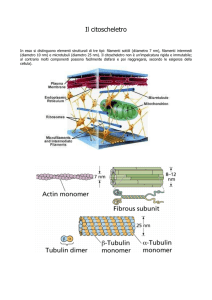
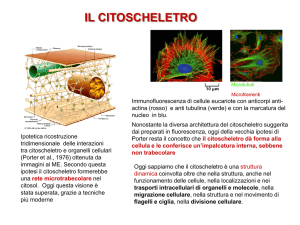
Il CITOSCHELETRO
¾ fornisce una struttura architettonica alle cellule
ma, per la sua natura dinamica, svolge un ruolo
fondamentale:
¾ nei processi di movimento e di divisione cellulare
¾ nel posizionare e muovere gli organelli collegati alle
membrane all’interno del citosol
¾ è anche strettamente associato con i processi di
segnalazione e di adesione cellulare
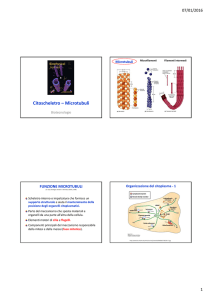
PRINCIPALI ELEMENTI STRUTTURALI DEL CITOSCHELETRO
I vari componenti del
citoscheletro sono collegati
tra loro sia strutturalmente
che funzionalmente.
Cono di
accrescimento
TECNICHE DI STUDIO DEL CITOSCHELETRO
Tecniche fluorescenti in cellule
vive
Videomicroscopia
-Microtubuli assonemali ( strutture stabili,
associate a ciglia, flagelli e corpi basali)
-Microtubuli
citoplasmatici
(strutture
dinamiche, che intervengono in una varietà
di funzioni cellulari come la preservazione
dell’assone, fuso mitotico e meiotico,
movimento
organuli
e
vescicole,
localizzazione AG e RE)
I dimeri di tubulina sono orientati nella
stessa direzione
Origine dei microtubuli: studi in vitro
Nucleazione
Allungamento
+GTP
+ ioni Mg2+
37°C
La polimerizzazione può essere misurata allo spettrofotometro come un
aumento della deviazione del fascio luminoso
veloce
La crescita dei microtubuli in vitro dipende
in modo lineare dalla concentrazione dei
dimeri di tubulina: a elevate concentrazioni
di dimeri i microtubuli crescono, a basse
concentrazioni si disorganizzano.
Esiste un valore di concentrazione di
tubulina in cui il processo di crescita è
controbilanciato
da
quello
di
disorganizzazione, condizione definita come
concentrazione critica.
L’aggiunta di tubulina avviene più
velocemente all’estremità + dei
microtubuli
La diversa velocità di crescita alle estremità
+ e – dei m. riflette le diverse concentrazioni
critiche richieste per l’assemblaggio alle due
estremità:
Bassa per l’estremità +
Alta per l’estremità -
Quando la concentrazione di tubulina libera è maggiore di quella
critica all’estremità +, ma minore di quella critica per l’estremità
-, allora il microtubulo cresce all’estremità + mentre si depolarizza
all’estremità -
La simultanea crescita e depolimerizzazione producono un fenomeno chiamato
TREADMILLING: si verifica quando ogni molecola di tubulina incorporata
all’estremità positiva si sposta lungo il microtubulo in crescita e si stacca
dall’estremità opposta
Per spiegare come i processi di polimerizzazione e
depolimerizzazione possono avvenire
contemporaneamente, Tim Mitchison e Marc
Kirschner hanno proposto il modello dell’instabilità
dinamica dei microtubuli
L’instabilità dinamica dei microtubuli avviene anche in vivo
MTOC: il centro organizzatore dei microtubuli ha la funzione di nucleare e ancorare i
microtubuli. Anche il corpo basale (alla base delle ciglia) serve da MTOC
pericentrina
pericentrina
Tubulina β
Tubulina γ
Fibroblasto in divisione
Gli MTOC organizzano e polarizzano i microtubuli all’interno delle cellule
Il cinetocore si lega all’estremità + del microtubulo
Il fatto che i microtubuli nelle cellule siano soggetti ad
instabilità dinamica giustifica la distribuzione casuale di
microtubuli con un ciclo di vita breve, ma non la
presenza di microtubuli organizzati in modo stabile
all’interno delle cellule.
Un modo per stabilizzare i microtubuli potrebbe essere
quello di “mascherare”, e quindi, proteggere le
estremità positive in crescita.
Es. cinetocore dei cromosomi nella mitosi e corteccia
cellulare, che contengono proteine di attracco
all’estremità +.
Le proteine associate ai microtubuli (MAP)
Alcune MAP si legano ad intervalli regolari lungo la parete del
microtubulo, proiettandosi al di fuori della parete stessa e
permettendo quindi l’interazione del microtubulo con altri
filamenti e strutture cellulari.
Le MAP sono importanti nel regolare l’assemblaggio dei
microtubuli e, legandosi alle estremità positive in crescita di un
microtubulo, ne favoriscono la crescita o ne aumentano la
stabilità.
Le MAP dei neuroni appartengono a due classi:
MAP MOTRICI (chinesina, dineina)
MAP NON MOTRICI (controllano l’organizzazione dei
microtubuli nel citoplasma)
Tau (assone): verde
MAP2 (dendriti): arancio
La funzione di alcune MAP può essere modificata dalla
fosforilazione.
Es. un livello anormalmente alto di fosforilazione della
proteina Tau sembra correlato con lo sviluppo di
diverse malattie neurodegenerative fatali, in cui le
cellule nervose contengono grovigli neurofibrillari
formati da molecole di tau eccessivamente fosforilate ed
incapaci di legarsi ai microtubuli.