ITTIOPATOLOGIA, 2011, 8: 53-62
Esposizione sperimentale a nonilfenolo di giovanili
di tinca (Tinca tinca, Linnaeus 1758): rilievi istologici
e determinazione dei livelli tiroidei
Experimental exposure of juvenile tench
(Tinca tinca, Linnaeus 1758) to nonylphenol: histological
evaluation and thyroid hormone levels determination
Rubina Sirri*, Luciana Mandrioli, Oliviero Mordenti,
Albamaria Parmeggiani, Dino Scaravelli, Annalisa Zaccaroni
Dipartimento di Scienze Mediche Veterinarie, Facoltà di Medicina Veterinaria, Alma Mater Studiorum Bologna,
Via Tolara di Sopra, 50 - 40064 Ozzano Emilia (BO).
______________________________
RIASSUNTO - Lo scopo del presente studio è stato valutare in giovanili di tinca (Tinca tinca) gli effetti del
4-nonilfenolo (NP), sostanza avente azione estrogeno-simile e classificata come interferente endocrino tiroideo. I
soggetti, di circa 120 giorni post-schiusa e 500 mg di peso medio, sono stati esposti per 45 giorni a 25 e 100 µg/l
NP in acqua. Sono state eseguite indagini istomorfologiche su tiroide, fegato e gonadi e sono stati determinati i
livelli di triiodotironina (T3) e tiroxina (T4). In tutti i gruppi nelle sezioni istologiche i follicoli tiroidei si
mostravano rivestiti da epitelio cubico basso e ripieni di colloide ed erano quindi riconducibili a follicoli normali;
nei gruppi trattati vi era una maggior prevalenza di follicoli di piccole e medie dimensioni. Nel fegato la
degenerazione idropica ha mostrato aumento della gravità in tutti i gruppi all’avanzare del periodo di esposizione,
ma è tendenzialmente regredita durante il periodo di sospensione del trattamento. Nel tessuto gonadico non sono
state riscontrate alterazioni istomorfologiche. Tutti i gruppi hanno evidenziato un calo dei livelli di T3
all’avanzare del periodo di esposizione; il gruppo 25 µg/l NP in particolare ha mostrato un drastico calo, mentre il
gruppo 100 µg/l NP ha presentato un andamento opposto rispetto agli altri gruppi, con un picco nel punto di
campionamento a 45 giorni di esposizione. Il T4 ha mostrato un andamento analogo al T3, ma il calo è apparso più
marcato e precoce nel gruppo 100 µg/l NP. I risultati, ancorché preliminari, evidenziano che il NP sembra
interferire con il metabolismo degli ormoni tiroidei, modificandone il bilancio.
SUMMARY - The purpose of this study was to assess the effects of 4-nonylphenol (NP), a substance with
estrogen-like action and classified as endocrine disruptor for thyroid, in juvenile tench (Tinca tinca). Subjects,
approximately 120 days post hatch and 500 mg mean weight, were exposed for 45 days to 25 and 100 µg/l NP in
water. Histological investigations were performed on liver, thyroid and gonads. The levels of triiodothyronine (T3)
and thyroxine (T4) were determined. In all groups the histological sections showed normal thyroid follicles lined
by a flattened epithelial layer and filled with colloid; in treated groups, there was a prevalence of small and
medium follicles. Hepatic hydropic degeneration showed an increase in severity in all groups directly
proportional to the exposure period, but it tends to decline during the suspension period. Gonadal tissue did not
show histological alterations. All groups revealed a decrease of T3 levels directly proportional to the exposure
period; the 25 µg/l NP group in particular showed a marked decline, while the 100 µg/l NP group showed an
opposite trend compared to other groups, with a peak at 45 days of exposure. The T4 level showed a similar trend
to T3, but the decline was more pronounced in the 100 µg/l NP group. The results, although preliminary, show
that NP seems to interfere with thyroid hormone metabolism, modifying its balance.
Key words: Nonylphenol; Environmental contaminant; Endocrine disruptor; Thyroid hormones; Histology;
Tench; Juvenile; Fish; Teleost.
______________________________
* Corresponding Author: c/o Servizio di Anatomia Patologica Veterinaria, Dipartimento di Scienze Mediche
Veterinarie, Facoltà di Medicina Veterinaria - Alma Mater Studiorum, Bologna, Via Tolara di Sopra, 50 - 40064
Ozzano Emilia (BO). Tel.: 051-2097972; Fax: 051-2097968; E-mail: [email protected].
53
ITTIOPATOLOGIA, 2011, 8: 53-62
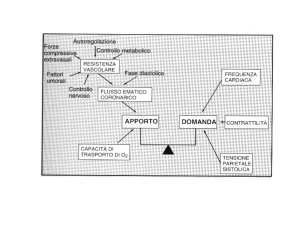
INTRODUZIONE
Gli ormoni tiroidei svolgono un ruolo essenziale nella regolazione dello sviluppo
embrionale, del metabolismo e della riproduzione. Nei teleostei, al pari degli altri vertebrati,
il controllo della secrezione tiroidea è regolato dall'ormone tireostimolante (TSH), che
determina un rilascio in circolo prevalentemente di tiroxina (T4); i livelli di triiodotironina
(T3) sono invece generalmente bassi e la conversione del T4 avviene principalmente a livello
periferico (Eales et al., 1999). Ruolo fondamentale nell’omeostasi degli ormoni tiroidei è
quindi svolto dagli enzimi deiodinasi, che sono responsabili della conversione del T4 in T3.
L’alterazione del metabolismo tiroideo conseguente ad esposizione a contaminanti
ambientali può manifestarsi a livello di sintesi, regolazione, metabolismo e attività degli
ormoni tiroidei (Brown et al., 2004). In particolare, gli interferenti endocrini ambientali nelle
acque possono comportare gravi alterazioni dello sviluppo larvale. Uno dei più comuni di
questi composti è il nonilfenolo (NP), sostanza nota per la sua azione estrogeno-simile e
dotato di attività di interferente endocrino tiroideo (Zaccaroni et al., 2009). Il NP proviene
dalla decomposizione del nonilfenolo etossilato (NPE), tensioattivo non ionico ampiamente
usato in pesticidi, spermicidi, vernici, agenti chimici, tessuti, materiale plastico e prodotti
dell’industria cartaria. Poiché la trasformazione del NPE a NP avviene ad opera dei
microrganismi nei processi di degradazione, queste due molecole arrivano all’ambiente
acquatico tramite i reflui derivanti da impianti di depurazione delle acque o da scarichi
fognari, oppure a causa del run-off dei terreni trattati con pesticidi. Inoltre, il NP, essendo
lipofilo e persistente, tende ad accumularsi negli organismi acquatici come crostacei,
molluschi e pesci, ma anche nel sedimento e nel particolato (John et al., 2000).
Questa molecola è stata ampiamente studiata come responsabile degli effetti che esercita
sulle gonadi e sul fegato; dati riguardanti gli effetti del NP sul bilancio degli ormoni tiroidei
sono invece scarsi e contrastanti (Ishihara et al., 2003; Yamauchi et al., 2003; Schmutzler
et al., 2004; Ghisari & Bonefeld-Jorgensen, 2005; Keen et al., 2005; McCormick et al.,
2005; Razia et al., 2006). Sperimentazioni in vivo hanno dimostrato che il NP può inibire
l’attività della perossidasi tiroidea che catalizza la iodinazione di T3 e T4 in ratti
ovariectomizzati (Schmutzler et al., 2004). Keen et al. (2005), hanno evidenziato come
l’alimentazione di salmonidi con 4-NP durante la smoltificazione non abbia influenzato
significativamente i livelli plasmatici dei due ormoni tiroidei. Al contrario, McCormick et al.
(2005), hanno dimostrato che l’esposizione intraperitoneale a NP nel salmone atlantico è in
grado di indurre una diminuzione dei livelli di T4, senza produrre effetto sulle concentrazioni
di T3. Considerando i molteplici ruoli svolti dagli ormoni tiroidei nella fisiologia dei pesci
(riproduzione, crescita e regolazione della metamorfosi), lo scopo del presente studio è stato
finalizzato alla valutazione degli effetti del 4-NP su tiroide, gonadi e fegato e alla
determinazione dei livelli di ormoni tiroidei in giovanili di tinca (Tinca tinca), specie
frequentemente usata come modello animale in studi tossicologici e metabolici (Gad, 2007).
MATERIALI E METODI
Ottocento soggetti, di circa 120 giorni post-schiusa e 500 mg di peso medio, sono stati
esposti per 45 giorni a 25 e 100 µg/l NP in acqua. Le concentrazioni utilizzate sono state
scelte, sulla base delle informazioni esistenti in letteratura, in modo da ricadere oltre la soglia
di effetto, senza per altro risultare acutamente tossiche per i pesci (Brian et al., 2005; Meucci
& Arukwe, 2005). Sono stati aggiunti un gruppo di controllo negativo (solvente etanolo) e
uno di controllo positivo (propiltiouracile, PTU). Venticinque esemplari per gruppo sono
stati prelevati ogni 15 giorni.
54
ITTIOPATOLOGIA, 2011, 8: 53-62
I rimanenti sono stati mantenuti in acqua pulita per ulteriori 15 giorni e sottoposti alle
medesime indagini. Ad ogni campionamento sono stati registrati peso e lunghezza di ogni
singolo esemplare, per il calcolo dell’indice di condizione k (peso (gr) x 100/lunghezza
(mm)3). Per l’istologia i soggetti sono stati fissati in toto in formalina tamponata al 10% e
processati secondo la metodica istologica di routine; le sezioni istologiche così ottenute,
tagliate al microtomo ad uno spessore di 3 µm, sono state colorate con Ematossilina-Eosina.
Durante l’osservazione dei preparati istologici sono stati contati i follicoli tiroidei e al
numero totale di follicoli tiroidei presenti nella sezione è stato attribuito un punteggio
crescente di 1, 2 o 3, in base al diametro. Gli epatociti sono stati valutati per la presenza di
lesioni degenerative a cui è stato attribuito un punteggio crescente (0, 1, 2), in base alla
gravità. Le gonadi sono state esaminate per eventuali lesioni degenerative e/o anomalie di
sviluppo. Le analisi dei livelli di ormoni tiroidei (T3 e T4) sono state eseguite su omogenato
dell’intero animale applicando la metodica di Crane et al. (2004) modificata (l’omogenato è
stato estratto con miscela di etanolo al 95% contenente 1 mM di PTU). Il dosaggio degli
ormoni tiroidei è stato effettuato con la tecnica del dosaggio radioimmunologico (RIA)
utilizzando kit del commercio (DRG International). L’analisi statistica dei dati è stata
condotta sia sui dati biometrici che sui risultati analitici (concentrazioni di T3 e T4) e
istologici. I confronti relativi all’indice k sono stati eseguiti per ogni punto sperimentale
utilizzando il test t di Student, mentre per gli altri parametri è stata applicata un’analisi della
varianza (ANOVA). Infine, relativamente al punteggio del diametro dei follicoli tiroidei, i
dati ottenuti sono stati valutati mediante analisi delle componenti principali (PCA).
RISULTATI
Il calcolo dell’indice di condizione k ha evidenziato nel gruppo di controllo un valore k
notevolmente inferiore rispetto a quello dei gruppi trattati solo a 15 giorni di trattamento
(Figura 1).
Figura 1 – Il calcolo dell’indice di condizione K ha evidenziato come il gruppo di controllo presenti
un valore K notevolmente inferiore (p<0,01) solo a quello dei gruppi trattati per 15 giorni. Ai successivi
punti di campionamento i valori dell’indice si equiparano tra i vari gruppi e non è più possibile evidenziare
differenze statisticamente significative.
Figure 1 – Condition factor K calculation showed a significantly lower value (p<0.01) in the control group
with respect to groups treated for 15 days. In the other sampling points the K values are similar among
the various groups and did not show statistically significant differences.
55
ITTIOPATOLOGIA, 2011, 8: 53-62
Ai successivi punti di campionamento i valori dell’indice si equiparano tra i vari gruppi e
non è più possibile evidenziare differenze statisticamente significative. Nelle sezioni
istologiche di tiroide di soggetti sottoposti a 15, 30 e 45 giorni di trattamento in tutti i gruppi
i follicoli tiroidei erano normali, mostrandosi rivestiti da epitelio cubico basso e ripieni di
colloide (Figura 2).
56
ITTIOPATOLOGIA, 2011, 8: 53-62
Figura 2 – Tiroide. Sezioni istologiche di soggetti sottoposti a 15 giorni (A, etanolo; B, 25 µg/l NP; C, PTU; D,
100 µg/l NP) e 45 giorni di trattamento (E, etanolo; F, 25 µg/l NP; G, PTU; H, 100 µg/l NP). In tutti i gruppi i
follicoli si mostravano rivestiti da epitelio cubico basso e ripieni di colloide (Ematossilina-Eosina, obbiettivo
40x).
Figure 2 – Thyroid. Histological sections of subjects treated for 15 days (A, ethanol; B, 25 µg/l NP; C, PTU; D,
100 µg/l NP) and 45 days (E, ethanol; F, 25 µg/l NP; G, PTU; H, 100 µg/l NP). In all groups the follicles were
lined by a flattened epithelium and filled with colloid (Hematoxylin-Eosin, 40x lens).
L’analisi statistica condotta tramite PCA sul punteggio conferito al diametro dei follicoli
tiroidei e al numero di follicoli presenti nella sezione ha evidenziato nel punteggio totale una
maggior presenza di follicoli di piccole e medie dimensioni (punteggio 1 e 2) nei gruppi 25
µg/l NP e 100 µg/l NP (Figura 3).
Standardized Biplot (t1 vs. t2)
5.00
-5.00
4.00
0.00
3.00
punteggio
grandezza 2
35.00
6.00
-5.00
30.00
5.00 25.00
0.00
t2
4.00
20.00 2.00
5.00
3.00
15.00
10.00
15.00
5.00
10.00
2.00
20.00
25.00
5.00
n.follicoli nel campo
1.00
5.00
1.00
0.00
0.00
-5.00
10.00
0.00
15.00
-1.00
grandezza 1
grandezza 3
t1
Figura 3 – L’analisi statistica condotta tramite PCA sul punteggio conferito al diametro dei follicoli tiroidei e al
numero di follicoli presenti nella sezione ha evidenziato nel punteggio totale una maggior presenza di follicoli di
piccole e medie dimensioni (punteggio 1 e 2) nei gruppi 25 µg/l NP e 100 µg/l NP.
Figure 3 – Statistical analysis conducted by PCA on the score given to the diameter of thyroid follicles and on the
number of follicles in the section showed in the total score more follicles of small and medium size (score 1 and 2)
in groups 25 µg/l NP and 100 µg/l NP.
57
ITTIOPATOLOGIA, 2011, 8: 53-62
L’analisi del punteggio conferito alla degenerazione idropica epatica ha mostrato un
aumento della gravità in tutti i gruppi all’avanzare del periodo di esposizione; nei gruppi 100
µg/l NP e PTU il punteggio era più alto e quindi la degenerazione più grave. Tuttavia la
degenerazione idropica, ad eccezione del gruppo PTU, tendeva a regredire durante il periodo
di sospensione del trattamento, compatibilmente con il carattere di reversibilità di questo
processo patologico (Figura 4).
Figura 4 – L’analisi del punteggio conferito alla degenerazione idropica epatica ha mostrato un aumento
della gravità in tutti i gruppi all’avanzare del periodo di esposizione; nei gruppi 100 µg/l NP e PTU
il punteggio era più alto e quindi la degenerazione più grave. Tuttavia, ad eccezione del gruppo PTU,
il processo degenerativo tendeva a regredire durante il periodo di sospensione del trattamento.
Figure 4 – The analysis of the score given to the hepatic hydropic degeneration showed an increase in severity
in all groups directly proportional to the exposure period; in 100 µg/l NP and PTU groups the score was highest
and the degeneration more severe. However, with the exception of PTU group, the degenerative process tended
to decline during the suspension period.
Figura 5 – Tutti i gruppi hanno evidenziato un calo dei livelli di T3 all’avanzare del periodo di esposizione; il
gruppo 25 µg/l NP in particolare ha mostrato un drastico calo, mentre il gruppo 100 µg/l NP ha presentato un
andamento opposto rispetto agli altri gruppi, con un picco nel punto di campionamento a 45 giorni di esposizione.
Figure 5 – All groups revealed a decrease in levels of T3 directly proportional to the exposure period; in
particular the 25 µg/l NP group showed a marked decline, while the 100 µg/l NP group presented an opposite
trend compared to other groups, with a peak at 45 days of exposure.
58
ITTIOPATOLOGIA, 2011, 8: 53-62
Nel tessuto gonadico non sono state riscontrate alterazioni istologiche. Tutti i gruppi hanno
evidenziato un calo dei livelli di T3 all’avanzare del periodo di esposizione; il gruppo 25 µg/l
NP in particolare ha mostrato un drastico calo, mentre il gruppo 100 µg/l NP ha presentato
un andamento opposto rispetto agli altri gruppi, con un picco nel punto di campionamento a
45 giorni di esposizione (Figura 5). Il T4 ha mostrato un andamento analogo al T3, ma il calo
è apparso più marcato e precoce nel gruppo 100 µg/l NP (Figura 6).
Figura 6 – Il T4 ha mostrato un andamento analogo al T3 ma il calo è apparso più marcato
e precoce nel gruppo 100 µg/l NP.
Figure 6 – The T4 showed a similar trend to T3 but the decline appeared more pronounced
in the 100 µg/l NP group.
DISCUSSIONE E CONCLUSIONI
I risultati, ancorché preliminari, evidenziano che il NP sembra interferire con il
metabolismo degli ormoni tiroidei, modificandone il bilancio. Il gruppo esposto a più alto
dosaggio ha mostrato infatti un picco del livello di T3 a 45 giorni di trattamento che, messo
in relazione con il drastico calo mostrato dal T4 a 30 giorni potrebbe rappresentare l’effetto di
un blocco della sintesi del T4 da parte della tiroide e di un conseguente aumento nella
trasformazione di quest’ultimo a T3 da parte delle deiodinasi periferiche, al fine di sopperire
al fabbisogno tissutale. La forma biologicamente attiva è infatti quella del T3, che si forma
dalla deiodinazione della T4 a livello periferico da parte degli enzimi deiodinasi. Gli ormoni
tiroidei sono coinvolti nei processi di sviluppo larvale in molte specie di teleostei,
intervenendo in particolare nella metamorfosi post-schiusa. Per questo motivo gli stadi
larvali e giovanili sono considerati molto più sensibili alla presenza di sostanze tossiche
ambientali rispetto agli adulti. In base a queste considerazioni assume un particolare
significato valutare gli effetti di molecole con ruolo di interferenti endocrini tiroidei nelle
prime fasi di vita dei pesci (Crane et al., 2004; Norris, 2007).
59
ITTIOPATOLOGIA, 2011, 8: 53-62
In questo studio, l’andamento del T3 nel gruppo a più alto dosaggio potrebbe essere anche
l’espressione di un’alterazione nell’accrescimento/metamorfosi dei giovanili di tinca
provocato da un’interferenza con la sintesi/metabolismo degli ormoni tiroidei.
La valutazione istologica del tessuto tiroideo è considerata uno strumento utile
all’identificazione di fenomeni patologici correlati con l’alterazione della funzionalità della
ghiandola tiroidea. Quando la tiroide è sottoposta a stimolazione è possibile riscontrare una
serie di modificazioni istomorfologiche dose-dipendenti come aumento dell’altezza delle
cellule epiteliali, incremento della vascolarizzazione, iperplasia dei follicoli, irregolarità del
contorno dei follicoli e diminuzione della densità della colloide, accompagnata da numerose
gocce endocitotiche di colloide ai bordi apicali delle cellule epiteliali dove i microvilli
diventano più abbondanti (Eales & Brown, 1993).
Con la presente sperimentazione è stato possibile contribuire all’approfondimento dello
studio degli aspetti istomorfologici della tiroide nei pesci ossei, un tessuto di non semplice
reperimento durante il campionamento in esemplari adulti e neppure in preparati istologici di
larve o giovanili. Dati bibliografici descrivono patologie tiroidee per lo più di tipo
proliferativo (Fournie et al., 2005). Nei pesci ossei la tiroide raramente è circoscritta da una
capsula connettivale, pertanto i follicoli si rinvengono distribuiti in modo sparso nel tessuto
connettivo della regione faringea, in vicinanza dell’aorta ventrale o della parete della cavità
branchiale posteriore e sono spesso numerosi in prossimità della prima e della seconda
arteria branchiale. Probabilmente proprio a causa della mancanza della capsula connettivale,
i follicoli tiroidei possono mostrare una distribuzione ectopica e localizzarsi pertanto nella
tonaca vascolare oculare, nel rene, nella milza, nell’intestino, nel fegato, nel cuore e in altri
tessuti. La presenza di tessuto tiroideo ectopico privo di connotazione patologica è stata
riportata in numerose specie ittiche, compreso il carassio, il pesce sciabola, la trota ed il
barbo. All’istologia i singoli follicoli tiroidei sono simili a quelli degli altri vertebrati, sono
solitamente rotondi od ovali e bordati da un singolo strato di epitelio che può essere
squamoso o cubico, a seconda del livello dell’attività metabolica della ghiandola (Fournie
et al., 2005). Il centro di ciascun follicolo contiene il secreto costituito dalla colloide,
rappresentato da proteine leganti gli ormoni tiroidei; la quantità di colloide varia a seconda
dell’attività metabolica dei follicoli e del grado di produzione degli ormoni. La fisiologica
deplezione di colloide, che prende avvio dalla periferia del follicolo, corrisponde
istologicamente alla comparsa di vacuoli chiari e rappresenta l’espressione dell’attività
metabolica della ghiandola (Leatherland, 1994). Le cellule parafollicolari (cellule C o cellule
chiare) non sono presenti nella tiroide dei pesci (Fournie et al., 2005).
In questo studio sperimentale condotto su giovanili di tinca i follicoli tiroidei sembravano
tendere complessivamente ad una riduzione del diametro, mentre non si sono evidenziati
fenomeni di ipertrofia-iperplasia dell’epitelio follicolare e/o una riduzione significativa della
colloide. La riduzione del diametro dei follicoli tiroidei, maggiormente evidente nel gruppo a
più alto dosaggio, potrebbe essere il risultato di una diminuita capacità biosintetica, che
confermerebbe il drastico calo della concentrazione di T4.
Poiché lo scopo della ricerca era valutare il possibile effetto del NP sul sistema endocrino
in giovanili in accrescimento riproducendo una esposizione ambientale, si è deciso di non
determinare i livelli residuali di NP nei tessuti.
E’ pertanto possibile concludere che, in questo studio sperimentale, l’esposizione a NP ha
determinato un rallentamento dell’attività biosintetica dei follicoli tiroidei (visibile attraverso
il drastico calo di T4), con una parallela maggior attivazione delle deiodinasi periferiche, che
per il periodo di esposizione e le concentrazioni utilizzate andrebbero considerate alla luce di
una “manovra“ omeostatica dell’organismo, volta a compensare le richieste di giovanili in
rapido accrescimento.
60
ITTIOPATOLOGIA, 2011, 8: 53-62
La presenza in questa specie, come in altre specie di teleostei, di follicoli tiroidei diffusi
rende difficoltosa una valutazione istopatologica complessiva rispetto ad una ghiandola in
toto. Ulteriori ricerche sono necessarie per stabilire se l’attività di interferente endocrino del
NP sia legata semplicemente al rallentamento dell’attività biosintetica o se un maggior
periodo di esposizione o una più alta concentrazione siano in grado di incidere
irreversibilmente sull’attività tiroidea.
BIBLIOGRAFIA
Brian J.V., Harris C.A., Scholze M., Backhaus T., Booy P., Lamoree M., Pojana G., Jonkers N.,
Runnalls T., Bonfà A., Marcomini A. & Sumpter J.P. (2005). Accurate prediction of the response of
freshwater fish to a mixture of estrogenic chemicals. Environ. Health Perspect., 113: 721-728.
Brown S.B., Adams B.A., Cyr D.G. & Eales J.G. (2004). Contaminants effects on the teleost fish
thyroid. Environ. Toxicol. Chem., 23: 1680-1701.
Crane H.M., Pickford D.B., Hutchinson T.H. & Brown J.A. (2004). Developmental changes of thyroid
hormones in the fathead minnow, Pimephales promelas. Gen. Comp. Endocrinol., 139: 55-60.
Eales J.G. & Brown S.B. (1993). Measurement and regulation of thyroidal status in teleost fish. Rev.
Fish Biol. Fisher., 3: 299-347.
Eales J.G., Brown S.B., Cyr D.G., Adams B.A. & Finnson K.R. (1999). Deiodination as an index of
chemical disruption of thyroid hormone homeostasis and thyroidal status. In: “Environmental
toxicology and risk assessment: standardization of biomarkers for endocrine disruption and
environmental assessment: eighth volume”, Henshel D., Balck M.C., Harrass M.C. (Eds), ASTM STP
1364, American Society for Testing and Materials, West Conshohocken.
Fournie J.W., Wolfe M.J., Wolf J.C., Courtney L.A., Johnson R.D. & Hawkins W.E. (2005).
Diagnostic criteria for proliferative thyroid lesions in bony fishes. Toxicol. Pathol., 33: 540-551.
Gad S.C. (2007). Alternative species. In: “Animal models in toxicology. 2th Ed.”, CRC Press, Boca
Raton.
Ghisari M. & Bonefeld-Jorgensen E.C. (2005). Impact of environmental chemicals on the thyroid
hormone function in pituitary rat GH3 cells. Mol. Cell. Endocrinol., 244: 31-41.
Ishihara A., Nishiyama N., Sugiyama S. & Yamauchi K. (2003). The effect of endocrine disrupting
chemicals on thyroid hormone binding to Japanese quail transthyretin and thyroid hormone receptor.
Gen. Comp. Endocrinol., 134: 36-43.
John D.M., House W.A. & White G.F. (2000). Environmental fate of nonylphenol ethoxylates:
differential absorption of homologs to components of river sediment. Environ. Toxicol. Chem., 19:
293-300.
Keen P.L., Higgs D.A., Hall K.J. & Ikonomou M. (2005). Effects of dietary exposure of 4nonylphenol on growth and smoltification of juvenile Coho salmon (Oncorhynchus kisutch). Sci. Total
Environ., 349: 81-94.
Leatherland J.F. (1994). Reflections on the thyroidology of fishes: from molecules to humankind.
Guelph Ichthyol. Rev., 2: 1-67.
61
ITTIOPATOLOGIA, 2011, 8: 53-62
McCormick S.D., O'Dea M.F., Moeckel A.M., Lerner D.T. & Björnsson B.T. (2005). Endocrine
disruption of parr-smolt transformation and seawater tolerance of Atlantic salmon by 4-nonylphenol
and 17[beta]-estradiol. Gen. Comp. Endocrinol., 142: 280-288.
Meucci V. & Arukwe A. (2005). Detection of vitellogenin and zona radiata protein expressions in
surface mucus of immature juvenile Atlantic salmon (Salmo salar) exposed to waterborne
nonylphenol. Aquat. Toxicol., 73: 1-10.
Norris D.O. (2007). The hypothalamus-pituitary-thyroid (HTP) axis of non-mammalian vertebrates.
In: “Vertebrate endocrinology. 4th Ed.”, Academic Press, New York.
Razia S., Maegawa Y., Tamotsu S. & Oishi T. (2006). Histological changes in immune and endocrine
organs of quail embryos: exposure to estrogen and nonylphenol. Ecotoxicol. Environ. Saf., 65: 364371.
Schmutzler C., Hamann I., Hofmann P.J., Kovacs G., Stemmler L., Mentrup B., Schomburg L.,
Ambrugger P., Grüters A., Seidlova-Wuttke D., Jarry H., Wuttke W. & Köhrle J. (2004). Endocrine
active compounds affect thyrotropin and thyroid hormone levels in serum as well as endpoints of
thyroid hormone action in liver, heart and kidney. Toxicology, 205: 95-102.
Yamauchi K., Ishihara A., Fukazawa H. & Terao Y. (2003). Competitive interactions of chlorinated
phenol compounds with 3,3′,5-triiodothyronine binding to transthyretin: detection of possible thyroiddisrupting chemicals in environmental waste water. Toxicol. Appl. Pharmacol., 187: 110-117.
Zaccaroni A., Gamberoni M., Mandrioli L., Sirri R., Mordenti O., Scaravelli D., Sarli G.
& Parmeggiani A. (2009). Thyroid hormones as a potential early biomarker of exposure to
4-nonylphenol in adult male shubunkins (Carassius auratus). Sci. Total Environ., 407: 3301-3306.
62