12/05/15
Negli animali l’ossidazione ad
acetil-CoA degli acidi grassi a
catena lunga è una delle vie
essenziali per produrre energia
GRASSI DELLA DIETA
40 %
LIPIDI DI DEPOSITO
Un mammifero può contenere
riserve pari al 5-25% del suo
peso corporeo
LIPIDI SINTETIZZATI IN
UN ORGANO
ed esportati in un altro
1
12/05/15
I GRASSI intracellulari contengono sei volte più ENERGIA
METABOLICA POTENZIALE, in rapporto alla massa, del
GLICOGENO intracellulare
1. La maggior parte degli atomi di carbonio negli acidi
grassi sono in forma altamente ridotta
2. Mentre il GLICOGENO lega 2gr di H2O / gr di
carboidrato, i GRASSI, essendo estremamente apolari,
sono ANIDRI
Circa il 40% del contributo calorico all’alimentazione
occidentale proviene dai grassi. Inoltre i carboidrati ingeriti in
eccesso, rispetto alla possibilità di essere degradati o
immagazzinati come glicogeno, vengono prontamente
convertiti in grassi. Il CERVELLO l’unico tessuto che non è in
grado di utilizzare i grassi come fonte energetica può,
quando il livello di glucosio ematico diminuisce, adattarsi ad
utilizzare i CORPI CHETONICI
ALIMENTAZIONE
BIOSINTESI
EX-NOVO
MOBILIZZAZIONE
RISERVE
adipociti
epatociti
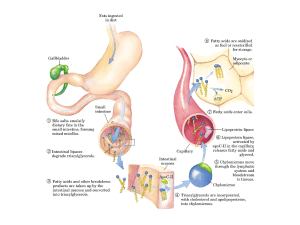
SALI BILIARI : azione detergente, emulsionano
i lipidi e producono micelle che consentono
l’attacco da parte degli enzimi digestivi,
solubili in acqua, e facilitano l’assorbimento
del lipide attraverso la mucosa intestinale
ASSORBIMENTO
LIPASI PANCREATICA
2
12/05/15
a. grassi
ossidazione
Catena
respiratoria
IN TUTTE LE CELLULE
MENO ENCEFALO E
GLOBULI ROSSI
Acetil-CoA
Ciclo di Krebs
3
12/05/15
TRIACILGLICEROLI
5% GLICEROLO
95% A.GRASSI
La maggior parte del
glicerolo è rilasciato nel
torrente circolatorio
viene incorporato dalle
cellule del fegato per la
sintesi di glucosio
Gli acidi grassi entrano nel
citosol dal sangue ma non
p o s s o n o p a s s a r e
direttamente le membrane
mitocondriali devono prima
subire la sequenza di tre
reazioni catalizzate
GLICOLISI o GLUCONEOGENESI
enzimaticamente
4
12/05/15
GLI ACIDI GRASSI SONO ATTIVATI E TRASPORTATI NEI MITOCONDRI
Acido grasso + CoA + ATP
acil-CoA + AMP + PPi
ΔG°’= -31 KJ/mole
Presente nella
membrana
mitocondriale
esterna
intermedio
Reazione
all’equilibrio che
viene spostata
verso la
formazione di
acil-CoA
dall’idrolisi del
pirofosfato
Diversi
isoenzimi
dell’acil-CoA
sintetasi
agiscono su a.
grassi a
diversa
lunghezza
INGRESSO DEGLI A. GRASSI NEI MITOCONDRI ATTRAVERSO IL
TRASPORTATORE ACIL-CARNITINA/CARNITINA
L’aciltransferasi I viene
inibita dal malonil-CoA(il
primo intermedio della
sintesi degli acidi
grassi)
Impedendo l’entrata
del gruppo acilico nel
mitocondrio e di conseguenza
l’ossidazione degli acidi grassi
5
12/05/15
Gli a.grassi, nella
matrice mitocondriale,
vanno incontro al
distacco ossidativo di
unità bicarboniose nella
fo r m a d i a c e t i l - C o A,
iniziando dall’estremità
carbossilica della
catena idrocarburica
6
12/05/15
Ossidazione
analoga a quella
della succinato
deidrogenasi.
Entrambe legate
alla membr ana
interna del
mitocondrio
Tre forme
isoenzimatiche
Specifico per lo
steroisomero L
Le quattro reazioni
sono ripetute per
formare acetil-CoA
e ATP
7 cicli
8 acetil-CoA
7
12/05/15
Palmitoil-CoA + 7 CoA + 7 FAD + 7 NAD+ + 7 H2O
8 acetil-CoA + 7 FADH2 + 7 NADH + 7 H+
3 NADH
1 FADH2
1 GTP
X8
24 NADH
8 FADH2
8 GTP
Palmitoil-CoA + 23O2 + 100 Pi + 100 ADP
31 NADH
2,5 ATP x 31
15 FADH2
1,5 ATP x 15
8 GTP
100 ATP + 46 H2O
46 – 7 = 39 molecole di H2O
Negli animali in letargo, l’ossidazione degli acidi
grassi fornisce l’energia metabolica, calore ed
acqua, necessari per un animale che non mangia e
non beve per un periodo molto lungo
La demolizione dei
triacilgliceroli
produce glicerolo,
che viene convertito
i n g l u c o s i o
m e d i a n t e l a
gluconeogenesi.
L’orso riassorbe al
livello renale e
ricicla anche l’urea
prodotta dalla
degradazione degli
aminoacidi
Il grizzly usa il grasso corporeo come unica fonte di energia nel periodo di
letargo, mantenendo una temperatura corporea di 32-35°C.
8
12/05/15
ossidazione degli acidi grassi nelle cellule è
controllata dalla disponibilità di substrato
L’azione del glucagone e
dell’adrenalina causa la demolizione e
il rilascio dei grassi dagli adipociti che
si traduce in un accumulo di acidi
grassi in altre cellule
regolazione
or monale della
mobilizzazione
del grasso negli
adipociti
Livello del malonil-CoA
9
12/05/15
L’acetil-CoA metabolita centrale dell’ossidazione sia
del glucosio che degli acidi grassi viene a sua volta
convertito nel citosol in acidi grassi. Costituisce un
meccanismo efficiente per la conversione degli
zuccheri in riserve lipidiche.
Come le altre vie biosintetiche le sequenze di reazione,
per la sintesi degli acidi grassi, sono endoergoniche e
riduttive, utilizzano ATP come fonte di energia
metabolica e necessitano di equivalenti riducenti
sottoforma di NADPH
BIOSINTESI
Precursore biosintetico
acidi grassi
fegato
a. grassi
ossidazione
Catena
respiratoria
Acetil-CoA
Corpi chetonici
Ciclo di Krebs
10
12/05/15
La for mazione di
malonil-CoA, a partire
da acetil-CoA e HCO3è la tappa iniziale
della biosintesi
ACETIL-CoA CARBOSSILASI
Gli acidi grassi vengono sintetizzati mediante una
sequenza di quattro reazioni ripetute che sono
uguali a ritroso (riduzioni) a quelle che
caratterizzano la β-ossidazione
Tutte le reazioni sono sintetizzate da un
complesso multienzimatico, chiamato ACIDO
GRASSO SINTASI Costituito da almeno sette
proteine differenti
β-OSSIDAZIONE
BIOSINTESI
trasportatori di e-
NAD+; FAD
NADPH
Gruppo (-SH)
attivante
coenzima A
gruppi –SH
legati all’enzima
11
12/05/15
AT
acetil-CoA-ACPtransacetilasi
KS
β-chetoacil-ACP
sintasi
MT
malonil CoA-ACP
transacetilasi
KR
β-chetoacil-ACP
reduttasi
HD
β-idrossiacil-ACPdeidratasi
ER
enoil-ACP reduttasi
Le proteine cooperano tra loro per formare acidi grassi a 16
atomi di carbonio a partire da Acetil-CoA e malonil-CoA
12
12/05/15
Gli intermedi che si formano durante il processo
restano legati covalentemente ad uno dei due gruppi
tiolici del complesso
Un punto di legame è l’SH di un residuo di cys in una
delle 7 proteine (la β-chetoacil-ACP sintasi)
L’altro è l’SH della proteina trasportatrice degli acili
(ACP)
13
12/05/15
4’-fosfo-pantoteina gruppo
prostetico dell’ACP (proteina che
trasporta acili)
Intermedio della sintesi del CoA
Ha la funzione di braccio flessibile
che ancora le catene dell’acido
grasso in fase di crescita sulla
superficie del complesso
dell’acido grasso sintasi e
trasporta gli intermedi delle
reazioni da un sito attivo all’altro
L’acido grasso sintasi
utilizza gruppi acetilici
e malonilici
Acetil-CoA-ACP-transacetilasi
(AT)
Trasferisce i gruppi acilici dal
CoA al gruppo SH di un residuo
di cys della β-chetoacil-ACP
sintasi (KS)
14
12/05/15
Trasferimento del gruppo
malonile dal CoA al gruppo
SH dell’ACP
Malonil-CoA-ACP
transferasi (MT)
Prima tappa
CONDENSAZIONE
β-chetoacil-ACP sintasi (KS)
L’atomo di carbonio della CO2
che si forma in questa reazione
è lo stesso introdotto nella
molecola di malonil-CoA
durante la reazione dell’acetilCoA con il bicarbonato
catalizzata dall’acetil-CoA
carbossilasi
15
12/05/15
Seconda tappa: riduzione del
gruppo carbonilico
β-chetoacil-ACP reduttasi
terza tappa deidratazione
β-idrossiacil-ACP deidratasi
Quarta tappa riduzione del
doppio legame
Enoil-ACP reduttasi
16
12/05/15
Il gruppo butirrilico viene trasferito dal
gruppo –SH della fosfopantoteina al
residuo cys della β-chetoacil-ACP sintasi,
che inizialmente era occupato dal gruppo
acilico, per iniziare un nuovo ciclo delle
quattro reazioni.
REAZIONE COMPLESSIVA DELLA SINTESI DEL PALMITATO
Prima parte
7 Acetil-CoA + 7 CO2 + 7 ATP
7 malonil-CoA + 7 ADP + 7 Pi
Seconda parte
Acetil- CoA + 7 malonil-CoA + 14 NADPH +14H+
palmitato + 7 CO2 + 8 CoA +14 NADP++ 6 H2O
REAZIONE COMPLESSIVA
8 Acetil-CoA + 7 ATP + 14 NADPH +14H+
palmitato + 8 CoA +14 NADP++ 6 H2O + 7 ADP + 7 Pi
17
12/05/15
Deriva dalla via del
fosfogluconato a partire da
glucosio-6-fosfato
Deriva dal catabolismo
dei carboidrati
Deriva dal piruvato e si
forma nei mitocondri
Intermedio chiave tra il
metabolismo lipidico e il
metabolismo dei carboidrati
Il citrato serve come
vettore per il trasporto
delle unità acetiliche dal
mitocondrio al
citoplasma per la sintesi
degli acidi grassi
FONTI DI NADPH PER LA SINTESI DEGLI A.GRASSI
Enzima malico
18
12/05/15
Acetil-CoA sintetizzato nei mitocondri viene
trasportato nel citosol sottoforma di citrato
19
12/05/15
Quando nel
mitocondrio
aumenta la
quantità di acetilCoA e ATP, il citrato
viene esportato
fuori determinando
attivazione
dell’acetil CoA
carbossilasi e nello
stesso tempo
determina
l’inibizione della
fosfofruttochinasi-1
BIOSINTESI DEGLI A. GRASSI
INGRESSO DEGLI A. GRASSI NEI MITOCONDRI ATTRAVERSO IL
TRASPORTATORE ACIL-CARNITINA/CARNITINA
CoA-SH con funzioni diverse nei due compartimenti
Trasportatore
acil-carnitina/carnitina
L’ACILTRANSFERASI I viene inibita dal malonil-CoA, il primo
intermedio della sintesi degli acidi grassi. Questo meccanismo di
controllo è un esempio dei vantaggi che si possono avere dal
separare le vie biosintetiche da quelle cataboliche in compartimenti
cellulari diversi.
20
12/05/15
L’ossidazione avviene nel
fegato quando l’ossidazione
dei grassi è molto elevata
prodotto in
piccole quantità
è eliminato con
la respirazione
Nel fegato l’acetil-CoA
può essere trasformato
in corpi chetonici, per
esser e espor tato ad
altri tessuti
ossidati
nel ciclo
dell’acid
o citrico
21
12/05/15
La produzione e l’esportazione dei
corpi chetonici dal fegato ai tessuti
extraepatici consente di continuare
l’ossidazione degli acidi grassi nel
fegato anche quando l’acetil-CoA
non viene ossidato dal ciclo
dell’acido citrico
1. Solubili in H2O
2. si formano quando la quantità di AcetilCoA eccede la capacità ossidativa del
fegato
3. sono utilizzati dai tessuti extraepatici in
proporzione alla concentrazione ematica
Rapporto acetoacetato / idrossibutirrato
dipende dal rapporto NADH/NAD+
all’interno dei mitocondri
Può formarsi
direttamente dalla βossidazione degli
acidi grassi
Affinchè possa avvenire
la chetogenesi entrambi
gli enzimi della via
devono essere presenti
nel mitocondrio
22
12/05/15
I corpi chetonici sono
un carburante per i
tessuti extraepatici
In un animale ben nutrito la
concentrazione di corpi
chetonici nel sangue non
supera la concentrazione di
0,2 mM
Inbisce la piruvato deidrogenasi
Attiva la piruvato carbossilasi
GLUCONEOGENESI
23
12/05/15
La produzione e l’esportazione di
corpi chetonici liberano CoA e
consentono di continuare ad ossidare
gli acidi grassi
L’acetil-CoA formato
dall’ossidazione degli
acidi grassi entra nel
ciclo di Krebs solo se la
degradazione dei grassi
e dei carboidrati sono
adeguatamente
bilanciate
IL DIGIUNO PROLUNGATO O IL DIABETE MELLITO
NON CONTROLLATO
L’ossalacetato viene
adoperato per la
gluconeogenesi e
diventa insufficiente
per legare l’acetil-CoA
prodotto in grandi
quantità dal
catabolismo degli a.
grassi, nel ciclo di
Krebs.
Portano alla sovrapproduzione di corpi
chetonici
24