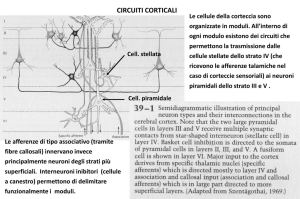
IL SOGNO NELLA SCIENZA
di Luisa Ghianda
Il sonno viene come l’avanzare della marea. Opporsi è impossibile. È un sonno così
profondo che né lo squillo del telefono né il rumore delle auto che passano fuori mi
arrivano all’orecchio. Nessun dolore, nessuna tristezza laggiù: solo il mondo del sonno
dove precipito con un tonfo.
Banana Yoshimoto, Sonno profondo
Il sonno, imperativo biologico necessario per sostenere la vita, è uno stato di riposo
contrapposto alla veglia. La distinzione principale tra il sonno e la veglia consiste in una
barriera percettiva eretta tra mondo cosciente e mondo esterno, nel primo stato, sebbene
uno stimolo sensoriale di un certo livello, ad esempio un rumore forte, può superare questa
barriera e svegliare il dormiente. Sospensione dello stato di coscienza, durante la quale
l'organismo recupera energia, interrompendo, completamente o parzialmente, la volontà,
rallentando
le
funzioni
neurovegetative,
arrestando
in
modo
parziale
i rapporti
sensomotori con l'ambiente, il sonno è una operazione indispensabile per il ristoro
dell'organismo.
Nonostante
l’apparente
stato
di
quiete,
dormire
è
un processo
fisiologico attivo, che coinvolge l'interazione di componenti multiple del sistema nervoso
centrale ed autonomo. A livello cerebrale avvengono complessi cambiamenti: alcune cellule
encefaliche, in alcune fasi del sonno, hanno una attività 5-10 volte maggiore rispetto a quella
che hanno in veglia1.
Le tre misure principali usate per definire la fisiologia del sonno sono:
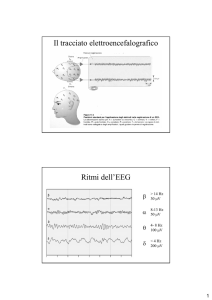
- l'elettroencefalogramma (EEG), che traduce l’attività cerebrale in onde elettriche;
- l'elettrooculogramma (EOG), che registra i movimenti oculari e li traduce in onde elettriche;
- l'elettromiogramma (EMG), che registra l'attività muscolare.
La misurazione può includere altri parametri come la pressione arteriosa, la ph-metria
esofagea, la temperatura corporea.
Nel 1953 Aserinsky e Kleitman scoprirono la presenza dei movimenti oculari
rapidi (REM) durante il sonno2. Questa osservazione permise di differenziare il sonno in
una fase REM e in una fase non-REM (nREM), fasi che si alternano ciclicamente, dando
vita a notevoli variazioni elettroencefalografiche. I due studiosi scoprirono che, svegliando
1
V. Sbrescia, “Il sogno e i dal punto di vista scientifico. Psico-Pratika” [Online]. N° 79. Disponibile all’indirizzo:
www.humantrainer.com (Consultato in data 28/06/2015).
2
E. Aserinsky; N. Kleitman, “Regularly occurring periods of eye motility, and concomitant phenomena, during sleep”.
Science, New Series, n. 3062, vol. 118, 1953, pp. 273-274.
soggetti dormienti nel momento in cui avvenivano questi caratteristici movimenti rapidi degli
occhi, essi riferivano che stavano sognando.
Nel
1968 Rechtschaffen e Kales, basandosi
sull'analisi
dei
parametri
elettroencefalografici, elettromiografici ed elettrooculografici classificarono il sonno in 5
stadi: 4 stadi non-REM (stadio 1; stadio 2; stadio 3; stadio 4) ed uno stadio REM.
Il sonno presenta un'alternanza regolare di fasi non-REM, sonno lento e sincronizzato, detto
sonno ortodosso, e REM, sonno rapido, desincronizzato, detto sonno paradosso. Dopo
l'addormentamento il soggetto passa progressivamente dallo stadio 1 del sonno non-REM
allo stadio 4, per poi ritornare allo stadio 3 e allo stadio 2 e quindi passare alla prima fase di
sonno REM. Ciò significa che, tra i 70 e i 90 minuti dopo l'addormentamento, si verifica la
prima fase di sonno REM, della durata di 15 minuti circa, al termine del quale il primo ciclo
di sonno si conclude. Ne susseguono poi altri di durata piuttosto costante, dove il sonno
REM tende, però, ad aumentare in durata, a scapito del sonno non-REM, ed in particolare
degli stadi 3 e 4 (sonno profondo), che si fanno più brevi. Durante la notte, il sonno REM
costituisce circa il 25% della durata totale del sonno. È possibile che tra i vari cicli vi siano
momenti di veglia.
Durante la veglia, l'EEG alterna fondamentalmente due pattern:
1- pattern di attivazione, caratterizzato da onde di basso voltaggio (10-30 microvolt) ed
alta frequenza (16-25 Hz), presente quando la persona è in stato di attenzione ad
occhi aperti; i movimenti oculari sono sia rapidi, sia lenti e il tono muscolare è medioalto;
2- pattern di attività alfa, caratterizzato da onde sinusoidali di 8-12 Hz, attività
tipicamente presente ed abbondante quando il soggetto è rilassato ad occhi chiusi.
Durante lo stadio 1, fase di addormentamento, l'attività alfa diminuisce, il pattern di
attivazione è scarso, l'EEG è costituito principalmente da onde di basso voltaggio di
frequenza mista tra i 3-7 Hz, i movimenti degli occhi sono ancora presenti ma lenti, rotanti
e oscillatori, l'EMG mostra una attività tonica persistente, benché di intensità inferiore
rispetto alla veglia. Trattasi di uno stato crepuscolare, fra veglia e sonno leggero.
Nello stadio 2, fase di sonno leggero, è presente una attività di fondo di voltaggio
relativamente basso, con frequenza variabile ma vicina alle onde theta (3-7 Hz). Tale stadio
è caratterizzato dalla presenza di due componenti: i complessi K3 e i fusi del
sonno (o spindles)4; i movimenti degli occhi sono lenti, l'EMG si riduce ulteriormente, la
coscienza è sopita, i muscoli si rilassano lentamente.
Nello stadio 3, fase di sonno profondo, l’EEG mostra onde delta di grande ampiezza
(>75 microvolt) e bassa frequenza (circa 0,5 - 4 Hz); il tono muscolare è lievemente ridotto
ed i movimenti degli occhi praticamente assenti; i fusi del sonno possono presentarsi oppure
no, mentre sono presenti i complessi K, sebbene spesso siano difficilmente distinguibili dalle
onde delta. In questa fase il sonno comincia a diventare profondo.
Lo stadio 4, fase di sonno profondo effettivo, è caratterizzato dalla presenza di onde
delta, che qui raggiungono la massima ampiezza e la minima frequenza; i fusi possono
essere assenti o presenti, mentre i complessi K sono presenti, ma pressoché irriconoscibili
dal ritmo delta di fondo; i movimenti degli occhi non sono presenti, mentre persiste uno stato
di attivazione muscolare tonica molto basso. In questa fase l'attività metabolica del cervello
è ridotta, cosicché vi è un minor consumo di ossigeno e glucosio. Con esclusione dello
stadio 1, transitorio, gli stadi del sonno più profondi (3 e 4) sono caratterizzati da una
3
Forme d'onda di alto voltaggio rilevabili in un elettroencefalogramma (EEG). Sono stati definiti il "più grande evento
nell'elettroencefalogramma di un uomo sano". Questi complessi sono più frequenti nei primi cicli di sonno. Si ritiene che
i complessi K assolvano ad almeno due funzioni: in primo luogo, sembra determinano la soppressione dell'eccitazione
corticale in risposta a stimoli che il cervello addormentato non ritiene debbano essere valutati come pericolosi, e la
seconda, favoriscono il consolidamento della memoria basata sul sonno. Il complesso K è stato scoperto nell'anno 1937
nei laboratori privati di Alfred Lee Loomis.
4
Treni di onde con frequenza di 12-16 Hz e della durata di 0,5-1,5 secondi, che inibiscono l'elaborazione di informazioni
non necessarie, evitando così l'interruzione dello stadio 2.
maggiore difficoltà di risveglio, tanto che se ci si sveglia nella fase 4 si può
rimanere particolarmente confusi per qualche minuto.
Lo stadio REM è caratterizzato da un EEG a basso voltaggio con frequenze miste,
che ricorda molto quello dello stadio 1, se non per le scariche di onde con la morfologia a
“dente di sega”, tipiche di questa fase. Compaiono le onde PGO (ponto-genicolo-occipitali);
l'attività dell'ippocampo si fa sincronizzata con la comparsa di onde theta. Lo stadio prende
il nome dai movimenti oculari rapidi (rapid eye movements). Caratteristica dello stadio REM
è la paralisi dei muscoli, per evitare di mimare i sogni. Il cervello consuma ossigeno e
glucosio come se il soggetto fosse sveglio e stesse svolgendo un'attività intellettuale. Se ci
si sveglia in questa fase si è perfettamente orientati. Vi è un controllo più impreciso delle
funzioni vegetative dell'organismo: la pressione arteriosa aumenta e subisce sbalzi; la
frequenza cardiaca aumenta e possono comparire extrasistoli; la frequenza respiratoria
aumenta, facendosi più irregolare; la termoregolazione è in parte compromessa. Si possono
verificare erezione del pene nell'uomo e modificazioni genitali nella donna. Il sonno REM
raggiunge un picco all'età di 1 anno, tende a ridursi con l'avanzare dell'età e diminuire in
favore del sonno non-REM. Il termine “sonno paradosso” deriva dal fatto che l'elevata
attività celebrale e i rapidi movimenti oculari presenti sono in contrasto con il grado di
generale rilassamento muscolare.
Nel corso della notte diminuiscono progressivamente le fasi di sonno profondo e aumentano
di durata e di intensità le fasi REM. Un giovane adulto arriva al sonno REM più o meno 90
minuti dopo l'addormentamento; questa fase, che si ripete all'incirca ogni 2 ore, dura sempre
un po' di più fino ad arrivare al momento più lungo che precede il risveglio. Se una persona
è disturbata in fase REM o nel momento di sonno profondo, facilmente presenta sintomi di
stress e di nervosismo.
Durante il sonno sono attivi il tegmento pontino, la circonvoluzione paraippocampale,
l'amigdala, l'ippocampo, la corteccia del cingolo anteriore, aree corticali temporo-occipitali,
aree limbiche, alcuni nuclei del talamo e parte del prosecefalo basale. Sono invece deattivati
la corteccia prefrontale dorsolaterale, la corteccia del cingolo posteriore e la corteccia
parietale.
Gli aspetti emozionali dei sogni sembrano avere origine nelle aree limbiche e paralimbiche,
nell'amigdala e nella corteccia anteriore del cingolo, mentre la deattivazione della corteccia
prefrontale comporta probabilmente la difficoltà nel ricordare i sogni nello stato di veglia.
Un primo sistema che controlla e mantiene lo stato di veglia è rappresentato dai
nuclei aminergici del tronco encefalico, in particolare dai neuroni noradrenergici del locus
coeruleus e dai neuroni serotoninergici dei nuclei del rafe; si presume che abbiano un ruolo
anche i neuroni dopaminergici della sostanza nera. Questi neuroni proiettano diffusamente
alla corteccia, al talamo, all'ipotalamo e all'ippocampo. Quando il soggetto è vigile, la
frequenza di scarica dei neuroni di questi sistemi è massima, mentre si riduce notevolmente
durante il sonno non-REM e quasi del tutto durante il sonno REM. Questi neuroni possono
anche andare incontro a fenomeni di auto-inibizione, che favoriscono il sonno. Condizioni
che ne stimolano l'attività promuovono, invece, la veglia.
Se la stimolazione del sistema noradrenergico sembra stimoli e mantenga la veglia,
la serotonina, pur stimolando anch'essa la veglia, favorisce, nel tempo, il rilascio di sostanze
che promuovono il sonno ed inibisce i neuroni colinergici del prosencefalo basale, coinvolti
nel mantenimento della veglia, svolgendo quindi un ruolo ambiguo.
Un
secondo
sistema
che
promuove
neuroni colinergici del prosencefalo basale.
Questi
la
veglia
neuroni
è
proiettano
costituito
dai
alla corteccia,
attivandola, all'ippocampo e all'amigdala e, oltre che durante la veglia, sono attivi durante la
fase REM, mentre sono poco attivi in quella non-REM. Sono inibiti da terminazioni
serotoninergiche provenienti dai nuclei del rafe. I nuclei colinergici del tronco encefalico
contengono una prima popolazione di neuroni attivi durante il sonno REM, che scaricano a
bassissima frequenza durante la veglia e il sonno non-REM, ed una seconda popolazione
costituita da neuroni la cui frequenza di scarica è massima durante la veglia e durante il
sonno REM e che proiettano al talamo e all'ipotalamo, attivandoli.
Il nucleo tubero-mammillare contiene i neuroni istaminergici ipotalamici che proiettano
diffusamente a quasi tutto il sistema nervoso centrale, promuovendo il mantenimento della
veglia. L'inibizione di questi neuroni con antistaminici induce sonnolenza.
L'ipotalamo postero-laterale comprende un piccolo gruppo di neuroni orexinergici, che
proiettano diffusamente alle strutture coinvolte nella regolazione del ciclo sonno-veglia e
sono coinvolti anche nella regolazione dell'assunzione di cibo.
Il nucleo preottico ventrolaterale dell'ipotalamo anteriore, altre aree ipotalamiche e del
prosencefalo basale contengono neuroni GABAergici e neuroni rilascianti galanina, che
proiettano alle strutture coinvolte nel mantenimento della veglia e la inibiscono, favorendo il
sonno.
Il rilascio di adenosina da parte del metabolismo cerebrale si accompagna ai periodi di
veglia. Questa sostanza, interagendo con i suoi recettori, inibisce i circuiti che promuovono
la veglia ed attiva quelli che promuovono il sonno, principalmente disinibendo i neuroni
GABAergici del nucleo preottico ventrolaterale dell'ipotalamo anteriore. La caffeina e
stimolanti correlati, invece, contrastano l'effetto dell'adenosina, perché le impediscono il
legame ai suoi recettori. Le citochine possono promuovere il sonno in condizioni fisiologiche
o patologiche.
Diverse sono le teorie circa la funzione del sonno nel nostro organismo.
Secondo la “teoria del recupero”, il sonno avrebbe il compito di ristorare l'organismo
durante le fasi non-REM, mentre durante la fase REM avrebbe una mansione di recupero,
svolgendo un ruolo di riprogrammazione genetica dei comportamenti innati, e di fissazione
della memoria, facilitando l'incorporazione di nuovi comportamenti appresi in veglia.
Solitamente, si consiglia di dormire almeno 8 o 9 ore. Se un individuo è sottoposto a più di
queste ore di base sarà pervaso da uno stato di tranquillità e di stanchezza, poiché il nostro
organismo tende a conservare lo stato di massima rilassatezza.
La National Sleep Foundation degli Stati Uniti, in collaborazione con l'American Geriatrics
Society e con l'American Academy of Pediatric ha approfondito il numero di ore congrue di
sonno secondo l’età dell’individuo5. Per ogni fascia di età non si dovrebbe superare il limite
massimo stabilito, sebbene non si tratti di regole ferree ma solo di consigli:
- Da 0 a 3 mesi: la durata ideale del sonno va dalle 14 alle 17 ore.
- Da 4 a 11 mesi: le ore di sonno raccomandate sono 12-15, ma il range è più ampio, 1018.
- Da 1 a 2 anni: 11-14 ore al giorno, ma il range da mantenere è entro le 9-16 ore.
- Da 3 a 5 anni: da 10 a 13 ore, non più di 14 ore e non meno di 8.
- Da 6 a 13 anni: da 9 a 11 ore, non meno di 7 e non più di 12.
- Da 14 a 17 anni: da 8 a 10 ore, non più di 11 e non meno di 7.
- Dai 18 ai 25 anni: da 7 a 9 ore, non meno di 6 e non più di 11.
- Dai 26 ai 64 anni: da 7 a 9 ore, non meno di 6 e non più di 10.
- Dai 65 anni in poi: 7 o 8 ore, non meno di 5 e non più di 9.
Lo studio citato dimostra che il sonno è preminente nella prima fase della vita, va man mano
a ridursi con lo sviluppo, resta costante durante l'adolescenza per poi diminuire nella vita
adulta. Nel neonato il sonno ha un ritmo polifasico: ritmico, ritmico ad onde lente, lento,
alternante. Con lo sviluppo il sonno diventa bifasico. L'anziano dorme circa 6-7 ore per notte,
ma la qualità del sonno è assai diversa da quella del giovane, in quanto è più frammentato
da momenti di veglia ed è a volte più suscettibile ai possibili disturbi ambientali.
5
M.
Albè,
“Ecco
quante
ore
dormire
ad
ogni
età”
[Online],
2015.
Disponibile
all’indirizzo:
http://www.greenme.it/vivere/salute-e-benessere/15635-tabella-giusto-sogno-eta (Consultato in data 27/06/2015).
Il sonno, secondo la “Teoria evolutiva”, si sarebbe sviluppato in relazione alle
influenze dell’ambiente, in virtù del rapporto preda-predatori: durante il sonno le
prede attraggono meno l'attenzione dei predatori, ma sono anche più vulnerabili in quanto
meno sensibili agli stimoli. Gli animali erbivori dormono per periodi brevi in modo da avere
il tempo di procacciarsi il cibo e vigilare contro i predatori. Gli animali carnivori, essendo
meno in pericolo e procurandosi più velocemente il cibo, possono dormire più a lungo. Il
gatto domestico, l'animale meno a rischio ambientale, presenta la quantità di sonno REM
maggiore (circa 200 minuti).
La “Teoria della conservazione dell'energia” si fonda sull'osservazione secondo la
quale durante il sonno si assiste ad una riduzione dell'attività metabolica del 10% e
della diminuzione della temperatura del corpo. Questo dato assume grande significato dal
punto di vista evolutivo: gli animali omeotermici, come i mammiferi e gli uccelli, hanno
bisogno di un notevole dispendio di energia per mantenere costante la temperatura interna.
Per questo motivo la riduzione di temperatura, che si verifica soprattutto durante le prime
fasi del sonno, avrebbe il significato di preservare energia. Questo processo è lo stesso che
permette a molti animali di iniziare il processo di letargo.
La “Teoria dell'apprendimento” sostiene che il sonno, e soprattutto il sonno REM,
avrebbe un ruolo determinante per la maturazione del sistema nervoso centrale, infatti,
durante la fase REM, si assiste ad un incremento dell'attività cerebrale. In studi sperimentali,
uomini sottoposti a sessioni intensive di apprendimento presentavano un aumento
significativo del sonno REM, espressione del processo di fissazione dei dati appresi nella
memoria a lungo termine. I neonati presentano una percentuale maggiore di sonno REM
rispetto agli adulti ed agli anziani, parallelamente alla maggiore capacità di apprendere.
Secondo la “Teoria della pulizia”, il cervello avrebbe una sorta di sistema addetto alla
“pulizia”, come lo è il sistema linfatico per il resto del corpo. Da recenti studi sui topi, effettuati
dall’Università di Rochester (USA) e dell'Università di Copenaghen, è emerso che, mentre
si dorme, il cervello ha un sofisticato sistema di autopulizia, che sfrutta l’espansione in
volume di una rete di canali tra i neuroni, permettendo al liquido cerebrospinale di scorrervi
in misura maggiore. Questo processo permette di smaltire prodotti di scarto del
metabolismo, come le proteine beta-amiloidi, e lavora con maggiore efficienza durante il
sonno. L'effetto ristoratore del sonno sembrerebbe essere legato, almeno in parte, a questo
meccanismo di smaltimento, da cui forse anche la ragione del torpore, della stanchezza e
dell’annebbiamento di cui si soffre per la deprivazione di sonno. La durata del sonno è
inversamente proporzionale alla grandezza del cervello, per cui un cervello più grande
impiegherà meno tempo a ripulirsi rispetto ad uno più piccolo, essendo le cellule già
predisposte all’attività di drenaggio del liquido cerebrospinale: un elefante dorme in media
tre ore al giorno, un uomo circa otto e un gatto dodici o più. Il cervello ha un’energia limitata
a sua disposizione e il lavoro di pulizia richiede un grosso sforzo, tale per cui il cervello non
può sostenere contemporaneamente tutti gli altri compiti svolti durante il periodo di veglia.
Manifestazione
singolare
all’interno
dello
stato
di
sonno,
il sogno è
un
fenomeno psichico che a lungo si è legato solo alla fase REM, teoria oggi messa in parte in
discussione. Nel 1957 Dement scoprì che la fase di sonno REM poteva verificarsi in
qualsiasi parte delle notte e una altissima percentuale di individui risvegliati durante o alla
fine di un episodio di sonno REM era in grado di esporre una esperienza onirica fortemente
connotata emotivamente, da cui si ipotizzò che l’esperienza del sogno si verificasse solo in
questa fase (ipotesi Aserinsky - Dement - Kleitmann).
Jouvet, grazie agli studi di neurobiologia sugli stati di vigilanza, apprese, invece, che
i contenuti onirici sono presenti in tutti gli stadi del sonno, nonché nella fase di veglia (rêverie,
flash onirici o sogni ad occhi aperti). Indubbiamente vi sono alcune differenze tra i sogni in
fase REM e non-REM: nella fase REM, la strutturazione spaziale e il livello di partecipazione
al sogno, il numero delle parole usate per narrarlo e la caratteristica di bizzarria emergono
in modo più netto; l’attivazione corticale del sonno non-REM è minore e comporta la
produzione di sogni con una elaborazione ridotta delle esperienze memorizzate e una
minore capacità narrativa.6
Molte le domande aperte circa i sogni. Ci si chiede se il sogno sia un effetto
secondario di qualche funzione espletata dal sonno, se sia un automatismo biologico o un
processo conoscitivo, se abbia lo scopo di respingere qualcosa di tossico accumulato
durante il giorno, se sia utile per il giorno successivo, ed altro ancora. E’ certamente ancora
un meccanismo piuttosto misterioso. Caratterizzato dalla percezione di immagini e suoni
riconosciuti come apparentemente reali dal sognante, e solo più raramente accompagnati
dalla consapevolezza della loro irrealtà, nel sogno governano le leggi dell'affettività, che
prescindono dalle norme logiche e sociali. Lo spazio e il tempo sono irreali, ovvero il
soggetto può essere contemporaneamente in posti diversi, essere attore e spettatore della
medesima scena; la persona sognata può essere vissuta dal sognatore come due diverse
persone contemporaneamente e senza contraddizione. Naturalmente tali caratteristiche
rendono assai complicata l’operazione di traduzione dei simboli nell'esperienza diurna.
Non esiste una visione unica circa il significato dei sogni; diverse le teorie proposte,
sebbene nessuna goda di pieno consenso. Un uomo in media sogna circa due ore per
notte. Quando il corpo avverte la necessità di dormire, i neuroni situati nelle vicinanze degli
occhi iniziano a mandare segnali per cui le palpebre si fanno “pesanti".
Negli studi del 1975, Jovanović, psicologo, propose una distinzione tra le allucinazioni
ipnagogiche e i sogni veri e propri7: le prime sono "esperienze fantastiche", la cui
caratteristica principale consiste nella sensazione di piena veridicità che conferisce loro la
percezione di poter essere toccate; i secondi emergono al culmine della curva del sonno,
vengono dimenticati prima del risveglio e i loro contenuti sono tanto più arcaici quanto più
profondo è il sonno. L'ipotesi è che al risveglio non si ricordi il sogno, ma l'esperienza
6
M. Jouvet, Il Sonno e il Sogno, Milano, Guanda, 1993.
7
U.J. Jovanovic, Il sonno e il sogno, tr. it. Ernesto Cipollini, Roma: Il Pensiero Scientifico editore, 1975.
allucinatoria, perché essa conserva un certo legame con la coscienza, proprio perché si
verifica in una fase transitoria del sonno. I sonnambuli, per esempio, evento che Jovanović
interpreta come una forma onirica motoria, primitiva e meno psichica, si alzano un paio di
minuti prima o dopo le fasi oniriche, scordandosi poi l’episodio. Lo stesso avviene per il
fenomeno di gridare nel sonno, altra forma che si verifica un paio di minuti prima della fase
onirica classica.
Jovanović osservò che le fasi oniriche durano in media 112,2 minuti a notte. I rapidi
movimenti oculari si dimostrano diversi da persona a persona e differenti, per la stessa
persona, all'interno dell'episodio di sonno. Nella formazione di un sogno intenso si può
arrivare ad una sincronizzazione di questi processi biologici con le scene del sogno; ma se
il sogno non ha per il sognatore un grosso significato emotivo, il roteare degli occhi non è
particolarmente connesso con il contenuto del sogno.
Osservando le onde cerebrali durante la fase REM, il sogno risulta amministrato da parti del
cervello più altamente organizzate rispetto a quelle che regolano il sonno non-REM, ma, al
tempo stesso, da regioni cerebrali funzionalmente più basse rispetto alla veglia. Jovanovic
osservò che i movimenti della mano sinistra in soggetti destrimani durante le fasi REM sono
più frequenti di quelli della destra, deducendo che nel sogno i destrimani appaiono mancini,
mentre i veri mancini diventano destri.
Nel 1977, Hobson e McCarley proposero la “teoria di attivazione di sintesi”8,
suggerendo che il sogno è sviluppato, specie nella fase REM, dalle onde ponto-genicolooccipitale che, stimolando la parte alta del mesencefalo e del prosencefalo, causano i
movimenti rapidi degli occhi. Il prosencefalo e il sistema limbico, così attivati, consentono
un recupero della memoria, la costruzione di una trama e la partecipazione emotiva al
sogno, inibendo selettivamente altre strutture cerebrali da cui dipendono la consapevolezza
8
J. A Hobson, R.W. Mccarley, “The brain as a dream state generator: an activationsynthesis hypothesis of the dream
process”, American Journal of Psychiatry, 1977, n. 134.
autoriflessiva e il ragionamento logico. Tale ipotesi si basa sul concetto che l’attività onirica
è funzionale al consolidamento dell’apprendimento cerebrale, attraverso la formazione di
“organizzazioni cerebrali interne”, che evolvono automaticamente mentre dormiamo.
Hobson, nei suoi libri “La fabbrica dei sogni” (1986) e “Sognare” (2011), arriva ad ipotizzare
che l’attività onirica sia solo funzionale ai bisogni fisiologici del corpo e che sia una sorta di
atto casuale con cui la mente elimina i prodotti inutili dell’attività catabolica. Il sogno sarebbe
costituito da meccanismi cerebrali di riscaldamento, determinati da casuali reazioni
sensoriali, al di là della storia personale, affettiva ed emozionale del sognatore. Al tempo
stesso, Hobson non crede che i sogni siano privi di valore, descrivendoli piuttosto come “il
nostro stato di coscienza più creativa, quella in cui dal caos degli elementi conoscitivi
vengono prodotte nuove configurazioni di informazioni, tanto che il tempo speso per sognare
non è sprecato.
L’ipotesi “Reverse Learning” di Crick e Mitchison, pubblicata sulla rivista scientifica
“Nature” nel 19839, propone il paragone tra il processo del sogno e un computer “off-line”. Il
cervello, nella fase REM, setaccia informazioni apprese durante il giorno, eliminando il
materiale neuronale residuo non utile a quello già immagazzinato nella memoria. Il sogno,
attività dei centri sottocorticali, avrebbe, dunque, la funzione di far dimenticare o meglio
disimparare, stimolando una sorta di “apprendimento inverso”, grazie al quale vengono
eliminate le tracce mnesiche in esubero, innescando un “processamento di selezionesilenziamento” automatico. La corteccia, infatti, non potrebbe far fronte alla grande quantità
di informazioni ricevute durante la giornata senza sviluppare pensieri "parassiti", che
andrebbero a disturbare l'organizzazione efficiente della memoria. Durante il sonno REM,
le connessioni indesiderate all’interno delle reti corticali sono presumibilmente spazzate via
da un meccanismo di impulsi, che bombardano la corteccia delle zone sub-corticali.
9
F. Crick; G. Mitchison, “We dream to forget. The function of dream sleep”, Nature n. 304, 1983, pp. 111-114.
Gli studi di Solms10 e Kaplan (2000) mettono in discussione l'ipotesi di Hobson
secondo la quale sia il tronco encefalico la fonte dei segnali che vengono interpretati come
sogni. Solms sostiene l'idea del sogno come funzione complessa di strutture cerebrali. Il
sonno REM e l’attività onirica apparterrebbero a strutture anatomiche differenti11: le strutture
del tronco encefalico regolerebbero i meccanismi fisiologici del sonno REM, mentre il
meccanismo del sogno sarebbe generato nelle aree del cervello anteriore ed in particolare
nella parte inferiore dei lobi parietali e di quella medio-basale dei lobi frontali. Le strutture
anatomiche del cervello anteriore interessate al fenomeno della costruzione del sogno
sarebbero quelle impegnate nelle funzioni delle emozioni e della memoria (il sistema
limbico, comprese le componenti limbiche delle aree frontali e temporali, cioè la giunzione
occipito-temporo-parietale e il sistema delle aree visive).
Solms verifica le sue ipotesi studiando alcuni pazienti con vari danni al cervello: coloro che
presentavano danni alle aree associative parieto-temporo-occipitale avevano smesso di
sognare, mentre i pazienti affetti da lesioni del tronco encefalico continuavano a sognare
(G. Marzi, 2005). Inoltre, se la lesione riguardava solo la corteccia associativa temporooccipitale, i sogni perdevano la loro componente allucinatoria ma non scomparivano, mentre
negli individui in cui erano compromesse le strutture associative limbiche non vi era più la
capacità di distinguere i sogni dalla realtà, trovandosi, così, a vivere in una situazione
continua di sogno.
Solms, le cui teorie vengono riprese da Foulkes (1985), Abe, Ogawa, Nittono, Hori (2008) e
Domhoff (2010), sostanzialmente riduce la visione analitica classica del sogno come
10
Mark Solms è sia neuroscienziato che psicoanalista; docente di neuro-psicoanalisi presso il Dipartimento di Psicologia
dell’Università di Città del Capo, ha ottenuto numerosi premi per il suo pionieristico lavoro in neuro-psicoanalisi, incluso
l’International Psychiatrist Award dal the American Psychiatric Association and Best Science Writing Awards. Fondatore
della “International Neuro-Psychoanalysis Society” il cui organo di stampa è la rivista Neuro-Psychoanalysis (disponibile
online all’indirizzo http://www.neuro-psa.org.uk/npsa/).
11
M. Solms, “Dreaming and REM sleep are controlled by different brain mechanisms”, Behavioral and Brain Sciences,
Cambridge Journals n. 23, 2000, pp. 793-1121.
struttura difensiva, pur confermando l’esistenza di un inconscio, che definisce “un fenomeno
biologico utile al funzionamento cognitivo, conscio ed inconscio.” (Rivista Mente & Cervello
2003).
Combinando
le
ricerche
di
Hobson
e
Solms,
la “Teoria
di
continual-
activation”12, presentata da Zhang, parte dal presupposto che sogno e fase REM siano
controllati da meccanismi cerebrali differenti. Il ricercatore spiega i sogni come il risultato di
una “attivazione cerebrale e di un lavoro di sintesi”. La funzione del sonno sarebbe quella di
trasferire le informazioni dalla memoria a breve termine a quella a lungo termine, sebbene
la teoria del “consolidamento” della memoria non trovi una conferma. Il sonno non-REM
tratterebbe la memoria consapevole esplicita (memoria dichiarativa), mentre il sonno REM
si occuperebbe della memoria implicita, inconsapevole (memoria procedurale). Zhang
immagina che, durante la fase REM, la parte inconscia del cervello sia occupata a
processare la memoria procedurale, mentre la parte consapevole del cervello sia
“disconnessa”. Un meccanismo di attivazione continua genera un fiume di dati nella
memoria immagazzinata dalla parte consapevole del cervello. Attraverso il sogno, la
memoria recente sarebbe assemblata con il materiale già presente, in modo da essere
compatibile con quella già contenuta nel cervello13.
Tarnow suggerisce che sognare aiuta a consolidare i ricordi immagazzinati nella
memoria a breve termine in quella a lungo termine: la fase REM serve proprio ad
organizzare i ricordi, mettendoli in relazione con altri passati, associandoli e catalogandoli.
I ricordi sarebbero memorizzati nella memoria a lungo termine, secondo ciò che in essa è
già presente. Studi recenti dimostrano che uno stato di privazione da fase REM provoca
12
J. Zhang, “Continual-Activation Theory of Dreaming”. Dynamical Psychology [Online]. 2005. Disponibile
all’indirizzo: http://www.goertzel.org/dynapsyc/2005/ZhangDreams.htm (Consultato in data 01/07/2015).
13
J. Zhang, “Memory precess and the function of sleep”. Journal of Theoretics [Online]. 2004, Vol. 6-6. Disponibile
all’indirizzo: http://journaloftheoretics.com/Articles/6-6/Zhang.pdf (Consultato in data 02/07/2015).
problemi di memoria, irritabilità, difficoltà di attenzione, rendendo la teoria di Tarnow
particolarmente calzante14.
Ernest Hartmann15, direttore dello Sleep Disorders Center di Boston, insieme ad altri
collaboratori ha elaborato la “Teoria Contemporanea del sogno”16, secondo la quale i sogni
permettono al sognante di integrare “cose e pensieri” che, altrimenti, verrebbero dissociati
nella fase conscia. Questo può avvenire perché il cervello sembra produca e distrugga in
continuazione connessioni neurali, fondamento delle nostre attività mentali. Durante questa
attività è possibile cogliere un continuum lungo il quale le connessioni si fanno sempre più
diradate ed imprecise: da un lato ci sono le connessioni della veglia, che lungo il
procedimento si fanno più vaghe, tanto che le attività mentali diventano più approssimative;
dal lato opposto ci sono i sogni, guidati dalle emozioni del sognante. Più questi è pervaso
da un’emozione forte, più individuabile sarà il senso dei suoi sogni. Il sogno sarebbe
un’interconnessione di ricordi legati ad altri, capace di rendere blanda l’azione disturbante
dell’emozione e funzionare come un “cancellino”: chi vive un trauma nei primi giorni torna a
riviverlo nei sogni in maniera altrettanto forte, ma, con il passare del tempo, inizia a sognare
eventi diversi, seppur traumatici, che possono richiamare anche altri eventi spiacevoli del
passato, come se i sogni inglobassero e si connettessero con altro materiale più neutro,
presente nella memoria del soggetto, perdendo il collegamento esplicito con l’evento
traumatico vissuto.
14
E. Tarnow, “How Dreams And Memory May Be Related”. Neuro-Psychoanalysis [Online]. 2003, vol. 5, n. 2.
Disponibile all’indirizzo: http://cogprints.org/2068/1/DreamsAndMemoryTarnow.pdf (Consultato in data 07/07/2015).
15
Membro della Boston Psychoanalytic Society, Past President della International Association for the Study of Dreams
(IASD), primo Editor-inChief della rivista della IASD “Dreaming”, Professore di Psichiatria alla Tufts University School
of Medicine ed ex-Direttore dello Sleep Disorders Center al Newton-Wellesley Hospital.
16
E.
Hartmann,
“Dreaming,
the
contemporary
theory”
[Online].
Disponibile
all’indirizzo:
http://web.me.com/ernesthartmann/ERNEST_HARTMANN_MD/HOME_files/Dreaming%20The%20Contemporary%
20 Theory%202010%20Powerpoint.ppt (Consultato in data 05/07/2015).
Griffin e Tyrrell17 sostengono che il sogno sia di vitale importanza per la salute
mentale, anche se ricordarlo non lo è. Tale teoria, detta "Teoria di adempimento
dell'aspettazione di sognare"18, suggerisce che, sognando metaforicamente si completano
modelli di aspettazione emotiva e, di conseguenza, si abbassano i livelli di stress, da cui
deriva la diminuzione della produzione del cortisolo19.
Per Foulkes e Antrobus, il sogno è un processo mentale complesso che svolge una
essenziale funzione adattiva e l’inconscio è un livello della cognizione riferito allo stato di
consapevolezza, anziché essere uno spazio energetico pulsionale che preme per
emergere. Esperimenti neuroelettrofisiologici (Rodolfo Llinàs, Mircea Steriade, Denis Parè)
affermano che la plasticità neuronale e le reti di interconnessioni (connectivity patterns) del
cervello creano con il sogno un’esperienza mentale simile a quella di veglia e poi la
integrano all’interno della struttura di personalità, decodificando la parte rimasta non
consapevole e definibile inconscia.
L’articolo di Wamsley20 offre una panoramica riassuntiva delle teorie ad oggi più
quotate, ponendo l’accento su alcuni concetti chiave a lui cari. Secondo l’autore, i sogni e lo
stato di coscienza attivo durante la veglia non sono necessariamente generati da
meccanismi distinti, contrariamente alle credenze più classiche: “Anche quando non stiamo
apparentemente facendo nulla, come durante gli stati di riposo, di sonno, nelle
fantasticherie, il cervello continua a elaborare informazioni: durante la veglia la mente
genera pensieri, progetti per il futuro e immagina scenari fittizi; durante il sonno, quando gli
input sensoriali sono ridotti, la mente si rivolge a pensieri e immagini che chiamiamo
17
Joe Griffin è ricercatore psicologo, psicoterapeuta, co-autore con Ivan Tyrrell di Human Givens; Ivan Tyrrell è scrittore,
psicoterapeuta e direttore del Mindfields College nel Regno Unito.
18
J. Griffin, L. Tyrrell, Dreaming Reality: How Dreaming Keeps Us Sane, or Can Drive Us Mad, Human Givens
Publishing, 2004..
19
J. Griffin, “The Origin of Dreams: How and why we evolved to dream”, in The Therapist, vol IV n. 3, 1997.
20
Erin J. Wamsley lavora al Department of Psychiatry, Center for Sleep and Cognition, Harvard Medical School and
Beth Israel Deaconess Medical Center, Boston, Ma, Usa.
“sognare”. Lungi dall'essere una distrazione senza senso, il contenuto di queste esperienze
soggettive fornisce una fonte importante di informazioni sulle attività della mente e del
cervello a riposo. Sia nella veglia che nel sonno ha luogo un'esperienza spontanea che
combina in nuovi scenari frammenti di memoria recente e remota. Tali esperienze coscienti
possono rappresentare il consolidamento della memoria recente in forme di stoccaggio a
lungo termine, un processo adattivo che serve ad estrarre conoscenze generali sul mondo
e rispondere in modo adattivo ad eventi futuri 21. Lo studioso prosegue elencando prove
emergenti a sostegno di quanto affermato. Le esperienze coscienti durante il sonno (vale a
dire, i sogni), classicamente ritenute un fenomeno del tutto distinto dal pensiero spontaneo
e dalle immagini della veglia, possono essere meglio concettualizzate come una estensione
naturale della coscienza vigile (Wamsley e Stickgold, 2010; Domhoff, 2011; Horikawa,
2013). Nella fase di dormiveglia, come nel sonno, la mente è al lavoro, elaborando le
situazioni e le preoccupazioni della giornata, consolidandole nella memoria (Plihal e Born,
1997; Mednick, 2002; Tucker, 2006) ed integrando le nuove informazioni con le conoscenze
esistenti (Tamminen, 2010; Lewis e Durrant, 2011), mescolandole con esperienze passate,
al fine di pianificare al meglio il futuro (Wilhelm, 2011). Lo studio sistematico di queste
esperienze soggettive, in tutti gli stati di coscienza, potrebbe rivelarsi fondamentale per una
più ampia comprensione del funzionamento del cervello durante gli stati “offline”.
Nonostante la proliferazione di dizionari sul simbolismo onirico, Wamsley sottolinea che non
c'è stato alcun riscontro empirico di una particolare decodifica di tali simboli.
La validità della teoria neurobiologia di Hobson, secondo la quale il sogno è prodotto
all’interno del tronco cerebrale pontino nella fase REM è stata negata dai nuovi studi della
comunità scientifica, che hanno evidenziato come i sogni avvengono anche al di fuori della
21
in
E. Wamsley, “Dreaming, waking conscious experience, and the resting brain: report of subjective experience as a tool
the
cognitive
neurosciences”.
Frontiers
in
psychology
[Online].
2013.
Disponibile
http://journal.frontiersin.org/article/10.3389/fpsyg.2013.00637/full (Consultato in data 05/07/2015).
all’indirizzo:
fase REM, incluse le fasi più profonde del sonno. Teorie più recenti (Cicogna e Bosinelli,
2001; Nir e Tononi, 2010; Wamsley e Stickgold, 2010; Perogamvros e Schwartz, 2012),
suggeriscono che la coscienza del sogno è generata dallo stesso substrato neurale di base
che supporta l'esperienza soggettiva spontanea durante gli stati “offline”, ovvero nei
momenti di “resting wakefulness” (dormiveglia, quiescenza).
Una ragione per cui il fenomeno del sogno è stato a lungo trattato separatamente dalla
esperienza cosciente della veglia è che lo si è sempre considerato un avvenimento bizzarro,
allucinatorio, cognitivamente carente. Certamente, l'esperienza cambia mentre ci si muove
attraverso i diversi stati di coscienza, ma forse i cambiamenti più notevoli durante il sogno
sono un aumento della vividezza delle immagini, insieme ad una consapevolezza attenuata
del mondo esterno (Hobson, 2000). Vi sono, altresì, drastici cambiamenti di neuromodulazione, elettrofisiologia e attivazione cerebrale regionale. Tuttavia, non ci sono prove
che suggeriscano la presenza di un meccanismo fondamentalmente diverso tra esperienza
cosciente durante il sogno ed esperienza cosciente durante la veglia. Al contrario, insiste
Wamsley, i dati disponibili indicano che la forma e il contenuto dell'esperienza cosciente di
veglia offline e di sonno creano rapporti di sovrapposizione così simili da essere
indistinguibili: immagini vivide e anche allucinatorie, per esempio, sono a volte riportate
anche in racconti di sogni ad occhi aperti (Foulkes e Scott, 1973; Foulkes e Fleisher, 1975);
i sogni nel sonno non sono necessariamente più bizzarri rispetto alle fantasie ad occhi
aperti: quest’ultime possono essere altrettanto stravaganti e ricche di improvvisi
cambiamenti discontinui di argomento (Wollman e Antrobus, 1986; Reinsel 1992). La
cognizione durante il sonno può essere sorprendentemente logica e coerente, incluse le
auto-riflessioni, le pianificazioni, l’attenzione focalizzata (Kahan, 1997; Kahan e Laberge,
2011). Nel sonno REM, le esperienze oniriche sono spesso più lunghe, più vivide, più
dettagliate e più bizzarre rispetto a quelle negli stadi non-REM, ma, al tempo stesso, è
possibile teorizzare una sovrapposizione tra le due esperienze (Foulkes, 1962, 1967;
Antrobus, 1995; Cicogna, 1998; Smith, 2004. Wamsley, 2007). Studi di neuroimaging dopo
il 1950 (EEG, PPET, fMRI) hanno evidenziato che il cervello e la mente non sono
completamente spenti durante il sonno: l’attività cerebrale nel sonno REM assomiglia a
quella dell’attività cerebrale della veglia; le fasi più profonde del sonno non-REM mostrano
che l'attività metabolica regionale è mantenuta in regioni selezionate (Nofzinger, 2002;
Peigneux, 2004); aree cerebrali attive durante i periodi di veglia in condizione di riposo,
ovvero quando le persone non sono intente a processare stimoli esterni, rimangono
relativamente attive anche durante il sonno, sia REM che non-REM, comprese l'area mediofrontale e l'area medio-temporale coinvolte nella elaborazione della memoria (Buckner,
2008) (Domhoff, 2011). Durante la veglia è attiva una rete di default, associata con la
generazione del pensiero cosciente e delle immagini (Mason, 2007; Andrews-Hanna, 2010;
Andrews-Hanna, 2012), che risulta attiva anche in condizioni di controllo sensoriale ridotto,
aumentando pensieri stimolo indipendenti (Andrews-Hanna, 2010). Individui che hanno
riportato pensieri irrilevanti del passato e del futuro nel corso di una condizione di riposo
hanno mostrato un aumento della connettività funzionale tra le strutture del lobo temporale
mediale e di altre componenti della rete di default di cui sopra (Andrews-Hanna, 2010).
Infine, l'attivazione della suddetta rete è maggiore negli individui con una forte propensione
verso i sogni ad occhi aperti (Mason, 2007), a dimostrazione di una comunanza strutturale
e funzionale tra la “modalità standard” di veglia in condizioni di riposo e un pattern di
attivazione funzionale durante il sonno. La relazione dell’attività di rete default con lo stato
del sogno durante il sonno non è ancora stata testata direttamente, ma ci sono prove
preliminari del fatto che alcune regioni di questa rete contribuiscano alla formazione dei
sogni, a dimostrazione della presenza di una rete condivisa per la generazione di esperienze
soggettive spontanee e altri stati di coscienza. Nella veglia e nel sonno, l’esperienza
spontanea cosciente riflette memorie del passato e pianificazioni per il futuro. L'ipotesi è che
una funzione del cervello durante il riposo sia quella di utilizzare ricordi del passato per
costruire simulazioni di future esperienze possibili, così da rafforzare la preparazione agli
eventi futuri (Schacter, 2007). A sostegno di questa ipotesi, va riportato che i pazienti con
danno bilaterale dell'ippocampo, struttura nel lobo temporale mediale essenziale per la
formazione di nuove memorie, mostrano non solo deficit di memoria, ma anche una
compromissione della capacità di immaginare scenari fittizi ed eventi futuri (Hassabis, 2007;
Race, 2012). Queste osservazioni suggeriscono, secondo Wamsley, che durante i periodi
di riposo, frammenti di esperienze passate vengono riattivati nella mente e combinati in
nuovi scenari immaginati, ricchi di possibili eventi futuri. Parecchie evidenze suggeriscono
che l'esperienza del sogno può similmente attivare l’elaborazione di memorie del passato,
così come utilizzare la memoria per simulare eventi nel futuro. Vi è oggi forte evidenza del
beneficio del sonno per il “consolidamento” delle informazioni di nuova acquisizione e per le
prestazioni della memoria procedurale e dichiarativa (Stickgold, 2000; Walker, 2002) (Plihal
e Born, 1997; Ellenbogen, 2006; Tucker, 2006). L'elaborazione da parte della memoria
durante il sonno sembra riflettersi direttamente nell'esperienza cosciente del sogno: anche
se le esperienze passate sono raramente, se non mai, riprodotte nei sogni nella loro forma
completa e originale, la maggior parte dei resoconti di sogni contengono almeno un
elemento che può essere fatto risalire ad una memoria recente specifica (Fosse, 2003). Se
è improbabile che il contenuto dei sogni possa essere determinato esclusivamente da
processi relativi alla memoria [per esempio, il sogno può anche essere influenzato da
sistemi motivazionali e di ricompensa (Pennartz, 2004; Perogamvros e Schwartz, 2012)],
appare chiaro il contributo del consolidamento della memoria durante il sonno alla
costruzione dell'esperienza vissuta all'interno del sogno. Nel loro insieme, queste
osservazioni, asserisce con forza Wamsley, suggeriscono l'ipotesi che sia le immagini
spontanee durante la veglia in condizioni di rilassamento, sia i sogni durante il sonno
possano essere influenzati dai medesimi processi cerebrali.
Il sogno rimane centro attrattore delle Psicoterapie, che lo considerano una fonte
importante da cui attingere per entrare meglio nelle dinamiche mentali del paziente. Benelli,
portavoce del pensiero psicoanalitico moderno, chiarisce che i sogni, più che distorti impulsi
nascosti, vanno considerati pensieri che riguardano avvenimenti della vita da svegli, più che
appagamento potrebbero essere completamento e ristrutturazione di una organizzazione
psicologica. Il sogno permette la comunicazione del mondo interno del sognatore, che
esprime anche tramite il sogno, il desiderio e l’angoscia di conoscere se stesso 22.
Scrive Sergio Lombardo: “L'interpretazione psicoanalitica classica dei sogni come
appagamento surrogato di desideri proibiti, non si adatta ai risultati sperimentali ottenuti
dalla fisiologia del sonno, non solo per quanto riguarda la mancanza dei caratteri di malattia
che quella interpretazione implicava, ma anche perché non si capirebbe la necessità di un
processo del sonno a cicli successivi allo scopo di appagare un desiderio in forma
allucinatoria, specialmente qualora si tratti di un desiderio covato fin dall'infanzia e
descrivibile in termini erotici infantili. Più che realizzare in surrogato desideri infantili, il sogno
sembra creare per tentativi qualcosa di assolutamente nuovo, sembra affrontare con
maggior successo un evento enigmatico, un problema senza formalizzazione, una
situazione imprevista. Il fatto che la fase REM non compare quando l'organismo discende
dalla veglia al sonno profondo, ma soltanto quando risale verso una maggiore eccitazione,
è in accordo con l'ipotesi proposta. La discesa verso il sonno è un processo di disgregazione
dell'Io, inteso come programma comportamentale generale, ovvero come programma di
utilizzazione del mondo. Disgregazione necessaria per far affiorare le informazioni
accumulate durante il giorno e accantonate nel preconscio perché inutilizzabili, non
essendovi istruzioni adatte nel programma funzionale della percezione cosciente. Nel
22
E. Benelli, Per una nuova interpretazione dei sogni, Bergamo: Moretti & Vitali, 2007.
risalire dal sonno passivo (SWS)23 al sonno attivo (REM) durante il processo di ricostruzione
dell'iniziativa di fronte al materiale sconosciuto accumulato nel preconscio, poco prima della
soglia di risveglio avviene la completa caduta del tono muscolare e la comparsa delle
allucinazioni oniriche. La caduta del tono muscolare blocca il comportamento esterno e
garantisce che il comportamento interno di ristrutturazione dei programmi, rispetto alle
modificazioni del mondo, non appaia sul piano esecutivo. 24
Concludendo, non si trova ad oggi una descrizione accettata su quale sia la zona
trigger che, stimolata, dia origine al sogno, né sulle vie nervose da esso utilizzate. I risultati
scientifici qui esposti non sono in grado di fornire una spiegazione esaustiva del fenomeno
sogno, ma sicuramente ne evidenziano la funzione evolutiva: fenomeno che sembra
permettere la messa in funzione della capacità di autoapprendimento, funzione chiave per
la sopravvivenza; manifestazione che sembra consentire una rielaborazione continua dei
contenuti, sia consci che inconsci, favorendo un equilibrio psico-emotivo; processo mentale
complesso che pare svolgere una funzione adattiva per la memoria, l’apprendimento e le
capacità cognitive di problem solving; meccanismo dal ruolo primario nello sviluppo
dell’organizzazione psicologica di personalità.
23
Il sonno senza sogni, N-REM, presenta durante gli stadi più profondi un particolare tipo di onde di grande ampiezza,
lente e sincronizzate (onde delta), che rivelano una fase contrapposta simmetricamente al sogno: la fase SWS (Slow
Waves Sleep) molto lunga nel primo periodo, progressivamente più corta nei successivi.
24
S. Lombardo, “Il sogno - una funzione biologica indicibile”, in Rivista di Psicologia dell'Arte, A. II, n.2, giugno 1980,
pp.15-29.