I sistemi viventi sono caratterizzati da un’enorme
varietà reazioni chimiche la maggior parte delle quali è
mediata da una serie di eccezionali CATALIZZATORI
BIOLOGICI :
GLI ENZIMI
L ’ ENZIMOLOGIA E ’
UNA
PARTE
DELLA
BIOCHIMICA
CHE
STUDIA GLI ENZIMI
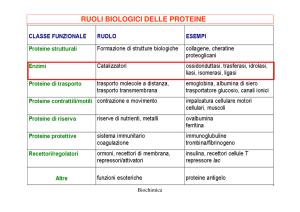
RUOLO
DEGLI
ENZIMI:
catalizzatori
biologici
Gli enzimi sono catalizzatori biologici
Capaci di aumentare la velocità di una reazione,
anche più di un milione di volte, senza subire
trasformazioni durante l’intero processo.
Per la maggior parte sono proteine la cui attività
catalitica dipende dall ’ integrità della loro
conformazione proteica nativa
QUALSIASI molecola che
lega una proteina prende il
nome di LIGANDO
I ligandi possono essere
piccole
molecole
o
macromolecole
Se il ligando muta nel
tempo
si
chiamerà
SUBSTRATO e PRODOTTO
il risultato del mutamento
2 H2O2
CATALASI
Fe3+
emoglobina
2H2O + O2
Reazione lenta
Reazione 1.000 volte più veloce se aggiungiamo ione ferrico
Fe3+
Reazione 1.000.000 di volte più veloce in presenza di
emoglobina, proteina contenente ferro
Reazione 1 miliardo di volte più veloce in presenza di
un ENZIMA: CATALASI presente in molte cellule
1) Il catalizzatore sebbene partecipi alla reazione, rimane
INALTERATO durante il processo
2) Il catalizzatore cambia la velocità del processo, ma non
influisce sulla posizione di EQUILIBRIO della reazione
3) Un processo TERMODINAMICAMENTE FAVORITO non
viene reso più favorito dal catalizzatore
OTTIMIZZANO LA RESA:
a. Svolgono solo le reazioni necessarie
b. Impediscono reazioni indesiderate (specificità di
substrato)
c. Portano a termine le reazioni in tempi brevi
d. Sono indipendenti dall ’ ambiente (temperatura,
pressione)
e. Riutilizzano della stessa “macchina” per molte volte
MINIMIZZANO GLI SPRECHI:
a. Non producono sottoprodotti (specificità di reazione)
b. Minimizzano il dispendio energetico
-
HANNO CAPACITA’ DI REGOLAZIONE
che risiede nelle loro proprietà strutturali
L’ENZIMA LEGA IL SUBSTRATO IN UNA REGIONE
PROTEICA SPECIFICA
Il potere catalitico di un enzima è dovuto, per
la maggior parte, al legame dei substrati in
un corretto orientamento
Il sito attivo è una regione della proteina che lega
il substrato e contiene residui aminoacidici che
partecipano alla catalisi
Caratteristiche comuni:
1. Occupa una parte relativamente piccola del volume totale
della molecola enzimatica
2. Entità tridimenzionale formata da gruppi che derivano da
parti diverse della sequenza aminoacidica lineare
3. I substrati si legano all’enzima mediante un certo numero di
attrazioni deboli
4. Sono cavità o fenditure (acqua normalmente esclusa se non
è un reagente, carattere non polare ma può contenere
residui polari)
5. La specificità del legame dipende dalla precisa disposizione
degli atomi in un sito attivo
COME GLI ENZIMI FUNZIONANO DA CATALIZZATORI
Le forze attraverso le quali i substrati e le altre molecole si
legano agli enzimi sono lo stesso tipo di forze che regolano
la conformazione delle proteine
L’enzima lega una o più molecole di SUBSTRATO e forma
un complesso enzima-substrato
COMPLEMENTARIETA’ GEOMETRICA ED
ELETTRONICA
E. Fischer nel 1894 propose il modello chiave-serratura
che descrive in modo adeguato la specificità degli enzimi.
Un’elaborazione successiva (D. Koshland, 1958) consentì
di aumentare le conoscenze sui meccanismi catalitici: il
modello
ad
adattamento
indotto.
*
Un’elaborazione successiva (D. Koshland,
1958) consentì di aumentare le
conoscenze sui meccanismi catalitici: il
modello ad adattamento indotto.
L’adattamento indotto implica distorsioni a carico sia
dell’enzima che del substrato. Queste possono essere
localizzate o comportare una grossa variazione
conformazionale
dell’enzima
come
nel
caso
dell’esochinasi.
ENZIMA DIIIDOFOLATO
REDUTTASI assieme al
suo substrato NADP+
Tasca dell’enzima
complementare al substrato
Processo detto di
adattamento indotto
La complementarietà proteina
ligando è di rado così perfetta.
Le interazioni tra una proteina e
un ligando sono più spesso
accompagnate da modificazioni
conformazionali di una o di
entrambe le molecole.
Anche i siti di legame del substrato subiscono cambiamenti
conformazionali
Variazione
conformazionale
indotta
nell’esochinasi
da parte del
glucosio
Gli enzimi non si limitano a posizionare e distorcere
i loro substrati ma mediante specifici residui
aminoacidici partecipano all’evento catalitico
Quando il processo è
completato l’enzima
rilascia il PRODOTTO(i) e
ritorna al suo stato
originale, pronto per un
altro CICLO CATALITICO
Oltre al substrato molti
enzimi
legano
altre
molecole necessarie per
lo svolgimento della loro
attività catalitica
Gran parte degli
enzimi è costituita da
una parte proteica ed
una non proteica
Come per l’emoglobina e la miosina, che riescono a svolgere
la loro capacità di legare ossigeno solo se contengono il gruppo
EME, alcuni enzimi riescono a svolgere la loro attività
catalitica, grazie a questi gruppi FUNZIONALI
Gran parte degli enzimi è costituita da una parte proteica
inattiva ed una non proteica che ne determina l’attività
APOENZIMA + COFATTORE = OLOENZIMA
COFATTORE
parte essenziale del sito attivo a cui si legano
IONI ESSENZIALI
COENZIMI
Molecole organiche
complesse
ATTIVATORI
COSUBSTRATI
legati debolmente
Legati debolmente
GRUPPI PROSTETICI
Legati fortemente
I cosubstrati come i substrati sono parte essenziale del sito
attivo
Si trasformano durante la reazione e si dissociano dal
sito attivo alla fine della reazione. Sono riconvertiti poi
nella forma iniziale attraverso altre reazioni enzimatiche.
GRUPPI METABOLICI PENDOLARI
ATP + glucosio
ADP + glucosio-fosfato
L’ADP viene rifosforilato
ad ATP in altre reazioni
Il glucosio-fosfato viene
ulteriormente trasformato
è permanentemente legato
all’apoenzima
Ed affinché l’oloenzima continui ad
essere cataliticamente attivo
il
gruppo prostetico deve essere
rigenerato nella sua forma iniziale,
così come un qualsiasi residuo del
sito attivo che partecipi alla catalisi
INSOLUBILI
Vitamin A
SOLUBILI
Tiamina (B1)
Riboflavina (B2)
Niacina (B3)
Acido Pantotenico
(B5)
Piridoxina (B6)
Biotina
Cobalamina (B12)
Acido Folico
Acido Ascorbico
Vitamin D
Vitamin E
Vitamin K
Le VITAMINE che
vengono assunte
con l’alimentazione
sono composti
organici essenziali
ai processi biologici
degli organismi
superiori che non
possono sintetizzarli
Molte vitamine sono precursori di
COFATTORI (vitamine idrosolubili)
altre pur giocando un ruolo
fisiologico essenziale non lo sono
(vitamine liposolubili).
Conosciuta anche come Vit.B1
È rapidamente convertita nella sua forma
biologicamente attiva tiamina pirofosfato
1,0-1.5 mg/giorno
beriberi
nell’individuo adulto
Tiamina
pirofosfatasi
METALLOENZIMI
Molti enzimi contengono ioni metallici,
solitamente legati con legame di coordinazione a catene laterali
aminoacidiche e talvolta legati a gruppi prostetici
I metalli funzionano come i
coenzimi, facendo assumere ai
metalloenzimi
proprietà
catalitiche che non potrebbe
possedere in loro assenza.
In molte reazioni enzimatiche certi ioni sono necessari per
l’EFFICIENZA CATALITICA.
Molti enzimi che accoppiano l’IDROLISI dell’ ATP ad altri
processi biochimici richiedono Mg2+ per funzionare in modo
efficiente.
Il complesso Mg-ATP è substrato migliore dell’ATP da solo
L’atomo di zinco
serve da
catalizzatore
metallico per
promuovere
l’idrolisi. Il
legame scisso è
indicato da un
piccolo cuneo
rosso
IN ALTRI CASI IL METALLO PRESENTE NELL’ENZIMA
SERVE DA AGENTE OSSIDO-RIDUTTIVO
Quasi tutti gli enzimi sono nominati aggiungendo il suffisso –asi al
nome del substrato od ad una frase che specifica la reazione catalitica
dell’enzima
UREASI
catalizza l’idrolisi dell’urea
ALCOL DEIDROGENASI
catalizza l’ossidazione degli alcoli alle loro corrispondenti aldeidi
L’esistenza di un enorme numero di enzimi ha portato la
Commissione degli Enzimi dell ’ Unione Internazionale di
Biochimica ad elaborare di un sistema razionale di
nomenclatura
International Union of Biochemistry and Molecular Biology (IUMB)
1)
OSSIDOREDUTTASI ossidoriduzione
2)
TRANSFERASI
trasferimento di gruppi funzionali
3)
IDROLASI
idrolisi
4)
LIASI
5)
ISOMERASI
6)
LIGASI
rimozione o aggiunta di un gruppo al doppio legame o altre
scissioni
riarrangiamenti intramolecolari
fusione di due molecole
Classificazione degli Enzimi
http://www.expasy.org/enzyme
Nome raccomandato
e nome sistematico.
Numero di
Classificazione:
Numero a quattro
cifre preceduto da EC
(Enzyme Commission:
la prima cifra indica
la classe, la seconda
la sotto-classe, la
terza la sotto-sottoclasse, la quarta il
numero seriale).
Ad ogni enzima sono associati 4 NUMERI
EC 6.4.1.1.
Numero di serie
Classe
Sottoclasse
Sotto-sottoclasse
PEPTIDIL-L-AMMINOACIDO-IDROLASI E.C.3.4.17.1
CARBOSSIPEPTIDASI A
Enzyme
commition
Glucosio-6-P
glucosio + Pi
Glucosio-6-fosfatasi
Estremità non
riducente
Rottura di un legame covalente mediato
dall’ACQUA
Rottura di un legame covalente mediato dal
FOSFATO
32
Velocità di reazione.
Consideriamo una generica reazione
Esprimiamo perciò la velocità di un processo chimico in
un certo periodo di tempo tramite il rapporto tra la
variazione di concentrazione di un reagente o di un
prodotto ed il valore di quell' intervallo di tempo. L'
espressione matematica della velocità di reazione è la
seguente:
Riferendoci ad una
concentrazione
generica C La velocità
di reazione v può
essere determinata
dalla variazione della
quantità di C
nell'unità di tempo:
v = d C / d t
Misure sperimentali
34
35
*
La velocità è dunque
dipendente dalla
concentrazione del
reagente
36
Se v aumenta proporzionalmente al crescere della
concentrazione di C, si dice che la reazione è del
primo ordine rispetto a C; la K di velocità è del I°
ordine ed è data dalla pendenza della retta v = K C
C
P
Misure sperimentali a
diversa concentrazione di C
Cinetica
temporale
a) i punti rossi
rappresentano
dati
ottenuti
sperimentalmente
misurando
la
velocità
a
concentrazioni
diverse di C; la K è
data
dalla
pendenza
della
retta che si ottiene
ottimizzando la retta
individuata dai punti
sperimentali; i punti
non giacciono tutti
esattamente
sulla
retta dato che sono
soggetti
a
errori
sperimentali.
37
EQUAZIONE
DI
VELOCITA’
V = K [C]
*
38
39
COLLISIONE TRA DUE MOLECOLE
Urto
insufficiente
Rottura di
legami e
formazione
di nuovi
legami
Urto
sufficiente
o efficace
Aumento della
concentrazione
Aumento della temperatura
40
Fattori che influenzano la velocità di reazione
Si rompono e si formano legami
v dipenderà dalla natura dei legami
OMOGENEE
ETEROGENEE
- fasi uguali
- fasi diverse
NATURA DEI REAGENTI
k
CAPACITA’ DEI REAGENTI
DI INCONTRARSI
con la concentrazione varia
il numero di collisioni
possibili
CONCENTRAZIONE
DEI REAGENTI
41
NO2Cl + Cl
a)
NO2 + Cl2
Collisione che non può produrre una molecola di Cl2
b) Collisione che produce Cl2 e NO2
Se venisse raddoppiata la [NO2Cl] si avrebbe un numero
doppio di possibili collisioni così come se si
raddoppiasse la [Cl]. Questo significa che la velocità
dipenderà da entrambe le concentrazioni elevate alla
prima potenza.
42
velocità = K[Cl][NO2Cl]
con la T varia il
numero di urti efficaci,
dato che cambia
l'energia cinetica
TEMPERATURA O
PRESSIONE DEL
SISTEMA
Particolari sostanze non
richieste dalla
stechiometria della
reazione
43
Da cosa è determinata la velocità di reazione?
La TERMODINAMICA studia le trasformazioni
ENERGETICHE delle reazioni ma non dice niente
riguardo alla velocità di reazione, dice soltanto se
una reazione può avvenire spontaneamente o no
(reazione ESOERGONICA o ENDOERGONICA )
Ciò che può avere importanza nel determinare la
velocità di un processo è quello che accade
nella TRANSIZIONE dei reagenti a prodotti
44
Coordinate di reazione
45
Stato di transizione
I reagenti formano il
complesso attivato
G
Allo STATO DI TRANSIZIONE
i
reagenti
che
hanno
superato
lo
SCOGLIO
ENERGETICO
possono
evolvere verso la formazione
dei prodotti o tornare alla
formazione dei reagenti
A+B
P+ Q
G‡ = ENERGIA LIBERA DI ATTIVAZIONE
Energia necessaria per portare le molecole dei
reagenti allo stato di attivazione
46
ENERGIA DI ATTIVAZIONE G0‡
Energia libera supplementare che le molecole devono
possedere per raggiungere lo STATO DI TRANSIZIONE
Se la BARRIERA ENERGETICA da
superare è alta, solo una piccola
frazione di molecole sarà in
grado di oltrepassarla
COME AGGIRARE L’OSTACOLO
Aumentando l’energia
cinetica delle molecole
Abbassando la barriera
energetica
Un CATALIZZATORE abbassa la barriera energetica di una
reazione, aumentando di conseguenza il numero di molecole
che hanno abbastanza energia per raggiungere lo stato di
transizione e rendendo la reazione
più veloce in entrambe le
47
direzioni
2 NO2 Cl
2NO2 + Cl2
Stadio lento
QUANDO UNA REAZIONE
AVVIENE A PIU’ STADI LO
STADIO PIU’ LENTO
DETERMINA LA
VELOCITA’ DI REAZIONE
2 NO2 Cl
NO2 Cl + Cl
48
NO2 + Cl
NO2 + Cl2
Velocità = K [NO2Cl]
* Le reazioni a molte tappe hanno una tappa
che determina la velocità
49
*
50
Stato di transizione
COMPLESSO ATTIVATO
A2 + B2
2 AB
Si rompono e
si formano
nuovi legami
51
•Si
combinano
transientemente col substrato
e ne abbassano l’energia di
attivazione
•Non spostano l ’ equilibrio
della
reazione,
ma
aumentano la velocità con
cui
l ’ equilibrio viene
raggiunto
•Non subiscono modificazioni permanenti durante la
reazione e sono subito disponibili per catalizzare un nuovo
52
ciclo di reazione
53
Gli enzimi sono biocatalizzatori che possono accelerare la
velocità delle reazioni chimiche da 106 a 1017 volte.
Gli Enzimi sono specifici sia per il tipo di reazione
catalizzata che per la scelta dei reagenti, chiamati
substrati.
Normalmente un enzima catalizza solo un tipo di reazione
chimica o una serie di reazioni strettamente correlate.
Rispetto ai catalizzatori chimici vi sono condizioni di
reazione più moderate.
Nella maggior parte delle reazioni enzimatiche non vi è la
formazione di sottoprodotti inutili come avviene spesso
nelle reazioni non catalizzate.
Gli enzimi hanno capacità di regolazione della loro
attività.
54