Principali molecole di interesse biologico
Il nostro sistema biologico è basato sul carbonio:
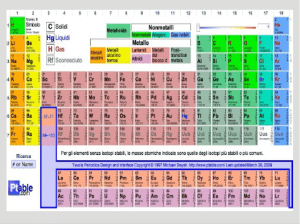
Il carbonio (simbolo C) è un non metallo del gruppo VI A del sistema periodico; è presente in
natura, allo stato elementare, in due differenti forme cristalline: la grafite e il diamante. Nella crosta
terrestre è presente in una piccola percentuale nei carbonati, nel petrolio, nel carbone e nel gas
naturale; nell’atmosfera si trova soprattutto come monossido (CO) e diossido (CO2) di carbonio (o
anidride carbonica), ma la percentuale maggiore di carbonio si trova come costituente delle strutture
di base degli organismi. Il numero dei composti organici supera di gran lunga quello di tutti i
composti formati dagli altri elementi (composti inorganici) messi insieme: il carbonio può essere
considerato il più versatile tra gli elementi chimici grazie a una singolare combinazione di proprietà.
Infatti l’atomo di carbonio:
forma fino a 4 legami covalenti a geometria spaziale tetraedrica;
ha piccole dimensioni;
non ha elevata elettronegatività, che permette la formazione di legami stabili con elementi
come l’idrogeno e l’ossigeno;
forma, con altri atomi di carbonio, catene, corte o assai lunghe, lineari o ramificate, oppure
chiuse ad anello, con una varietà di combinazioni quasi infinita;
forma un elevatissimo numero di isomeri (molecole costituite dagli stessi atomi, che però
sono disposti in modo differente nello spazio).
La Composizione della materia vivente
Tutte le cellule degli organismi viventi sono composte da molecole semplici, come acqua e ioni, e
molecole complesse, dette macromolecole come proteine, carboidrati, acidi nucleici e lipidi.
Acqua
2
H20 70%
Ioni inorganici
Composizione della
materia vivente
i.e. Na+ Cl- K +
1%
Piccole molecole
Amminoacidi
Zuccheri Nucleotidi
Acidi grassi
6%
Macromolecole
DNA
RNA
Proteine
23%
L’IMPORTANAZ DEI LEGAMI CHIMICI
Il tipo di legame chimico che si stabilisce tra atomi, ioni e molecole determina fenomeni diversi
come:
la miscibilità e la solubilità delle sostanze, quando si formano soluzioni,
la conducibilità elettrica, sia delle sostanze che delle soluzioni.
Questo tipo di legame chimico dipende in larga misura dalla polarità delle molecole.
La polarità delle molecole dipende da due fattori:
la polarità dei legami covalenti, all'interno della molecola,
la forma della molecola.
La polarità dei legami covalenti dipende dalla differenza di elettronegatività degli atomi degli
elementi legati (maggiore è la differenza di elettronegatività dei due elementi, maggiore è la polarità
del legame) ; più complessa è la questione della forma della molecola.
Forma delle molecole
La forma di molte molecole può essere spiegata da una teoria indicata con la sigla VSEPR (dalle
iniziali delle parole inglesi ‘Valence–Shell Electron-Pair Repulsion’ che significa: "repulsione
delle coppie di elettroni del guscio di valenza") .
Tale teoria si basa sul principio che gli elettroni che fanno parte dell’ultimo strato - sia le coppie
di elettroni dei legami covalenti, sia eventualmente quelle di elettroni non condivisi - tendono a
respingersi (a causa della loro carica negativa) e si posizionano alla massima distanza possibile fra
loro, formando angoli di legame i più ampi possibile.
Nel metano (CH4), attorno all'atomo centrale di carbonio, ci sono - nello strato di valenza - quattro
coppie di elettroni condivisi con quattro atomi di idrogeno:
Il massimo angolo di legame che si può realizzare tra le quattro coppie di elettroni - in modo che
tutti gli angoli siano uguali - è di 109,5°. Per questo gli atomi di idrogeno si dispongono come ai
vertici di un ideale tetraedro, al centro del quale sta l'atomo di carbonio:
Consideriamo adesso la molecola dell’ammoniaca (NH3). Attorno all’atomo di azoto ci sono tre
coppie di elettroni di legame e una coppia di elettroni non condivisa:
Se confrontiamo questa struttura con quella precedente del metano, vediamo che in entrambe le
molecole c'è un atomo centrale circondato da quattro coppie di elettroni. Ma c'è una differenza: una
coppia di elettroni che prima era condivisa (tra il carbonio e l'idrogeno) ora appartiene solo
all'atomo centrale (l'azoto).
La coppia di elettroni non condivisa dell'azoto - essendo più vicina all'atomo di azoto di quanto non
fosse la coppia di elettroni condivisa tra carbonio e idrogeno nella molecola del metano - esercita
una più forte azione di repulsione sulle coppie di elettroni di legame: ne consegue che l'angolo di
legame diventa di 107,3°, cioè leggermente inferiore a quello di 109,5° del metano, e la molecola
assume una struttura di tipo piramidale (struttura tetraedrica distorta se consideriamo anche la
coppia di elettroni non condivisa):
Nella molecola d’acqua le coppie non condivise sono due:
Le considerazioni da fare sono analoghe a quelle fatte precedentemente.
Rispetto alla molecola dell'ammoniaca c'è una coppia di elettroni non condivisa in più che - essendo
più vicina all'atomo di ossigeno di quanto non fosse la coppia di elettroni condivisa tra azoto e
idrogeno nella molecola dell'ammoniaca - esercita una più forte azione di repulsione sulle coppie di
elettroni di legame: ne consegue che l'angolo di legame diventa di 105°, cioè leggermente inferiore
a quello di 107,3° dell'ammoniaca, e la molecola assume una struttura detta a V o angolata (struttura
tetraedrica distorta se consideriamo anche le coppie di elettroni non condivise):
Una diversa geometria riscontriamo invece in molecole come BF3 e C2H4 (etilene). Il boro forma 3
legami covalenti ma non ha nell’ultimo livello elettroni non condivisi: di conseguenza la geometria
che comporta la massima distanza tra i legami e trigonale planare con angoli di 120°. In C2H4 il
carbonio forma sempre 4 legami ma con 3 atomi , 2 con atomi di idrogeno e uno doppio con un
altro atomo di carbonio: è da prevedere quindi una geometria planare con angoli di 120°. Le misure
sperimentali indicano un angolo tra gli atomi di idrogeno leggermente più piccolo (117°) : ciò
potrebbe essere interpretato attribuendo al doppio legame una azione repulsiva maggiore rispetto al
legame semplice.
Nell’anidride carbonica (CO2) c'è invece un angolo di 180° .
Anche in questo caso la teoria VSEPR spiega la geometria della molecola.
Il carbonio forma due doppi legami covalenti con due atomi di ossigeno e gli elettroni di questi
legami staranno alla massima distanza possibile fra loro, formando appunto un angolo di legame di
180°.
La stessa geometria la ritroviamo nel cloruro di berillio (BeCl2) (molecola in cui si riscontra una
eccezione alla regola dell’ottetto):
Il berillio forma due legami covalenti con il cloro utilizzando i due soli elettroni posseduti
nell’ultimo livello.
Molecole polari e apolari
Una molecola è apolare quando le cariche positive (dei protoni) e negative (degli elettroni) sono
distribuite in modo che non ci sia una parte positiva ed una parte negativa della molecola.
Forse ti stai chiedendo da che cosa dipenda ciò. Ebbene i fattori importanti sono due: la polarità dei
legami covalenti all'interno della molecola e la forma della molecola.
Quando i legami sono apolari la molecola è sicuramente apolare e non c'è bisogno di considerare
la forma della molecola (per es. le molecole degli elementi I2, P4 e S8).
Quando i legami sono polari si possono verificare due casi:
1. la molecola è simmetrica: in questo caso la molecola è apolare (è il caso dell'anidride carbonica,
CO2).
Il carbonio forma due legami covalenti doppi con due atomi di ossigeno e c'è un angolo di legame
di 180°.
Il legame tra C e O è polare( EC = 2,5 ; EO = 3,5 ) , ma i due dipoli sono esattamente opposti e la
molecola risulterà complessivamente apolare.
2. la molecola è asimmetrica: in questo caso la molecola risulta essere polare (è il caso dell'acqua,
H2O).
Il legame tra l’ossigeno e l’idrogeno è polare (EO=3,5 ; EH = 2,1), ciò significa che gli elettroni di
valenza trascorrono più tempo attorno all'atomo di ossigeno piuttosto che attorno agli atomi di
idrogeno. Ciò determina una parziale carica negativa dalla parte dell'ossigeno (a causa della carica
negativa degli elettroni). Per lo stesso motivo la parte degli atomi di idrogeno presenta una parziale
carica positiva). Queste due caricheopposte e dello stesso valore poste ad una certa distanza
costituiscono un dipolo.
In altre parole i due dipoli dei due legami covalenti tra idrogeno e ossigeno si sommano, e la
molecola risulta complessivamente polare.
Un dipolo si rappresenta con una freccia che punta verso la zona negativa della molecola.
L'acqua
L'acqua e' il componente predominante di tutte le cellule. Nell'uomo, il suo contenuto
relativo varia con con l'età: e' massimo nell'embrione (dal 90 al 95%) e diminuisce
progressivamente nell'adulto e nell'anziano, senza scendere, comunque, mai al di sotto dell'80%.
La principale funzione svolta dall'acqua e' quella di solvente, entro il quale le molecole di
importanza biologica possono tra loro interagire nelle varie reazioni chimiche che caratterizzano la
vita della cellula. L'acqua stessa può entrare in molte reazioni chimiche come reagente o derivarne
come prodotto finale ed in essa vengono disciolti molti composti di rifiuto che devono essere
eliminati dalla cellula. L'acqua, inoltre, grazie alla sua capacità termica (elevato calore specifico),
funge da termostato all'interno della cellula, assorbendo calore ed evitando brusche variazioni della
temperatura cellulare.
L'acqua, per la distribuzione asimmetrica delle sue cariche elettriche, e' una molecola polare,
in grado di interagire sia con molecole cariche positivamente che negativamenete (Fig.1).
L'acqua si dissocia in continuazione, in maniera reversibile, in un protone H+ (idrogenione)
ed un anione OH- (ossidrilione). Ad ogni istante solo una piccola frazione di molecole di acqua si
trova nello stato ionizzato: in condizioni normali di temperatura e pressione (25 C al livello del
mare) sono dissociate solamente 1x10-7 moli per litro di acqua e questa concentrazione corrisponde
ad un valore di pH 7, cioe' alla neutralità. La misura della concentrazione degli ioni H+si esprime in
unità di pH. Poiche' il pH e' una grandezza che indica variazioni logaritmiche, una variazione di una
singola unità di pH corrisponde allo spostamento di un ordine di grandezza della concentrazione di
protoni H+ (Fig.2). Nonostante la loro bassissima concentrazione, idrogenioni ed ossidrilioni sono
importantissimi nella cellula, poiche' influenzano tutte le interazioni tra le molecole implicate
nell'attività cellulare.
Gli ioni
Elementi chimici come il sodio, il potassio, il cloro, il fosforo, il calcio, il magnesio, il
manganese, il ferro e altre giuocano un ruolo chiave nello svolgimento di molteplici funzioni
cellulari. Questi elementi, per le peculiarità della loro interazione con l'acqua, si trovano in forma di
ioni, cioe' cariche elettricamente.
La concentrazione relativa dei singoli ioni e' differente all'interno della cellula rispetto
all'esterno e tale differenza di concentrazione viene mantenuta selettivamente da complessi
meccanismi di trasporto a livello della membrana cellulare.
Il sodio (Na+) ed il cloro (Cl-) hanno una localizzazione prevalentemente extra-cellulare e rientrano
nel controllo della pressione osmotica e dell'equilibrio acido/base.
Il potassio (K+) ha una localizzazione prevalentemente intra-cellulare e rientra nel controllo della
pressione osmotica e dell'equilibrio acido/base
Gli ioni fosfato (HPO4--, H2PO4-) costituiscono la specie anionica più abbondante nella cellula,
dove contribuiscono tra l'altro a formare l'adenosina trifosfato (ATP), la principale riserva
energetica della cellula. Nel compartimento extra-cellulare contribuiscono alla regolazione
dell'equilibrio acido/base, stabilizzando il pH del sangue e dei liquidi interstiziale
Il calcio (Ca++) all'interno della cellula entra nel controllo di funzioni specializzate come la
contrazione muscolare e la trasmissione dello stimolo nervoso; all'esterno della cellula, invece, il
calcio si trova principalmente accumulate a livello di ossa e denti
Il Magnesio (Mg++) e il Manganese (Mn++) rientrano come cofattori nell'azione di molteplici
enzimi.
Il Ferro (Fe++/+++), grazie alla sua capacità di cambiare valenza e' l'elemento chiave del trasporto
dell'ossigeno mediante l'emoglobina a tutte le cellule e quindi a tutti i tessuti dell'organismo.
LE MACROMOLECOLE
Acidi nucleici
Gli acidi nucleici sono le macromolecole in cui e' contenuta la informazione genetica, cioe'
le informazioni per la costituzione di tutti i componenti cellulari e per il loro corretto
funzionamento. Conosciamo due specie principali di acidi nucleici, l'acido desossiribonucleico
(DNA) e l'acido ribonucleico (RNA), che sono presenti in tutti gli organismi conosciuti.
Il DNA
Il DNA e' un polimero di nucleotidi ed e' presente in tutte le cellule sotto forma di una
doppia elica costituita da due filamenti anti-paralleli (Fig. 3).
I nucleotidi sono composti da una base azotata che può essere purinica (adenina e guanina) o
pirimidinica (timina e citosina) (FIG.), da uno zucchero a cinque atomi di carbonio, il
desossiribosio, e da un gruppo fosfato (Fig. 4).
I nucleotidi sono concatenati l'uno all'altro da un legame fosfodiesterico tra il gruppo fosfato legato
al carbonio in posizione 3' ed il gruppo ossidrilico legato al carbonio 5' del nucleotide successivo
(Fig. 5).
I nucleotidi si allineano lungo la doppia elica con lo scheletro zucchero-fosfato rivolto all'esterno e
le basi azotate all'interno in posizione ortogonale rispetto allo scheletro zucchero-fosfato. Le due
catene della doppia elica sono tenute insieme da legami idrogeno che si instaurano tra la basi
affacciate secondo la regola della complementarietà: una adenina può legarsi solo ad una timina ed
una citosina solo ad una guanina. Tra adenina e timina si formano due legami idrogeno e tra
guanina e citosina se ne formano tre (fig. 6).
La doppia elica si avvolge attorno ad un asse centrale ideale in direzione destrorsa, formando un
giro completo ogni dieci coppie di basi (3.4 nm).
La doppia elica può essere reversibilmente denaturata (si possono, cioe', separare i due
filamenti) sia con mezzi fisici, l'aumento della temperatura, sia con mezzi chimici, l'aumento della
forza ionica. Poichè tra citosina e guanina vi sono tre legami ad idrogeno, maggiore è il contenuto
di coppie CG maggiore sarà la temperatura necessaria per separare i due filamenti dell'elica. Questa
caratteristica del DNA è stata particolarmente utile per capire alcune caratteristiche del genoma
come, per esempio, la sua dimensione: quanto più grande è il filamento di DNA che si sta
denaturando, tanto maggiore sarà il tempo, necessario per la sua rinaturazione in quanto i due
filamenti si potranno appaiare solo quando avranno trovato gli accoppiamenti giusti in base alla
complementarietà delle basi. Attraverso studi di denaturazione e rinaturazione sono stati messi in
evidenza tratti ripetuti di DNA: quando questi sono presenti la velocità di rinaturazione è di gran
lunga maggiore rispetto a quella che si ha in presenza di sequenze singole.
Sommario del DNA
L'RNA
L'RNA è costituito da un singolo filamento polinuclotidico in cui lo zucchero a cinque atomi di
carbonio è rappresentato da un ribosio e le basi azotate sono, adenina uracile (al posto della timina),
citosina
e
guanina.
Un'altra differenza tra DNA e RNA è che mentre il DNA esegue essenzialmente la funzione di
contenere l'informazione) l’RNA è presente in diverse forme, ognuna specializzata in una
determinata funzione: RNA messaggero (mRNA), RNA di trascrizione (tRNA), RNA ribosomiale
(rRNA). Tutti e tre queste famiglie molecolari sono implicate nella sintesi delle proteine
Principali tipi di RNA
L'RNA messaggero (mRNA) è una molecola prodotta dalla trascrizione del DNA, che porta il codice
genetico sul sito della sintesi proteica
L'RNA transfer (tRNA) è costituito da piccole molecole a forma di trifoglio, ciascuna delle quali porta
uno specifico aminoacido.
L'RNA ribosomiale (rRNA) è il principale costituente dei ribosomi, i microscopici organuli cellulari su
cui avviene la sintesi proteica.
Sommario delle differenze tra DNA ed RNA
Legame fosfodiesterico uguale nel DNA e
nell’RNA
Le Proteine
Le proteine sono polimeri costituiti dalla sequenza lineare di aminoacidi tenuti insieme da
legami peptidici.
posizione della catena laterale
Il nome aminoacido deriva dalla contemporanea presenza nella stessa molecola di un gruppo
aminico (NH2) e di uno carbossilico (COOH).
Gli aminoacidi utilizzati in natura per la costituzione delle proteine sono 20 e tutti condividono la
medesima struttura di base ma sono caratterizzati da un differente gruppo R, detto residuo o catena
laterale. Ad esempio, se e' costituito da un singolo atomo di idrogeno da'
luogo alla glicina, se da un gruppo metile -CH3 dà luogo alla alanina.
La biosintesi degli aminoacidi
Le catene laterali conferiscono caratteristiche peculiari ai singoli aminoacidi:
due hanno
caratteristiche acide, acido aspartico e acido glutammico; tre hanno caratteristiche basiche, lisina,
arginina e istidina; sette hanno caratteristiche neutre e sono polari, cioè idrofili, serina, treonina,
tirosina, triptofano, asparagina, glutamina e cisteina ; otto hanno caratteristiche neutre e sono
apolari, cioè idrofobi, glicina, alanina, valina, leucina, isoleucina, fenilalanina, prolina e metionina.
Inoltre, metionina e cisteina contengono un atomo di zolfo che permette la costituzione di ponti
disolfuro (-S-S-), particolarmente frequenti tra due molecole di cisteina (Fig. 7).
Gli aminoacidi si legano fra di loro con un legame peptidico che si instaura tra il gruppo aminico di
un aminoacido e quello carbossilico dell'aminoacido precedente:
Nel polimero lineare vi è sempre un gruppo aminico ad una estremità (aminoacido N-terminale) ed
un gruppo carbossilico all'altra estremità (aminoacido C-terminale).
Molto spesso alla catena polipeptidica sono legati gruppi non proteici più o meno grandi,
come zuccheri, lipidi o gruppi metallici, che prendono il nome di gruppi prostetici. E' questo il caso
dell'eme, un gruppo metallo-organico contenente il ferro, che nella costituzione dell'
nell'emoglobina per il trasporto dell'ossigeno (Fig. 8).
L’EMOGLOBINA
Le proteine svolgono nella cellula una serie molto diversa di funzioni e questo è dovuto all'enorme
numero di diverse proteine che si possono costituire mescolando variamente i venti aminoacidi, che,
come abbiamo visto, hanno caratteristiche fisico chimiche diverse che vengono trasferte alla
proteina di cui fanno parte o, per lo meno, a segmenti di essa.
Le proteine, come tutte le molecole, tendono ad assumere la conformazione spaziale piu'
vantaggiosa da un punto di vista termodinamico, cioe' quella che consente la migliore distribuzione
dell'energia molecolare in tutta la struttura, evitando cosi' accumuli regionale di cariche sia
elettriche che chimiche. In base proprio a questo principio di termodinamica le proteine assumono
la loro forma definitiva, passando, durante e dopo la loro sintesi, attraverso diversi livelli di
organizzazione spaziale.
Il primo livello è dato dalla sequenza lineare degli amminoacidi ed è detto struttura primaria. Gli
aminoacidi che si allineano lungo la struttura primaria influenzano la forma che la catena
polipeptidica assume nello spazio.
Il secondo livello, detto struttura secondaria rappresenta il primo momento di
organizzazione spaziale di una proteina ed e' reso possibile dalla rotazione del carbonio a, cioe'
quello non coinvolto nel legame peptidico. Il legame peptidico, infatti, e' rigidamente planare
(Fig.9), per cui possono ruotare nello spazio solo gli atomi di carbonio posti ai suoi vertici, cosi' da
consentire la formazione di legami idrogeno e di forze di attrazione e repulsione elettrostatica (forze
di van der Waals) atte a stabilizzare la strutture secondaria. Analizzando la probabilita' di differenti
gradi di rotazione intorno ai legami peptidici su entrambi i lati degli atomi di carbonio a e le
possibilita' per la formazione di legami idrogeno atti a stabilizzare la struttura, i chimici delle
proteine hanno individuato due disposizioni che risultano particolarmente stabili in termini
termodinamici: α elica e struttura b o a foglietto ripiegato (Fig. 10).
Nel caso dell'α elica la porzione polipeptidica coinvolta si avvolge intorno ad un ideale cilindro con
le catene laterali rivolte verso l'esterno; legami idrogeno si instaureranno tra gli atomi di idrogeno e
di ossigeno legati al carbonio ed all'azoto coinvolti nel legame peptidico di amminoacidi che si
susseguono lungo l'elica, mentre tra gli atomi delle catene laterali si instaureranno forze di van der
Waals. La struttura b prevede il ripiegamento a zig zag di singoli segmenti della catena
polipeptidica, detti b strand senza formazione di legami idrogeno all'interno di ciscuna b strand.
Quando le strutture b di una data proteina si trovano allineate l'una affianco all'altra esse sono tenute
in registro da legami idrogeno tra gli atomi di idrogeno e di ossigeno legati al carbonio ed all'azoto
coinvolti nel legame peptidico e formano un piano b o foglio ondulato. Una terza disposizione che
non presenta tendenza alla formazione di a eliche o di b strand, e' detta spirale casuale (random
coil) e fornisce regioni flessibili che permettono alle catene polipeptidiche di curvarsi e di
arrotolarsi. Le porzioni a spirale casuale agiscono frequentemente come cerniere che permettono a
regioni diverse di una proteina di muoversi le une rispetto alle altre.
La struttura terziaria è il frutto dell'interazione nella stessa molecola di zone ad α elica, β strand ed a
spirale casuale grazie all'instaurarsi di legami idrogeno di attrazioni tra gruppi laterali caricati
positivamente e negativamente, di forze di van der Waals, di associazioni polari ed apolari e persino
di legami covalenti. Anche questa struttura è determinata dalla composizione aminoacidica della
catena polipeptidica; infatti, se in una sequenza sono presenti, ad esempio, due cisteine, esse
tendono a formare tra loro un ponte disolfuro inducendo la proteina ad assumere una particolare
forma spaziale.
La struttura quaternaria si riscontra quando una proteina è composta non da una singola catena
polipeptidica ma da più subunità. La funzione della proteina è assicurata dalla corretta relazione
spaziale che le varie subunità prenderanno l'una rispetto alle altre (Fig 11).
Le funzioni delle proteine
Le proteine svolgono una elevata varietà di funzioni e da esse dipendono la maggior parte delle attività
cellulari. La capacità di ogni proteinedi svolgere una determinata funzione è strettamente legata alla sua
struttura.
Si possono riconoscere alcune classi funzionali tra le proteine:
- enzimi, che catalizzano le reazioni metaboliche
- proteine strutturali (ad esempio i filamenti del citoscheletro)
- proteine
regolatrici che agiscono da fattori trascrizionali o controllano i vari processi intracellulari
-proteine in grado di agire come messaggeri intracellulari o come recettori
- proteine che permettono il passaggio e il trasporto di varie molecole
- proteine che svolgono
funzioni di riconoscimento come gli anticorpi
Gli enzimi
Tra le varie funzioni svolte dalle proteine all'interno di una cellula c'è quella enzimatica. Gli enzimi
sono proteine che agiscono accelerando le reazioni chimiche che avvengono in una cellula, ma non
subiscono nessuna modificazione durante questo processo, per cui possono essere usati e riusati
innumerevoli volte. È importante ricordare che un enzima favorisce una reazione che avviene
naturalmente, ma non può far avvenire reazioni che naturalmente non possono avvenire. Queste
molecole proteiche hanno strutture molto diverse per forma e per sequenza primaria, ma hanno
sempre come caratteristica comune: una porzione della molecola che si comporta da sito attivo, cioè
da punto di attacco per il substrato - la sostanza che deve essere sintetizzata o demolita. Gli enzimi
sono caratterizzati dal fatto di avere una altissima specificità per il substrato, cio' significa che un
dato enzima è in grado di catalizzare solo una certa reazione e nessun'altra. Questo garantisce
l'accuratezza necessaria a tutte le reazioni chimiche che avvengono in una cellula. Alcuni enzimi
per svolgere la loro azione catalitica hanno bisogno di un cofattore, un'altra molecola, in genere di
natura non proteica, che favorisce o addirittura permette che l'enzima svolga la sua attività. Il
substrato (S) si lega al sito attivo dell'enzima (E) in maniera transitoria formando un complesso
chiamato ES; una volta che il substrato è stato convertito in uno o più prodotti (P) l'enzima torna
libero (E+S=ES= E+P). Come abbiamo accennato sopra, la specificità dell'enzima per il substrato è
molto elevata al punto che l'enzima è inacapace di agire su composti analoghi al substrato ma di
forma molecolare anche solo leggermente diversa. La teoria che spiega l'alta specificita del attività
enzimatica è basata sul fatto che il centro attivo ed il substrato hanno forme complementari e quindi
solo quando l'incastro tra substrato ed enzima è perfetto può avvenire la reazione enzimaticaca, in
maniera analoga al sistema chiave-serratura dove solo una chiave è capace di aprire una determinata
serratura. Poichè il ruolo di un enzima è quello di aumentare la velocità con cui avviene una certa
reazione e poichè abbiamo detto che l'enzima non partecipa alla reazione stessa, è intuitivo che la
velocità della reazione dipenderà dalla concentrazione del substrato. La curva caratteristica che
rappresenta una reazione enzimatica è un'iperbole e la reazione raggiunge la velocità massima
(Vmax) quando i siti attivi di tutti le molecole di quel determinato enzima sono legate al substrato.
Un altro valore importante in una reazione enzimatica è la costante di Michaelis o Km che
rappresenta la concentrazione di substrato quando la velocità di reazione è la metà della velocità
massima ossia Vmax/2. Da ciò si deriva che quanto più piccola sarà Km tanto più elevata sarà
l'affinità dell'enzima per il suo substrato (Fig. 12). L'attività enzimatica può essere inibita in maniera
reversibile o irreversibile. Una inibizione reversibile può a sua volta essere competitiva o non
competitiva. Una inibizione competitiva si ha in presenza di un inibitore che ha struttura simile a
quella del substrato; la inibizione non competitiva si verifica quando l'inibitore non ha omologie
strutturali con il substrato e quindi interagisce con l'enzima in un sito diverso da quello attivo
compromettendo il legame del substrato al sito attivo.
Una caratteristica fondamentale delle proteine e' la loro capacita' di cambiare conformazione
a secondo dell'ambiente chimico-fisico in cui si trovano o della loro interazione con altre molecole.
Un esempio particolare di ciò sono le proteine allosteriche, a questa categoria appartengono anche
molti enzimi che hanno due siti attivi e quindi specificità per due substrati. In alcuni casi il legame
di un substrato al suo sito specifico provoca una modificazione conformazionale dell'altro sito attivo
impedendo al secondo substrato di legarsi ad esso; in altri casi la modificazione conformazionale
indotta dal legame di un substrato al suo sito specifico e' condizione necessaria perche' il secondo
sito attivo possa riconoscere il suo substrato specifico (Fig. 13).
Acidi grassi e lipidi
I lipidi sono un gruppo di sostanze caratterizzate dalla loro relativa insolubilità in acqua e
dall'elevata solubilità in solventi organici. Questa solubilità è dovuta al fatto che nelle molecole
lipidiche predominano lunghe molecole idrocarburiche o anelli benzoici, strutture entrambe apolari
e idrofobe. I lipidi possono essere divisi in semplici (trigliceridi) e complessi (colesterolo e
fosfolipidi di membrana).
Gli acidi grassi sono costituiti da una molecola di idrocarburo (molecole formate solo da
atomi di carbonio e idrogeno) a lunga catena, alla cui estremità è presente un gruppo carbossilico
COOH:
HOOC-CH2-CH2-CH2-CH2-CH2-CH2-CH2-CH2-CH2-CH2-CH2-CH2-CH2-CH2-CH2-CH2CH3
questo, ad esempio, è l'acido stearico
HOOC-CH2-CH2-CH2-CH2-CH2-CH2-CH2-CH=CH-CH2-CH2-CH2-CH2-CH2-CH2-CH2-CH3
questo è, invece, l'acido oleico.
Nell'acido stearico tutti gli atomi di carbonio della catena, tranne il primo e l'ultimo, sono
legati a due atomi di idrogeno, cioè la molecola è satura di atomi di idrogeno, per cui viene detto
acido grasso saturo.
Nell'acido oleico, invece, gli atomi di carbonio in posizione 9 e 10 sono legati tra loro con
un doppio legame e sono legati ciascuno ad un solo atomo di idrogeno, quindi la molecola non è
satura di idrogeno e viene chiamata acido grasso insaturo.
La presenza di un doppio legame nella struttura di un acido grasso determina delle
modificazioni strutturali nella molecola che non sarà più perfettamente lineare, come nel caso
dell'acido stearico, ma presenterà un gomito (Fig. 14a). La presenza del doppio legame è
importante, in quanto modifica la temperatura di fusione degli acidi grassi, che sono i costituenti
principali della membrana plasmatica. Gli acidi grassi sono prevalentemente apolari e non ionici, di
conseguenza sono insolubili in acqua.
Un ruolo molto importante in varie funzioni cellulari è svolto dai trigliceridi, che sono
costituiti da tre catene di acido grasso saturo legate ad una molecola di glicerolo
TRIGLICERIDI = esteri del glicerolo con 3 acidi grassi
I trigliceridi sono i grassi di deposito del nostro
organismo, accumulati nel tessuto adiposo, dentro gli
adipociti e sono la principale forma di deposito di
energia metabolica del nostro organismo (120000 kcal
totali contro le 1200 del glicogeno).
Il tessuto adiposo è localizzato nel sottocute e intorno
agli organi interni.
L’idrolisi dei trigliceridi ad acidi grassi e glicerolo è catalizzata dagli enzimi lipasi.
I trigliceridi non si sciolgono in acqua (non hanno alcuna parte polare nella molecola), ma
viaggiano nel sangue ricoperti da una guscio proteico, sotto forma di chilomicroni e di particelle
lipoproteiche (VLDV, LDL, HDL).
I trigliceridi (Fig. 14 b)così formati si accumulano nel tessuto adiposo e costituiscono un
vantaggioso deposito di energia di riserva. Infatti, dato il basso stato di ossidazione delle lunghe
catene idrocarburiche, quando i trigliceridi vengono ossidati nelle cellule in CO2 e H2O, liberano
una grande quantità di energia, circa il doppio di calorie per grammo rispetto ai carboidrati ed alle
proteine.
Quando allo scheletro del glicerolo si trovano legate, invece che tre molecole di acido grasso
saturo come nel caso dei trigliceridi, al primo atomo di carbonio un gruppo fosforico ed un alcool
(come la colina, l'etanolammina, l'inositolo o la serina), al secondo una molecola di acido grasso
insaturo ed al terzo una molecola di acido grasso saturo si otterrà un fosfolipide di membrana (Fig.
14 c).
FOSFOGLICERIDI = esteri del glicerolo con 2 acidi grassi e una “testa polare” contenente
fosfato (da cui il nome fosfogliceridi) che può essere fosfo-etanolammina (in figura), fosfo-serina,
fosfo-inositolo o fosfo-colina.
Un fosfolipide di membrana, quindi, è normalmente costituito da quattro diverse molecole:
due di acido grasso, uno saturo ed uno insaturo, che costituiscono la coda non polare o idrofobica
della molecola, una di acido fosfoglicerico ed una di colina (etanolammina, inositolo o serina), che
rappresentano la porzione polare o idrofilica della molecola. Per la contemporanea presenza di una
porzione polare o idrofila e di una porzione apolare o idrofoba queste molecole vengono dette
anfipatiche.
È importante tenere ben presenti queste caratteristiche dei lipidi, perchè esse influenzano il
funzionamento di queste molecole quali costituenti delle membrane cellulari.
Gli steroidi sono lipidi formati da una complessa struttura di anelli carboniosi condensati,
essi svolgono numerose funzioni biologiche ad esempio entrano a far parte della membrana
cellulare
COLESTEROLO = struttura a 4 anelli carbociclici condensati (a ogni angolo e su ogni punta c’è
un carbonio).
Il colesterolo è un componente importante delle membrane cellulari e dal colesterolo il fegato
produce i sali biliari (importanti per la digestione).
L’unica parte polare della molecola è l’OH: nei doppi strati delle membrane l’ossidrile si orienta
verso la fase acquosa (intra- o extra-cellulare) e il resto della molecola (che è planare) si infila in
mezzo alle code degli acidi grassi. Il colesterolo impedisce l’impaccamento delle code degli acidi
grassi e assicura fluidità alle membrane.
o entrano a far parte della struttura molecolare di ormoni come estrogeni, il progesterone ed il
testosterone:
o di vitamine come la Vit.A o retinolo
Gli Zuccheri
Gli zuccheri o carboidrati sono molecole composte da carbonio idrogeno ed ossigeno e
rappresentano la principale fonte di energia per le cellule. Essi vengono classificati in base al
numero di monomeri da cui vengono formati.
I monosaccaridi sono zuccheri semplici costituiti da tre (triosi), cinque (pentosi) o sei
(esosi) atomi di carbonio. I pentosi di interesse biologico sono il ribosio che entra nella costituzione
dell'RNA ed il desossiribosio che entra nella costituzione del DNA (Fig. 15 a). Esosi di importanza
biologica sono, invece, il glucosio, molecola chiave per la produzione di energia, non che il
fruttosio, il mannosio ed il galattosio (Fig. 15b).
Classificazione dei monosaccaridi più importanti
carboidrati con
3 atomi di
carbonio
(triosi)
D-gliceraldeide diidrossiacetone
carboidrati con
5 atomi di
carbonio
(pentosi)
D-ribosio
carboidrati con
6 atomi di
carbonio
esochetosio —
—→
(esosi)
D-glucosio
D-galattosio
D-fruttosio
Tutti i monosaccaridi, escluso il diidrossiacetone, hanno uno o più atomi di carbonio
asimmetrico (centri chirali) e quindi possiedono le forme alternative di isomeri ottici
(enantiomeri).
Per n centri chirali esistono 2n enantiomeri. La gliceraldeide ha un solo centro e quindi ha 21
enantiomero, cioè 2. Il glucosio ne ha 4 per cui le forme totali di glucosio sono 24 cioè 16.
I monosaccaridi con più di tre atomi di C, in soluzione acquosa, assumono una forma ciclica
in cui il gruppo carbonilico forma un legame covalente con l'O di un gruppo ossidrilico.
Per il nostro studio è importante la ciclizzazione del glucosio che avviene tra il gruppo
aldeidico e l'O dell'OH in C5 assumendo la configurazione del pirano e quella del fruttosio che
avviene tra il gruppo carbonilico chetonico in C2 con il C5, assumendo la configurazione del
furano.
Nel passaggio dalla struttura aperta la ciclizzazione potendo avvenire in due "direzioni"
determina la formazione di un nuovo atomo di carbonio chirale, il C in 1, e di due nuovi
isomeri chiamati anomeri α e β.
Le formule in prospettiva di Haworth indicano le forme ad anello dei monosaccaridi, ma è bene
sapere che l'anello piranosico o quello piranosico non è planare ma può assumere
conformazioni diverse come quelle chiamate "a barca" o "a sedia".
conformazione a barca
conformazione a sedia
conformazione a sedia del
glucosio
glucopiranosio
I disaccaridi sono zuccheri che provengono dalla condensazione di due molecole di esosi come il
saccarosio (glucosio e fruttosio) e il lattosio ( glucosio e galattosio) (Fig. 15 c). I polisaccaridi
risultano dalla condensazione di moltissime molecole di esoso; essi di suddividono in semplici e
complessi.
I polisaccaridi semplici più importanti sono l'amido ed il glicogeno che rappresentano i depositi di
zucchero rispettivamente per le piante e per gli animali. Anche la cellulosa è un polisaccaride
semplice e rappresenta il principale elemento strutturale della parete delle cellule vegetali. Tutte e
tre queste molecole sono polimeri di glucosio (Fig. 15 d). I polisaccaridi complessi si formano
quando gli zuccheri si legano ad altre famiglie molecolari, quali lipidi e proteine, dando origine a
glicolipidi e glicoproteine, molecole che svolgono ruoli importantissimi sia nell' organizzazione
strutturale che nel controllo di molteplici funzioni cellulari.
I polisaccaridi o glicani sono formati da molte unità di monosaccaridi unite fra loro con legami glicosidici. I
polisaccaridi possono essere costituiti da un unico tipo di monosaccaride (omopolisaccaridi) o da differenti
tipi di monosaccaridi (eteropolisaccaridi). Possono essere a struttura lineare o a struttura ramificata.
Secondo la loro funzione biologica possono essere divisi in polisaccaridi di riserva o polisaccaridi funzionali.
Polisaccaridi di riserva.
Si accumulano di solito nel citoplasma cellulare sotto forma di granuli (100-400 A). La composizione di
questi granuli è piuttosto eteoìgenea in quanto oltre ai polisaccaridi questi sono composti da proteine ed
enzimi. I più importanti sono:
Amido
L’amido si trova in granuli di forma e di dimensione caratteristiche per ogni pianta. L’amido è un
polimero del (+)-glucosio ed è un polimero α-glicosidico.
Nell’amido esiste una frazione solubile in acqua che si chiama amilosio (20% del totale) che è un
polimero lineare di glucosio (60-6000 unità) legato con ponti α-glicosidici C1-C4.
L’amilopectina è un polisaccaride ramificato e forma la frazione insolubile (80%) dell’amido. Il
peso molecolare dell’amilopectina è molto più alto di quello dell’amilosio (circa 1.000.000 di unità)
ma le catena sono molto più corte (20-25 unità) e le ramificazioni sono date da ponti α C1-C6.
Glicogeno
Il glicogeno costituisce il materiale di riserva delle cellule umane (amido animale) presente
soprattutto nel fegato. La sua struttura è simile a quella dell’amilopectina ma le catene sono più
corte (12-18). Peso molecolare molto elevato.
Destrani
Sono sostanze simili alle precedenti ma costituite da legame α C1-C6, mentre le ramificazioni si
trovano in posizione 1-2, 1-3 1-4 a seconda della provenienza. Sono diffusi nei lieviti, nei batteri e
in diversi vegetali (si tratta generalemente di mannani, xilani e arabinani).
Polisaccaridi strutturali
Cellulosa
La cellulosa è il polisaccaride naturale più abbondante, essendo il componente del tessuto fibroso
delle pareti cellulari dei vegetali (costituisce circa la metà del carbonio organico presente nella
biosfera).
La cellulosa è un polimero del β-D-glucosio e a seconda delle piante da cui proviene ha un peso
molecolare compreso fra 5 X104 e 5 X 106.
Emicellulose
Sono sostanze che variano a seconda della provenienza, ma in genere sono corti polimeri ramificati
di pentosi: xilosio β 1-4 o 1-3 0 1-6 con ramificazioni di arabinosio 1-3 e 1-6.
Chitina
E’ il principale costituente degli esoscheletri degli insetti e dei crostacei ma si trova nella parete
cellulare di molti lieviti e muffe. Chimicamente è un omopolimero della N-acetil-D-glucosammina
ed ha una struttura simile alla cellulosa.
Peptidoglicano
Le pareti cellulari dei batteri hanno un carattere di rigidità dovuto alla presenza di un biopolimero ,
il peptidoglicano, formato da catene polisaccaridiche unite tra loro da catene peptidiche. Le catene
polisaccaridiche sono formate da un’unità ripetitiva disaccaridica costituita da N-acetil-Dglucosammina e acido N-acetilmurammico.