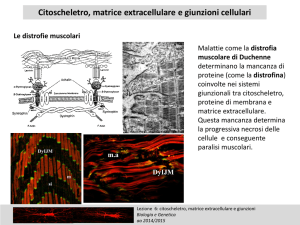
Esiste una relazione tra morfologia e funzione
cellulare
La forma di una cellula è
altamente indicativa della
sua funzione e della sua
“normalità”
La morfologia delle cellule è
strettamente collegata ad un
complesso di fibre che si
estende nel citoplasma e che
si proietta lungo tutte le
estroflessioni della membrana
plasmatica:
il CITOSCHELETRO
Il citoscheletro è formato da una complessa rete di filamenti
proteici che si estende dal nucleo alla membrana
attraverso il citosol
Il citoscheletro è presente solo nelle cellule eucariotiche
Una cellula in ipossia o con
blocco della fosforilazione
ossidativa assume
morfologia tondeggiante
per depolimerizzazione del
citoscheletro.
forma una struttura di sostegno che determina, oltre che la
forma della cellula, l’organizzazione del citoplasma
(posizionamento e distribuzione degli organuli cellulari,
compreso il nucleo)
FUNZIONI DEL CITOSCHELETRO
contribuisce al mantenimento dell’integrità dei contatti
cellula-cellula
è responsabile dei movimenti cellulari
è coinvolto nell’ Organogenesi e nell’ Istogenesi (Migrazione
cellule embrionali, Orientamento).
è determinante nella divisione cellulare (formazione del fuso
mitotico e citodieresi)
I tre principali elementi strutturali del citoscheletro
Microtubuli
Microfilamenti
Actina: 6 nm – Miosina: 15 nm
Filamenti intermedi
8-11 nm
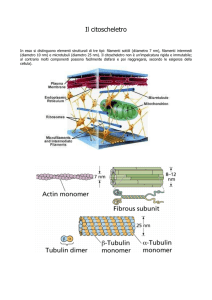
MICROTUBULI
● Organizzazione e mantenimento della forma della cellula
● Disposizione e movimento degli organelli
●Trasporto vescicolare
● Movimento di cromosomi alla divisione cellulare
● Motilità cellulare tramite appendici (ciglia e flagelli)
Eterodimero
di tubulina
Struttura
Tubo cavo con una parete formata da 13 protofilamenti
Diametro
Esterno 25 nm - Interno 15 nm
Subunità
α-tubulina e β-tubulina
L’ α-tubulina è associata a GTP, la β-tubulina al GDP (che
viene scambiato con GTP prima dell’assemblaggio)
I microtubuli hanno una polarità
La tubulina ha conformazione
globulare, con una tasca per una
molecola di GTD/GDP. I monomeri
contenenti GTP dimerizzano
facilmente (eterodimerizzano).
Nel dimero il GTP della tubulina alfa
è in qualche modo protetto, mentre
quello della beta è rapidamente
idrolizzato in GDP.
La stessa tubulina ha attività
GTPasica.
Una terza isoforma di tubulina, la γ-tubulina costituisce
circa il il 5% di tutta la tubulina, è localizzata
prevalentemente nel centrosoma e nei centrioli ed è
cruciale per l’assemblaggio dei microtubuli.
Negli ultimi anni sono state individuate almeno altre 4
isoforme di tubulina, non presenti in tutte le cellule e dalle
funzioni poco note.
Una preparazione di microtubuli purificati è formata per
l’80-90% da tubulina e per il restante 10-20% da MAP
(proteine associate al microtubulo)
I microtubuli hanno origine dal centrosoma
*
Il centrosoma è una
struttura complessa
che contiene due
centrioli circondati
da materiale
pericentriolare (PCM)
amorfo ed
elettrondenso.e
funziona come Centro
di Organizzazione dei
Microtubuli (MTOC)
*nove fibrille regolarmente spaziate ognuna
delle quali formata, in sezione trasversale,
da una banda di tre microtubuli (A,B e C)
Il centrosoma è
costituito
prevalentemente da
γ-tubulina e da altre
proteine (Spc97 e
Spc98) che insieme
formano il γ-TURC (γtubulin Ring Complex)
Il corretto
assemblaggio del
complesso γ-TURC è
assistito dalla
Pericentrina, una
proteina la cui
concentrazione
fluttua con il ciclo
cellulare
Benché i centrioli non
siano direttamente
coinvolti nella
nucleazione dei
microtubuli, hanno
probabilmente un ruolo
nel reclutare il
PeriCentriolarMaterial
Nella matrice del centrosoma sono presenti molti
complessi γ-TURC, ognuno dei quali funziona come centro
di nucleazione dei microtubuli
L’estremità – dei microtubuli è immersa nella matrice del centrosoma
I microtubuli si rimodellano di continuo in seguito al
verificarsi di cicli di polimerizzazione/depolimerizzazione,
sono cioè dinamicamente instabili
L’instabilità dinamica
è essenziale per i cambiamenti di forma della cellula e per alcuni tipi
di movimenti cellulari
Live-cell imaging: a) le proteine fluorescenti sono sintetizzate come
proteine di fusione contenente GFP
b) le subunità vengono rese fluorescenti in vitro e poi iniettate
nella cellula
L’instabilità dinamica
dei microtubuli risponde alle esigenze della cellula
Distribuzione dei microtubuli in una cellula vegetale durante il ciclo
cellulare: 1) interfase 2) avvicinandosi alla mitosi delineano la fascia
del piano di divisione 3) in mitosi formano il fuso 4) riorganizzazione
della parete delle due cellule figlie
Nel dimero il GTP della tubulina beta è facilmente
idrolizzato in GDP, ma quando la polimerizzazione
è abbastanza veloce (alta concentrazione di
eterodimer isolubili) nuovi dimeri con beta-GTP
vengono reclutati così che l’estremità – contiene
sempre GTP.
Quando la velocità di polimerizzazione diminuisce
il GTP dell’estremità + viene idrolizzato a GDP il
che induce una modifica conformazionale
dell’eterodimero.
L’eterodimero diventa più instabile così come le
interazioni laterali tra protofilamenti.
La conclusione di questa serie di eventi è la
depolimerizzazione del microtubulo
Proteine associate ai microtubuli
La funzionalità dei microtubuli dipende da numerose
proteine accessorie (MAP = Microtubule Associated
Proteins) che svolgono vari compiti:
1) Proteine stabilizzanti i microtubuli
2) Proteine destabilizzanti i microtubuli
3) Proteine motrici dei microtubuli ( chinesine e dineine)
1) Proteine stabilizzanti i microtubuli
La stabilizzazione selettiva dei microtubuli, da parte di
proteine accessorie che si legano alla loro estremità,
determina la polarità cellulare
Altre proteine stabilizzanti si legano alla parete dei
microtubuli difendendoli da attacchi idrolitici e
favorendone l’associazione in fasci.
La proteina Tau è specifica delle cellule nervose.
La sua funzione è legata allo stato di fosforilazione di
suoi specifici residui aminoacidici.
Patologia Umana
Una mutazione* della proteina Tau, è presente in molte
malattie neurodegenerative in cui si osserva accumulo di fasci
anomali di proteina Tau, assemblaggio ridotto e
disorganizzazione dei microtubuli, inibizione del trasporto
anterogrado assonemale, degenerazione neuronale
*la proprietà delle MAP di
legarsi ai microtubui è
controllata dallo stato di
fosforilazione di alcuni residui
aminoacidici. La mutazione della
Tau comporta una
iperfosforilazione della proteina.
2) Proteine destabilizzanti i microtubuli
Tra le proteine destabilizzanti, alcune
sequestrano l’eterodimero/GTP riducendo la sua
concentrazione e quindi la velocità di
polimerizzazione, (alcune di queste proteine sono
iperespresse in certe forme tumorali), altre
hanno la proprietà di frammentare i microtubuli
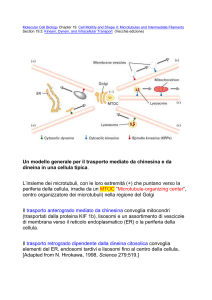
3) Proteine motrici dei microtubuli (chinesine e dineine)
Dineina
Chinesina
La chinesina e la dineina usano l’energia derivante dall’idrolisi
dell’ATP per “camminare” lungo i microtubuli, trasportando
organelli membranosi
Chinesina
Dineina
Le proteine motrici associate ai microtubuli sono
importanti per:
-il trasporto di vescicole
-il trasporto assonale
-il movimento di ciglia e flagelli
Trasporto assonale: la maggior parte del materiale (compresi i
neurotrasmettitori) viene inserito in vescicole membranose. Ma
lungo i microtubuli dell’assone vengono trasportate anche
molecole libere (RNA, ribosomi…). Nei neuromotori il percorso può
coprire anche la distanza di un metro (vel. max 5 μm/sec.)
Movimento assonale anterogrado (chinesine) e
retrogrado (dineine)
Le vescicole trasportate legano sia la
chinesina che la dineina ma a seconda
del trasporto una delle due è
inattivata.
Ambedue le proteine sono
legate alla membrana della
vescicola tramite intermediari:
la chinesina utilizza proteine
integrali o periferiche di
membrana, la dineina proteine
solubili.
Patologia Umana
Tra i carichi mossi dalla dineina troviamo gli endosomi, i
lisosomi, le vescicole derivanti dal RE e il virus dell’HIV che è
trasportato nel nucleo della cellula infettata.
Difetti nel trasporto assonale sia anterogrado che retrogrado
sono stati associati ad alcune malattie neurologiche.
I microtubuli e le proteine motrici si assemblano in una
struttura altamente specializzata nelle ciglia e nei flagelli:
L’ASSONEMA
Assonema = 9+2
9 coppie di mt periferiche e
1 centrale
Nelle coppie periferiche un
mt (A)
completo ed uno addossato
incompleto
Zona transizione= ha
organizzazione
intermedia tra assonema e
corpo basale
Corpo basale= 9 triplette di
mt disposte
a cerchio (mt completi)
ha la stessa struttura di
centriolo (=9
triplette in cerchio);
Patologia Umana
Mutazioni dei geni che codificano per la dineina si associano a
quadri patologici caratterizzati da broncopatie ostruttuive e
sterilità.
Le ciliopatie rappresentano una vasta gamma di gravi patologie
molte delle quali indicative di errori durante lo sviluppo (sono
ciliate le cellule che durante la gastrulazione determinano il
piano di organizzazione di un mammifero).
ESEMPIO: sindrome di KARTAGENER (deficit di dineina)
alterazioni dell’apparato respiratorio, infertilità maschile, situs
viscerum inversum
tre microtubuli costituiscono il fuso mitotico: mt del
cinetocore, mt dei cromosomi, mt polari (non si attaccano
ai cromosomi ma si stabilizzano sovrapponendosi l’un
l’altro).
I mt dell’aster partono dal centrosoma e si dirigono in
periferia; servono ad allontanare i poli del fuso.
I cromosomi si muovono
verso i poli del fuso lungo
i microtubuli del
cinetocore. Il movimento
è guidato da proteine
motrici associate al
cinetocore e dirette
verso l’estremità- del mt.
L’azione di queste
proteine è associata al
disassemblaggio del mt.
FILAMENTI INTERMEDI
● Mantenimento della forma della cellula
● Supporto strutturale e resistenza meccanica
● Formazione dell’impalcatura nucleare
Struttura Subunità fibrose avvolte in un filamento simile a una corda
Diametro
8-12 nm
Subunità
Parecchie proteine diverse tessuto-specifiche
I filamenti intermedi sono costituiti da proteine diverse
nei diversi tipi cellulari
Cheratine
Vimentina e
vimentino-simili
Negli epiteli
Nel tessuto connettivo,
muscolare e neurogliale
Neurofilamenti
Nelle cellule nervose
Lamìne nucleari
In tutte le cellule nucleate
citoplasmatici
nucleari
Le proteine dei filamenti intermedi sono tutte
proteine fibrose e condividono un dominio centrale ad
alfa elica che è conservato per dimensioni, struttura e
in alcuni casi sequenza.
Cheratine: Epiteli
Vimentina: fibroblasti e cell mesenchimali
Nella TRANSIZIONE EPITELIO/MESENCHIMA (EMT)
Scompaiono le cheratine, sostituite dalla vimentina nella stessa cellula
Stabilità dei filamenti intermedi
Regolazione della polimerizzazione dei filamenti intermedi
da parte della fosforilazione (PKA, p38 MAPK)
INIBIZIONE DI p38 MAPK:
Riduzione di citocheratine,
Aumento di espressione di
Vimentina
La diversità dei filamenti intermedi è clinicamente utile nella
diagnosi dei tumori
Man mano che i tumori crescono le caratteristiche
strutturali del tessuto normale scompaiono
Tuttavia, tumori di origine sconosciuta possono essere
diagnosticati mediante l’identificazione dei loro filamenti
intermedi con anticorpi specifici.
Ad esempio, i tumori che esprimono citocheratine possono
essere classificati come carcinomi (di origine epiteliale) e
distinti dai sarcomi (di origine mesenchimale) che esprimono
vimentina.
Disposizione dei filamenti intermedi all’interno della cellula
Filamenti intermedi citoplasmtici
Làmina nucleare
Patologia Umana
Epidermolisi Bollosa
Dovuta ad alterazioni genetiche o acquisite del collagene,
cheratina, laminina, desmocolina, desmoplachina,
desmoleina, placoglobina, placofillina, integrine e altre che
formano desmosomi ed emidesmosomi, strutture proteiche
deputate alla adesività cellulare.
LAMINE NUCLEARI: A, B, C
La lamina B ha un ruolo strutturale e la delezione di questo gene è
Incompatibile con la vita
Mutazioni della lamina Lamina A sono presenti nella Hutchinson–Gilford
progeria syndrome. Gli individui affetti acquisiscono precocemente
caratteristiche senili (scleroderma, aterosclerosi, cecità) sie muoiono in
genere prima dell’adolescenza. Lo studio della progeria è estremamente
interessante perché riguarda lo studio dei meccanismi di invecchiamento
cellulare
Lamine: modificazioni dinamiche del nucleo. Ruolo nell’espressione
genica e nel controllo della forma del nucleo
La regolazione della forma del nucleo operata dalle lamine ha un impatto anche
Nella migrazione cellulare, come evidenziato da saggi di migrazione
Migrazione cellulare su Transwell
Nucelo di soggetto normale
Deficit di LAMINA A
Stimolo chemiotattico




![[ XI ] INDICE GENERALE Riconoscimento e interazioni tra cellule](http://s1.studylibit.com/store/data/001175494_1-9062ea75af318561914dbe68aa0e27ca-300x300.png)