Richiami di biologia cellulare e
proprietà dei tessuti
28/09/2016
Le funzioni tessutali, di organo, di apparato e di organismo
riflettono
Le azioni coordinate di molte cellule
•quelle dotate di nucleo, che si chiamano
cellule eucariotiche e che possono essere
organismi indipendenti (per es. l’Ameba) o
parte di organismi più complessi (per es.
l’uomo);
• quelle senza nucleo, note come cellule
procariotiche, che rappresentano la forma
vivente più primitiva, della quale fanno parte
per esempio i batteri (tra cui molte forme
patogene per l’uomo).
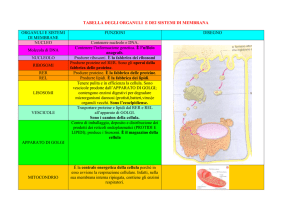
MEMBRANA PLASMATICA
È un sottile involucro che avvolge la cellula
separandola dall’ambiente esterno e che regola
attivamente lo scambio di sostanze nutritive e
di altre molecole tra la cellula e l’ambiente e
viceversa.
fornisce una barriera con permeabilità selettiva
All’interno della membrana è contenuto il CITOPLASMA:
•reticolo endoplasmatico rugoso e liscio
• ribosomi
• apparato di Golgi,
• mitocondri
• lisosomi
• centrioli
NUCLEO
Il
nucleo,
nucleare
è
circondato
costituito,
da
dall’involucro
due
membrane
separate e provvisto di pori.
Esso contiene i cromosomi, filamenti di DNA
che si evidenziano durante la divisione
cellulare.
Nel
DNA
dei
cromosomi
è
racchiusa
l’informazione genetica per la trasmissione
dei caratteri ereditari.
Il DNA può copiare se stesso, cioè può autoduplicarsi. Da una molecola di DNA se ne
ottengono due perfettamente uguali alla
prima.
La
fenomeno
duplicazione
che
precede
del
e
DNA
è
il
consente
la
duplicazione o divisione cellulare.
IL CITOSCHELETRO
Le cellule hanno uno scheletro interno detto
citoscheletro. Esso è formato da una struttura reticolare
caratteristica costituita da filamenti proteici (strutture) i
quali possono essere:
1. MICROTUBULI = i microtubuli hanno un ruolo
chiave in tutte le cellule eucariote perché creano un
sistema reticolare lungo il quale si spostano gli organuli
cellulari. Sono importanti nella mitosi (processo di
divisione cellulare) e sono elementi costitutivi di ciglia e
flagelli, fondamentali organo propulsori di molte cellule
eucariote.
2. MICROFILAMENTI = permettono alle cellule
animali di compiere movimenti di strisciamento sulle
superfici e fagocitare (circondare) particelle di varia
natura. Un esempio sono i microvilli (microfilamenti
nell’intestino).
3. FILAMENTI INTERMEDI = hanno la funzione di
rendere la cellula più resistente ai danni meccanici
provocati da urti o altri microtraumi.
CITOSCHELETRO:
Sistema di filamenti che svolge funzioni sapaziali e meccaniche nella cellula
MICROTUBULI
•
organizzazione spaziale delle
cellule
•
forma e movimento
•
riarrangiamento dei componenti
interni durante la crescita e la
divisione cellulare
•
traffico intracellulare di organelli
e molecole sostegno alla
membrana plasmatica
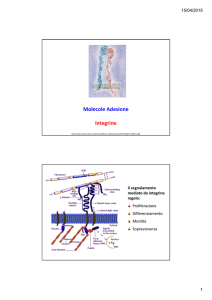
Integrine
• I recettori presenti sulla membrana
cellulare che “contattano” fibronectina,
laminina e tenascina (proteine della
CME) appartengono alla famiglia delle
integrine.
• Le integrine sono in grado di interagire
anche con proteine di membrana
(sempre interazione eterofilica)
Le integrine sono eterodimeri. Ogni eterodimero è
composto di una subunità a e una subunità b.
Entrambe le catene sono caratterizzate da un importante
dominio
extracellulare,
una
zona
idrofobica
transmembrana e da una corta coda citoplasmatica.
Nei vertebrati esistono 16
tipi di catena ed 8 tipi di
catena che generano più
di 20 integrine distinte.
Legame tra Fibronectina ed Integrina
I domini extracellulari dell’integrina legano i
componenti della ECM attraverso un legame tra
la subunità b e il motivo RGD (Arg-Gly-Asp)
l'adesione cellula-substrato è generalmente
mediata da queste piccole sequenze RGD nella
regione di legame alle cellule di proteine della
matrice extracellulare, come collagene,
fibronectina, vitronectina, laminina,
riconoscendo questa sequenza le integrine
legano le proteine della ECM, mentre grazie alla
porzione intracellulare legano i filamenti di actina
del citoscheletro via talina e vinculina
Le integrine mediano interazioni deboli
cellula-matrice e cellula-cellula
L’adesione cellula-matrice è
modulata
da
variazioni
dell’attività e del numero di
integrine
Fattori de-adesivi promuovono
la migrazione cellulare
Legame tra citoscheletro e ECM mediato da integrine
Il legame con la matrice promuove il clustering e
l’associazione con il citoscheletro.
Questo a sua volta promuove ulteriore clustering
delle integrine e organizzazione della
matrice(quindi il segnale è bidirezionale)
SINTESI PROTEICA
Le cellule, per la loro riproduzione, devono produrre proteine: sono infatti proteine gli
enzimi che consentono le numerosissime reazioni chimiche che avvengono all’interno
della cellula e sono proteine anche molte strutture della cellula.
Ogni proteina è costituita da una o più lunghe catene di aminoacidi legati in una precisa
sequenza.
Il controllo è esercitato dal DNA dei cromosomi, contenuti nel nucleo.
Il DNA contiene infatti le informazioni in codice per la sintesi di tutte le proteine
della cellula. In pratica, nel nucleo abbiamo il “manuale di istruzioni” in codice, per la
produzione di tutte le proteine della cellula.
Sebbene la sintesi proteica sia un processo “continuo” può essere,
idealmente, suddiviso in tre distinte fasi:
INIZIO, CRESCITA o ALLUNGAMENTO, TERMINAZIONE
1. Sintesi dell’RNA messaggero
(mRNA), come copia di una
porzione di DNA (un gene), ad
opera dell’enzima RNA
polimerasi: avviene nel nucleo.
Le proteine vengono nel
citoplasma, da organuli speciali
chiamati ribosomi
I ribosomi sono in grado di
leggere il messaggio trasmesso,
sono cioè in grado di capire
(decodificare) il codice genetico.
2. L’RNA messaggero attraversa i pori della
membrana nucleare per raggiungere, nel
citoplasma, i ribosomi, organuli della sintesi
proteica.
Riassumendo:
1) Un mRNA viene trascritto dal DNA;
2) l’mRNA si porta dal nucleo ai ribosomi;
3) ai ribosomi giunge l’RNA messaggero,
con la sua sequenza di nucleotidi, che
formano precise triplette o codoni;
4) giungono gli RNA di trasporto con gli
anticodoni, corrispondenti alle triplette
dell’RNA messaggero e si allineano uno
di seguito all’altro;
5) all’altra estremità degli RNA di trasporto
si vengono a trovare allineati,
nell’esatto ordine, gli aminoacidi
corrispondenti, che vengono poi legati
tra loro nella sequenza precisa per
costituire la proteina “X” richiesta dalla
cellula
utilizzo
VITALITA’ CELLULARE
Altri metodi:
- calceina-AM + etidio bromuro
-Fluoresceina diacetato + bromuro d’etidio
Si usa moltissimo in ingegneria dei
tessuti per valutare, per esempio,
la citotossicità di un materiale
-Alamar blu (attività metabolica)
Marker e separazione magnetica
Marker e separazione magnetica
Visualizzazione cellule, marcatura in fluorescenza
Vale per tutte le cellule, non solo le staminali…
Funzione meccanica delle fibre della matrice
Fibre collagene: conferiscono sostegno (cartilagine, ossi,
legamenti)
Fibre elastiche: conferiscono elasticità (pareti dei grossi vasi
sanguigni).
Fibre reticolari: costituiscono reticolati all’interno di organi pieni
(fegato).
solitamente legate alle proteine per formare i proteoglicani, che possono
essere ricchi di gruppi solfato (il condroitinsolfato, il dermatansolfato,
l'eparansolfato e il cheratansolfato) o privi (l'acido ialuronico)
due gruppi: uno con funzione principalmente strutturale (i collageni e
l'elastina), e uno con funzioni principalmente adesive (la fibronectina, le
laminine, le entactine o nidogeni e la vitronectina). Tutte queste
macromolecole sono caratterizzate da grande varietà di forme e di
dimensioni
Le molecole dei glicosamminoglicani e dei proteoglicani formano, nei tessuti
connettivi, una sostanza 'fondamentale' gelatinosa e fortemente idratata, in cui sono
immerse le proteine fibrose; questo gel di polisaccaridi consente la diffusione di
sostanze nutritive, metaboliti e ormoni tra il sangue e le cellule dei tessuti e resiste
alle forze compressive esercitate sull'ECM
FIBRE ELASTICHE
•Conferiscono il recupero elastico dei tessuti:
possono estendersi fino al 150% della loro lunghezza
originaria e poi tornare alla loro lunghezza originaria
•Sono molto più sottili dei fasci di fibre di collagene,
RAMIFICATE e molto resistenti ad agenti chimici e
fisici; nel connettivo si riuniscono a formare una rete
a maglie lasse;
•Sono costituite da –una componente PROTEICA ,
ELASTINA,che conferisce la elasticità ed una
componente GLICOPROTEICA, FIBRILLINA, che
conferisce stabilità.
Alcuni collageni, interagendo con le laminine, le entactine o nidogeni e i
proteoglicani, costituiscono le impalcature delle lamine basali.
Le proteine d'adesione agevolano inoltre la connessione delle cellule tessutali
all'ECM stessa e ne influenzano la polarizzazione: la fibronectina, infatti,
favorisce l’interazione e adesione dei fibroblasti e di altre cellule con la
matrice dei tessuti connettivi, mentre le laminine favoriscono quello delle
cellule epiteliali con le lamine basali.
La vitronectina, infine, interagisce con l'elastina, i glicosamminoglicani e i
collageni e modula l'angiogenesi e la degradazione dell'ECM stessa.
Oltre ai GAG e alle proteine fibrose strutturali e adesive sopra descritte,
nell'ECM sono presenti anche altre proteine, definite 'proteine matricellulari'.
Esse costituiscono una classe di proteine di secrezione che non hanno
funzioni strutturali, ma rivestono il ruolo di adattatori molecolari in quanto
interagiscono con le proteine fibrose e adesive dell'ECM, con recettori
cellulari o con altre molecole come fattori di crescita, citochine e proteasi.
Sebbene diverse tra loro per funzione, le proteine matricellulari hanno in
comune la capacità di modulare le interazioni cellula-matrice
I principali recettori delle macromolecole
dell'ECM: le INTEGRINE
Gran parte delle macromolecole
dell'ECM può stabilire interazioni altamente
funzionali con le cellule di un tessuto
grazie ai recettori che queste esprimono. Tra
questi, i più importanti sono le integrine, in
quanto rappresentano la via fondamentale con
cui le cellule si legano all'ECM e rispondono ai
suoi stimoli.
Le integrine sono proteine eterodimeriche transmembranarie costituite da due
subunità associate, indicate con le lettere greche α e β, che presentano entrambe
un'estesa porzione extracellulare, un dominio transmembrana e una porzione
intracitoplasmatica .
Al momento sono state descritte più di 10 differenti catene β (β1, β2, β3, β4, ecc.) e
più di 20 diverse subunità α (α1, α2, α3, α4, α5, α6, α7, ecc., con l'eccezione di due,
definite αv e αE). Mentre le catene β hanno la possibilità di unirsi a numerose
catene α, queste ultime si associano generalmente a una sola subunità β, a
differenza della subunità αv che ha la capacità di legare diverse catene β come le
β1, le β3, le α5, le α6, e le α8.
Le integrine fungono da collegamenti transmembrana, mediando le interazioni fra il
citoscheletro e la matrice extracellulare richiesti per permettere alle cellule di aderire
alla matrice. La maggior parte delle integrine è collegata ai filamenti di actina.
Nelle condizioni giuste, questo collegamento porta all’aggregazione delle integrine e
alla formazione di adesioni focali tra la cellula e la matrice extracellulare.
I complessi delle adesioni focali, identificati come strutture
specializzate di adesione, sono composti da:
• Integrine
• Proteina chinasi, quali le «focal adhesion kinase», FAK, e il «non‐
receptor tyrosine kinase receptor Src
• Proteine adattatrici (es. Shc) Intermediari di segnalamento (es.
famiglia Rho delle GTPasi)
• Proteine citoscheletriche che legano l’actina (es. talina,
la α‐actinina, la paxillina, la tensina e la vinculina)
• Altre proteine di segnalamento.
Il signaling mediato dalle integrine regola l’espressione genica, la
crescita cellulare, il differenziamento e la sopravvivenza.
La fibronectina : proteine fibrose adesive
La fibronectina (FN) è la prima delle proteine adesive dell'ECM a essere stata
caratterizzata. Queste proteine hanno il compito sia di organizzare l'ECM sia di
permettere alle cellule di connettersi con essa. La fibronectina si lega a diversi
altri componenti dell'ECM, tra cui i collageni, in modo particolare al collagene di
tipo III, e ai proteoglicani tramite domini specifici, e alle cellule tramite i recettori
integrinici.
La fibronectina è una grossa glicoproteina composta
da due subunità, unite da due legami disolfuro
all'estremità carbossiterminale della molecola.
Ciascuna subunità è ripiegata in una serie di domini
funzionali, che consistono di moduli ripetuti.
Coinvolta direttamente nell'adesione, nello spreading (il
processo col quale la cellula si appiattisce sul
substrato, aderendovi attivamente), nella migrazione,
nella proliferazione, nella differenziazione e nella
modulazione dell'apoptosi delle cellule dei tessuti