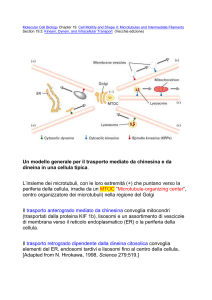
Il motore piu’ conosciuto che agisce a livello
di microtubuli e’ la chinesina
La chinesina è importante per il posizionamento
del Reticolo Endoplasmatico all’interno della
cellula; il RE si estende dalle aree comunicanti con
l’involucro nucleare ed arriva quasi al limite della
cellula raggiungendo la membrana plasmatica;
esso deve ai microtubuli e alle chinesine questa
sua collocazione
Le chinesine si attaccano all’esterno della
sua membrana e lo tirano lungo i
microtubuli in direzione centrifuga
distendendolo come una rete
*
Le Dineine, invece, sospingono lungo i
microtubuli l’apparato del Golgi verso il
centro della cellula
L’apparato del Golgi si trova, infatti, molto
all’interno
In questo modo si creano delle differenze di
localizzazione tra i differenti organelli ed
anche su questo si basa l’efficacia del loro
funzionamento
*
Modalità di funzionamento della chinesina per il movimento delle vescicole o degli
organelli (per es. il reticolo endoplasmatico)
F:\Cell Biology Interactive\media\animations\16.7kinesin.mov
*
Le cellule hanno bisogno di spostare “carichi”
diversi verso specifiche regioni citoplasmatiche
Come vengono indirizzati questi “carichi” verso
specifiche regioni del citoplasma?
“Il carico” si deve accoppiare con lo specifico motore proteico. Il
legame del carico con il motore proteico è mediato dal dominio
della coda del motore proteico. Nel caso ad esempio dei motori
appartenenti alla famiglia delle chinesine i domini della coda
sono diversi da proteina a proteina e rendono ciascun motore
proteico unico.
In genere il dominio della coda non si attacca proprio
direttamente al carico, ma c’è una specifica proteina adattatrice
che si lega sia al “carico “ da trasportare sia alla coda del
motore
In questo modo i domini della coda dei motori proteici
sono come dei rimorchi specifici che vengono caricati da
specifici carichi
Come si originano i microtubuli?
Le cellule utilizzano i centri organizzatori dei
microtubuli per nucleare i microtubuli
• I centri organizzatori dei microtubuli (MTOC) nucleano i
microtubuli all’interno delle cellule
• La posizione dei MTOC determina l’organizzazione dei
microtubuli all’interno delle cellule
• Il centro di organizzazione dei microtubuli piu’ comune
nelle cellule animali è il centrosoma
• I centrosomi sono formato da una coppia di centrioli
circondati dalla matrice pericentriolare
• La matrice pericentriolare contiene γ tubulina; è la γ
tubulina che nuclea i microtubuli agendo in un
complesso formato da parecchie altre proteine
• Le cellule animali mobili contengono un secondo centro
di organizzazione dei microtubuli detto corpo basale
Il centro di organizzazione dei microtubuli
• I microtubuli si
organizzano
attorno al MTOC
che controlla la
nucleazione e la
crescita orientata
dei microtubuli
*
*
*
I centrioli
Sono posizionati vicino al MTOC nel centrosoma
“immersi “ in una matrice proteica
Sono piccoli organelli a forma di barile disposti
ad angolo retto l’uno rispetto all’altro nel centro
del centrosoma
Sono costituiti da particolari strutture di
microtubuli chiamate triplette di microtubuli
9delle quali sono disposte simmetricamente in
modo da formare le pareti del barile (oltre alla a
e β tubulina, i centrioli contengono anche la
tubulina δ e ε
*
Durante ogni ciclo cellulare i centrosomi si
duplicano in preparazione della mitosi.Innanzi
tutto avviene la duplicazione dei centrioli che è
contemporanea alla duplicazione del DNA Il
centriolo si duplica prima della Mitosi e
probabilmente è connesso con la segregazione
dei cromosomi e la corretta divisione cellulare
(vedi argomento Mitosi)
*
Microfilamenti (Filamenti di
Actina)
filamenti ubiquitari
resistenti e flessibili
costituiti da catene di actina avvolte fra loro a
spirale
Distribuiti al di sotto della membrana
plasmatica (cortex cellulare)
Formano strutture caratteristiche come:
l’anello contrattile nella divisione cellulare ( in
particolare nella citodieresi o divisione del
citoplasma); le miofibrille delle cellule
muscolari; i microvilli che formano l’orletto a
spazzola delle cellule intestinali
Le cellule sono capaci di notevole motilità, ad
esempio durante lo sviluppo embrionale nei vertebrati
alcune abbandonano il sistema nervoso in sviluppo e
migrano per tutto l’embrione dando origine a formazioni
diverse come le cellule pigmentate della cute, i denti, la
cartilagine delle mascelle; i leucociti perlustrano i tessuti
del corpo alla ricerca di detriti o microorganismi.
D’altra parte il movimento puo’ anche
interessare solo alcune specifiche parti di
una cellula, ad esempio alcune parti di una cellula
possono espandersi come nel caso degli assoni che
estendono i propri margini ed esplorano il substrato
guidando la cellula verso una sinapsi.Tutti questi
processi di motilità dipendono dalla presenza
di microfilamenti
I microfilamenti sono anche coinvolti nei processi
mobili all’interno della cellula come
nei movimenti delle vescicole nel traffico
vescicolare, nella fagocitosi, nella citocinesi o
citodieresi (divisione del citoplasma)
I microfilamenti hanno un diametro di 8 nm e sono
composti di subunità globulari della proteina actina;
in presenza di ATP i monomeri di actina
polimerizzano formando un filamento flessibile
Un filamento di actina è sostanzialmente una
struttura a doppio filamento
I termini “filamento di actina” “microfilamento” e “F
actina” sono tutti sinonimi
*
Poiché ogni subunità di actina (G actina) ha una
sua polarità e tutte le subunità di un filamento
sono orientate nella stessa direzione, l’intero
microfilamento risulta avere una sua polarità
*
I filamenti di actina formano
diverse strutture cellulari
I filamenti di actina formano i
microvilli dell’orletto a
spazzola delle cellule
intestinali, le stereociglia
dell’orecchio interno, i
lamellipodi, i filopodi
(estroflessioni citoplasmatiche
di cellule migranti),i coni di
accrescimento dei neuroni,le
fibre da stress(fasci contrattili
di actina in cellule non
muscolari) e i sarcomeri
A differenza di quanto
avviene per i microtubuli
dove i protofilamenti si
associano in senso
trasversale direttamente fra
loro, i filamenti di actina non
si associano direttamente tra
loro in senso trasversale, ma
utilizzano vari tipi di proteine,
che si legano all’actina
regolando la
polimerizzazione e
l’organizzazione dell’actina
Le proteine che
formano legami
crociati interagiscono
con i filamenti di
actina formando fasci
di filamenti paralleli o
reticoli
Il motore proteico dei filamenti di actina è
la MIOSINA
• Formata da 2 catene pesanti e 4 catene leggere
• Le catene pesanti sono formate da TESTA globulare e CODA lunga
• A livello della coda le 2 catene pesanti si intrecciano in una struttura
coiled-coil
• Le teste contengono:
testa
– Il sito di legame e idrolisi di ATP
– Il sito di legame per actina
coda
*
La miosina e il movimento dei
microfilamenti
• In tutte le cellule i filamenti di actina interagiscono
con alcune proteine (MIOSINE) per generare forze di
movimento
• Si conoscono 7 classi diverse di Miosine
*
• Tutte possono legare actina e idrolizzare ATP
– Il legame sequenziale di miosina a singole
subunità di actina nei filamenti promuove lo
scorrimento della miosina lungo il filamento di
actina
– Poiché i filamenti di actina sono polarizzati si
generano forze di contrazione nella cellula
• La maggior parte di queste forze di contrazione è
transitoria (es. durante la separazione delle cellule
figlie dopo la mitosi)
• Nei tessuti muscolari specializzati per la contrazione
il sistema dei filamenti di actina e miosina è
altamente specializzato
*
La Miosina II
e la
contrattilità
muscolare
Un muscolo
scheletrico
consiste di fibre
muscolari
(cellule) che
contengono
miofibrille
disposte
longitudinalmen
te; le miofibrille
sono gli
apparati
contrattili e
sono costituiti
da unità
ripetute
chiamate
sarcomeri
Le estremità di
ciascun sarcomero
sono definite da un
disco Z a cui i
filamenti di ACTINA
si attaccano tramite
una proteina (CapZ);
i filamenti spessi di
MIOSINA sono
connessi ai dischi Z
tramite un’altra
proteina (titina) e si
intercalano con i
filamenti di actina
La contrazione
muscolare avviene
quando i filamenti
spessi di miosina si
attaccano ai filamenti
di actina e li
trascinano in modo
da avvicinare i dischi
Z, diminuendo la
lunghezza dei
sarcomeri
*
Riepilogo sulle funzioni dei
microfilamenti
1. Movimenti di trasporto e di contrazione. In
tutte le cellule i filamenti di actina interagiscono
con le miosine generando forze motrici
•
•
Cellule muscolari: contrazione
Altre cellule: trasporto vescicole e organelli sui
microfilamenti
*
2. Strato corticale
• I microfilamenti insieme ad altre proteine,
formano uno strato sotto la membrana
plasmatica che costituisce una rete
resistente a forze deformanti
• Le rete permette variazioni di forma della
cellula mediante modificazioni promosse
da proteine che tagliano i microfilamenti
(es. gelsolina)
• Implicato in numerosi processi
(endocitosi, esocitosi, contrazione dei
microvilli, migrazione cellulare)
3. Supporto meccanico alla membrana
plasmatica mediante proteine di
ancoraggio (es. spectrine e anchirina dei
globuli rossi)
4. Formano strutture rigide di
stabilizzazione per i microvilli. Nei
microvilli l’actina si associa a piccole
proteine di collegamento (fimbrina, fascina,
α-actinina)
5. Responsabili delle forze che controllano
la migrazione cellulare: protrusione locale
di citoplasma evidenti nelle cellule migranti
(fillopodi, lamellipodi)
*
Interazioni dell’actina
L’actina si lega ad una
grande varietà di
proteine accessorie in
tutte le cellule
eucariotiche. Questo
schema mostra la
maggior parte delle
interazioni che sono
state dimostrate,
usando tecniche
genetiche o
biochimiche nel lievito
Saccharomyces
cerevisiae. Le proteine
accessorie che
operano nello stesso
processo intracellulare
sono mostrate nello
stesso colore, come
indicato nella legenda
Figure 16-18 Molecular Biology of the Cell (© Garland Science 2008)
La Falloidina, tossina
dell’Amanita
Phalloides, si lega e
stabilizza i monomeri
di actina causando un
netto decremento
della
polimerizzazione dei
microfilamenti e di
fatto la progressiva
perdita dell’intera
funzione
citoscheletrica
*
FILAMENTI INTERMEDI
classe molto eterogenea di filamenti
a differenza delle altre strutture citoscheletriche sono
stabili
In genere sono costituite da subunità proteiche fibrose
ad α-elica
• Sono un sistema diverso dai microtubuli e
microfilamenti
• Hanno un diametro Ø 10 nm
• Sono formati da fibre proteiche resistenti e
durevoli
• Sono particolarmente presenti nelle cellule
soggette a stress meccanici: epiteli, cellule
nervose (assoni), cellule muscolari
• Sono ancorati alla membrana plasmatica in siti
specifici (desmosomi e emidesmosomi)
• I filamenti intermedi possono essere
citoplasmatici o nucleari
• Citoplasmatici: Cheratine negli epiteli;
Vimentina e Vimentino-simili nel tessuto
connettivo, muscolare e neurogliale
Neurofilamenti, nelle cellule nervose
• Nucleari: le lamine nucleari in tutte le
cellule animali
• Si possono anche trovare nelle
connessioni intercellulari (desmosomi), nel
tessuto epiteliale: Caderine
*
• Filamenti di cheratina:
– Presenti in tutte le cellule epiteliali
– Eterogenei (composti da cheratine diverse:
acide, basiche, neutre), con funzioni simili
– Partono da regione nucleare e
raggiungono le membrana plasmatica dove
si associano a strutture specializzate:
Desmosomi (aree di giunzione tra cellule
adiacenti)
– Funzioni: difesa contro abrasioni, perdita di
acqua, perdita di calore
*
• Filamenti di vimentina e desmina:
– Presenti in molte cellule diverse di origine
mesenchimale
– Desmina: abbondante nelle cellule
muscolari dove forma strutture
specializzate (linee Z-muscolo scheletrico,
dischi intercalari-muscolo cardiaco, corpi
densi-muscolo liscio)
– Funzione dei filamenti di desmina:
ancoraggio dei filamenti contrattili di actina
legandoli alla membrana plasmatica e
fissandoli in registro nei sarcomeri
*
• Neurofilamenti:
– Citoscheletro dei neuroni è costituito da
microtubuli e neurofilamenti
– I neurofilamenti determinano la forma
allungata dei neuriti e dendriti e li
rinforzano
*
Un forte filamento formato da subunità fibrose allungate con forti contatti
laterali
I filamenti intermedi si formano in questo modo e sono di conseguenza
particolarmente resistenti a forze di stiramento, sebbene si pieghino facilmente
Proprietà simili a
quelle di una corda
Figure 16-9 Molecular Biology of the Cell (© Garland Science 2008)
Sostanze che interferiscono con i filamenti di
actina e con i microtubuli
Table 16-2 Molecular Biology of the Cell (© Garland Science 2008)
Matrice Extracellulare (MEC)
Negli eucarioti pluricellulari le cellule dei vari
tessuti secernono sostanze che vanno a
costituire la MEC. Essa ha una importante
funzione di sostegno, soprattutto nel tessuto
osseo dove appare calcificata e rigida e nel
tessuto cartilagineo. La MEC influisce
anche sulla flessibilità dei tessuti e sui
meccanismi di adesione intercellulari.
La MEC è costituita da: Collageni,
Proteoglicani,Glicoproteine di
collegamento
*
Collageni
Componenti preponderanti della MEC.
Elaborati da cellule dei tessuti connettivi
Una molecola di collagene è costituita da
tre catene polipeptidiche ad alto
contenuto di glicina (catene α), avvolte
ad elica fra loro e tenute insieme da
legami a ponte H.
Varie molecole di collagene si associano e
formano una fibrilla.
Più fibrille costituiscono una fibra di
collagene dotate di resistenza
meccanica
Le catene α sono sintetizzate nel RER e portano
amminoacidi aggiuntivi alle estremità. Le catene
α formano triple eliche (procollagene). Esse
tramite traffico vescicolare arrivano al Golgi, lo
attraversano e sono poi introdotte in vescicole di
secrezione e secrete per esocitosi; nello spazio
intercellulare il procollagene tramite rimozione
degli amminoacidi aggiuntivi diviene collagene;
esso si associa in fibrille e poi in fibre
*
Proteoglicani
Le fibre di collagene sono avvolte da una matrice
costituita in prevalenza da proteoglicani, glicoproteine
che risultano da una proteina di peso molecolare molto
vario (da 10 000 a 500 000 dalton), che costituisce
l’asse della molecola (core), cui sono legate catene
laterali di carboidrati detti glicosaminoglicani (GAG); essi
sono in media costituiti da 800 residui di monosaccaridi.
Il peso del proteoglicano può superare il milione di
dalton. I proteoglicani si legano alle fibre del collagene
costituendo il complesso fibroreticolare della MEC. I
proteoglicani sono in grado di trattenere elevate quantità
di acqua e da questa caratteristica dipende la
compattezza ed elasticità del tessuto cartilagineo.
Glicoproteine di collegamento
Esse presentano una struttura a domini multipli e collegano il
complesso fibroreticolare della MEC alle cellule a livello della
membrana plasmatica direttamente tramite i proteoglicani o tramite
recettori specifici situati sulla membrana plasmatica. Le glicoproteine di
collegamento comprendono le fibronectine e le laminine. Le
fibronectine sono presenti nel tessuto connettivo e nel circolo
sanguigno essendo implicate nel processo di coagulazione del sangue.
Le laminine sono costituite da tre polipoeptidi e si trovano soprattutto a
livello della lamina basale, una sottile struttura extracellulare che si
trova tra le cellule epiteliali e il tessuto connettivo sottostante. Le
laminine possono così svolgere un ruolo di molecole di ancoraggio
delle cellule suddette alla lamina basale. Le proteine di collegamento si
possono legare alle cellule per la presenza sulla membrana plasmatica
di recettori transmembrana detti integrine . Dal lato citoplasmatico
della membrana le integrine si legano alle strutture del citoscheletro,
mentre all’estremità opposta si legano ad una molecola della MEC
come il collagene, la laminina o la fibronectina. Le integrine svolgono
importanti ruoli nell’ancoraggio, nella migrazione, nella motalità
cellulare
")