Organografia ed Anatomia
del fusto
1. Sostenere e distribuire
nello spazio le parti
aeree
2. Collegare tra loro foglie
e radici: trasporto
acropeto e basipeto
3. Organo di riserva
4. Partecipare alla fotosintesi
5. In molti casi funge da organo
perennante
(rizomi,bulbi,tuberi)
Fasci caulinari e fogliari
Hanno funzione di trasporto
I fusti rizomatosi hanno
Funzione di riserva
Modificazioni del fusto
Viticci
Spine
Rizomi
Bulbi
Fusti succulenti
Es. Echinocactus
Crescita e
differenziamento del
fusto
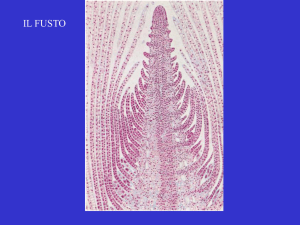
Il fusto si origina dall’apice
caulinare, i rami dagli apici delle
gemme ascellari
Tessuti primari del fusto
Tessuto tegumentale: Epidermide.
Cellule vive, con parete modificata, anche in funzione delle
condizioni ambientali, prive di spazi intercellulari. Svolge la
funzione di protezione e regola i rapporti della pianta con
l’ambiente esterno. Tra le sue funzioni, quindi, rientrano quello
di limitare la perdita d’acqua, protezione dall’eccessivo calore e
di opporre resistenza all’attacco dei patogeni.
Le pareti esterne dell’epidermide presentano depositi di cutina,
una sostanza idrofoba formata da acidi grassi a lunga catena.
La cutina riduce fortemente l’evaporazione. In piante che
vivono in ambienti non umidi esternamente alla cutina può
esserci deposizione di uno strato di cuticola al quale possono
aggiungersi anche delle cere. La cutina e le cere difendono dai
patogeni.
Tessuti tegumentali esterni: epidermide caulinare e
fogliare, rizoderma
I parenchimi
Le cellule parenchimatiche hanno una parete primaria sottile,
rappresentano il tipo più comune di tessuto e caratterizzano
tutte le parti molli di una pianta. Rappresentano il TESSUTO
DI RIEMPIMENTO.
Sono cellule metabolicamente attive e in genere rimangono vive
dopo che hanno completato il loro differenziamento.
Esistono numerosi tipi di parenchimi, specializzati per svolgere
particolari funzioni.
Parenchima di assimilazione o fotosintetico, con cellule
specializzate a svolgere la fotosintesi.
Parenchima di riserva, con cellule specializzate per l’accumulo
di sostanze di riserva, sono localizzate in varie parti del fusto –
parenchima amilifero le cui cellule sono ricche di amiloplasti in
cui viene condensato l’amido secondario o di riserva.
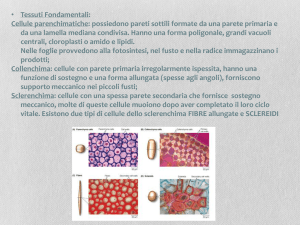
I tessuti meccanici
Svolgono la funzione di sostegno e
COLLENCHIMA e SCLERENCHIMA.
sono
distinti
in
COLLENCHIMA:
Le cellule del collenchima sono vive ed hanno una parete
primaria che si ispessisce in modo particolare, o solo agli angoli
(collenchima angolare) o solo sulle pareti tangenziali esterne ed
interne (collenchima tangenziale o lamellare) o presenta spazi
intercellulari e si ispessisce solo le porzioni di pareti che
affacciano su questi spazi (collenchima lacunare).
Il collenchima è presente negli apici lunghi e flessibili dei
germogli e può differenziare subito sotto l’epidermide o in
prossimità dei fasci conduttori.
COLLENCHIMA ANGOLARE
SCLERENCHIMA
Le cellule dello sclerenchima hanno pareti lignificate e, a,
differenziamento concluso sono per lo più morte. Le pareti
lignificate conferiscono elasticità alla cellula e infatti possono
essere deformate ma tornano alla loro forma originaria.
Le cellule sclerenchimatiche si differenziano negli organi che
hanno completato lo sviluppo ed hanno raggiunto le
dimensioni definitive, la sua rigidità sarebbe poco adatta ad
assecondare la crescita per distensione ad es. in un apice in
crescita.
Le cellule dello sclerenchima sono di due tipi: le fibre e le
sclereidi. Entrambe con pareti secondarie ispessite e
lignificate.
Le fibre sono lunghe e flessibili e si trovano in zone dove sono
necessari sostegno e flessibilità.
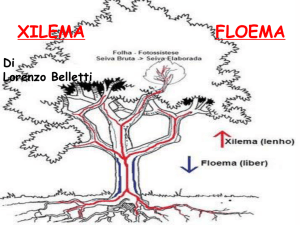
I tessuti conduttori
Nelle piante sono presenti due tipi di sistemi conduttori:
xilema o legno e floema o libro.
XILEMA
Lo xilema è formato da due diversi tipi di cellule: le tracheidi e le
trachee; entrambe hanno parete ispessita e lignificata. In entrambi i
tipi cellulari, detti complessivamente ELEMENTI TRACHEALI, dopo
la lignificazione della parete il protoplasto muore e tutta la cellula si
riduce alla solo parete. Allo xilema sono associate anche cellule
parenchimatiche e fibre.
La parete secondaria è impermeabile all’acqua, quindi alcune zone
della parete primaria devono rimanere prive di parete secondaria.
La deposizione della parete secondaria può avvenire secondo
diversi modelli.
L’area della parete in cui manca la deposizione della parete
secondaria è detta punteggiatura. Gli elementi tracheali più
resistenti sono quelli con punteggiature circolari areolate, in
queste punteggiature quasi tutta la parete primaria è ricoperta
dalla secondaria
Le punteggiature areolate possono essere abbastanza ampie da
consentire un flusso idrico rapido tra gli elementi tracheali
adiacenti.
Le punteggiature delle pareti trasversali, oblique, sono molto
ampie.
Nelle trachee il movimento dell’acqua avviene con minor attrito
che nelle tracheidi.
Nelle trachee manca l’intera regione di parete primaria e
secondaria trasversale, perché alla fine del differenziamento
questa viene digerita e si forma una perforazione, di
conseguenza ogni elemento tracheale ha due perforazioni
complete, una ad ogni estremità.
Floema
Il floema è formato da: cellule cribrose e tubi cribrosi
(elementi cribrosi).
Anche il floema è spesso associato a cellule parenchimatiche
e fibre.
Gli elementi cribrosi differentemente da quelli tracheali
devono rimanere vivi per svolgere la loro funzione.
Quando gli elementi cribrosi incominciano a differenziare i
loro plasmodesmi si allargano formando i pori cribrosi.
Questi sono raggruppati nelle aree cribrose.
La cellula cribrosa ed il vaso cribroso differiscono per forma e
nella disposizione delle aree cribrose. Se la cellula è
allungata,
affusola alle estremità ed ha aree cribrose
identiche su tutta la superficie è una cellula cribrosa. Questo
tipo si è evoluta per primo ed è presente nelle piante non
angiospermiche.
Nelle regioni internodali la stele presenta il tessuto vascolare ,
xilema e floema, organizzato in FASCI di conduzione
eventualmente intervallati da tessuti parenchimatici (RAGGI
MIDOLLARI).
A seconda del tipo di stele, al centro del cilindro vascolare il
tessuto parenchimatico può estendersi in un’area omogenea
detta MIDOLLO.
Ai nodi, la struttura stelare viene interrotta dalla presenza delle
lacune fogliari o rameali. I fasci divergono dal cilindro centrale
e, attraversata la corteccia vanno ad innervare la foglia o il
ramo.
La pre-protostele delle Briofite (formata da cellule parenchimatiche
o meccaniche allungate) precorre la stele delle cormofite.
Il fusto delle crittogame
vascolari
Esempio di protostele di Psilotum
(crittogame vascolari,
generazine sporofitica)
Fusto primario di Dicotiledoni e
Gimnosperme
Fusto di dicotiledone erbacea - eustele
Esempio:
Sezione trasversale di fusto di Cucurbita in
struttura primaria.
Dall'esterno all'interno si notano:
l'epidermide, il collenchima angolare, il
parenchima corticale, una guaina
sclerenchimatica e i fasci immersi nel parenchima
midollare.
In particolare, tali fasci sono di tipo bicollaterale,
cioè con floema sia all'interno che all'esterno dello
xilema e con cambio fra di essi interposto.
Solo il cambio tra xilema e floema esterno è attivo
Fusto di Monocotiledone
Es.
Altro es.
Tipico fascio collaterale
chiuso con lo xilema a
forma di V.
Fascio concentrico
perixilematico nel rizoma di
Acorus calamus
Da notare, inoltre, la
presenza di un parenchima
aerifero (che ha anche
funzioni di riserva) che rivela
chiaramente il carattere
idrofita di questa pianta.
Fusto di monocotiledoneATACTOSTELE
Altro es.
Sezione trasversale a
livello dell'internodo
del fusto di Secale
sp. (Monocotiledoni).
Esempio di fusto
cavo.
Da notare, a livello
sottoepidermico, le
isole di parenchima
clorofilliano tra le
aree
sclerenchimatiche
Altro es.:
Fusto di Zea mays
Altro esempio di struttura
atactostelica, con i fasci cribrovascolari che si spingono a
ridosso dell'epidermide.
La transizione a struttura
secondaria nel fusto degli
alberi (angiosperme
dicotiledoni e gimnosperme
Le piante legnose non solo diventano più
alte in seguito all’attività dei meristemi
apicali, ma si accrescono anche in spessore
grazie alla deposizione annuale di legno, di
libro e di sughero. Poiché il legno ed il libro
contengono tessuti di conduzione, il loro
aumento garantisce alla pianta una
maggiore capacità di trasporto dell’acqua e
dei sali. Di conseguenza la pianta può
formare un maggior numero di foglie e di
radici ed aumentare il suo tasso
fotosintetico.
Va sottolineato che una pianta legnosa è
formata dalla combinazione di tessuti
primari e secondari. Le regioni terminali del
fusto e delle radici così come le foglie, i fiori
ed i frutti fanno parte della struttura
primaria. La struttura secondaria si realizza
nei fusti e nelle radici.
Attualmente
un
vero
accrescimento
secondario è presente in tutte le
Gimnosperme ed in molte Dicotiledoni, è
invece assente nelle Monocotiledoni e nelle
Felci.
I meristemi primari e secondari
Accrescimento in altezza
dovuto ai M I apicali caulinari
Accrescimento in spessore
dovuto ai M II cambiali
Accrescimento in altezza
dovuto ai M I apicali radicali
>Il passaggio dalla struttura primaria
alla
secondaria
non
avviene
contemporaneamente per la zona
interna e per l’esterna del fusto (di
solito si formano prima xilema e
floema II.
>Lo
sviluppo
della
struttura
secondaria porta alla distruzione di
gran parte dei tessuti primari del
fusto.
L’attività di divisione delle cellule
del cambio
• Divisioni periclinali diseguali creano
xilema e floema secondari
• Divisioni uguali anticlinali generano
nuove cellule di cambio
• Divisioni trasversali uguali generano
parenchima dei raggi midollari
La struttura del legno delle angiosperme è molto
più varia. Per la presenza di diversi tipi cellulari.
Trachee, tracheidi, fibre e cellule
parenchimatiche.
La presenza delle tracheee distingue i legni delle
gimnosperme da quelli delleangiosperme.
I raggi midollari del legno delle angiosperme sono
molto più grandi di quelli delle gimnosperme.
Il legno delle Angiosperme è ETEROXILO.
Sez
Longi
tudinale
Tangen
ziale
Sezione trasversale
raggio
Legno eteroxilo ed omoxilo
Libro II°
Il floema o libro secondario presenta un sistema
assiale ed uno radiale come il legno. Il sistema assiale
provvede alla conduzione della linfa elaborata in
senso verticale.
Nelle ginmosperme è costituito da sole cellule
cribrose mentre nelle dicotiledoni sono presenti tubi
cribrosi e cellule compagne.
In entrambi i gruppi di piante è presente
parenchima e fibre floematiche.
Nel floema secondario non si osservano andamenti
stagionali come per lo xilema.
Il floema secondario è attivo per meno di un anno.
Solo lo strato più interno del floema è attivo nella
conduzione.
I raggi presenti nel floema hanno caratteri
morfologici simili a quelli dello xilema, sono
entrambi prodotti dalle stesse iniziali dei
raggi.
I raggi sono formati solo da cellule
parenchimatiche con funzione di riserva, i
raggi floematici sono però più efficienti.
L’attività periodica del cambio cribro-vascolare, nelle regioni
temperate è legato alle stagioni e produce aumenti dovuti alla
crescita, anelli di crescita.
Se uno strato di crescita rappresenta l’attività di una stagione si
chiama cerchia annuale.
Durante i periodi freddi o estremamente secchi il cambio va in
quiescenza. In primavera, cessata la quiescenza, si riattiva e
riprende a dividersi e contemporaneamente la pianta produce
nuove foglie. Queste ultime hanno l’epidermide non ancora
cutinizzata e quindi perdono molta acqua di conseguenza il
primo legno che si forma, detto legno precoce o primaverile per
essere efficiente deve avere un’elevata percentuale di vasi ampi.
Nelle gimnosperme questo si traduce nella formazione di
tracheidi con diametro maggiore.Successivamente quando le
foglie si sono adattate a limitare la perdita d’acqua per evitare
un flusso troppo veloce vengono prodotti vasi con lume più
piccolo e in minor quantità. Questo legno è detto legno tardivo o
In tutti i fusti e le radici legnose dopo l’inizio
della produzione di xilema e floema secondari
si forma il PERIDERMA.
Sostituisce l’epidermide.
È costituito da tre parti:
Cambio del sughero o fellogeno
Sughero o fellema
Felloderma
Periderma
si origina per attività del fellogeno.
Strato di cellule compreso fra le porzioni più
esterne della corteccia I o del libro e l’
epidermide (o l’esoderma nella radice)
prodotto dal fellogeno che produce sughero
all’ esterno e felloderma all’ interno
Cambio
suberofellodermico
Passaggio Ia
- IIa
Nel fusto della maggior parte delle piante legnose il primo
fellogeno si forma durante il primo anno di crescita e
principalmente dalle cellule corticali poste subito sotto
l’epidermide, in qualche caso anche l’epidermide è coinvolta.
In qualche pianta può formarsi più in profondità in genere
dal floema primario.
L’attività del cambio del sughero porta alla formazione di
strati radiali di cellule disposte in modo compatto la maggior
parte sono cellule di sughero.
Le pareti delle cellule del sughero sono ricoperte da lamelle di
suberina, cioè strati di suberina alternati a strati di cere che
rendono estremamente impermeabile la parete. Queste pareti
possono anche lignificare.
A maturità le cellule del sughero sono morte.
Scorza o ritidoma:
nella maggioranza delle piante arboree,
rappresenta
l’ insieme dei tessuti che vanno dal sughero più
interno fino all’ esterno della pianta
Le cellule del felloderma a differenza di quelle del sughero
sono vive, mancano di lamelle di suberina e somigliano a
cellule prenchimatiche.
Passaggio dalla struttura primaria a quella
secondaria
> Formazione di un anello cambiale continuo
> Le cellule cambiali danno origine : mediante divisioni longitudinali
tangenziali, a xilema e floema secondari; per divisione longitudinale radiale
ad altre cellule di cambio, e per divisione trasversali a parenchima
midollare secondario;
> Il libro primario viene spinto verso la periferia del fusto, il legno primario
viene compresso verso il centro.
> Il fellogeno genera sughero sul lato esterno e felloderma su quello interno
(l’insieme dei tre tessuti costituisce il periderma). Le lenticelle sono zone
del periderma ricche (a differenza del sughero) di spazi intercellulari. A
differenza del cambio che vive quanto il fusto (nelle piante arboree), il
fellogeno dura, generalmente un solo anno. Come conseguenza sempre
nuovi peridermi si originano sempre più internamente. Tutti i tessuti
esterni al fellogeno più interno, costituiscono la corteccia esterna o
RITIDOMA. Questa è completamente costituita da tessuti morti. La parte
viva della cortecciache si trova all’interno del fellogeno attivo è detta
corteccia interna.
Elementi vivi di un fusto
secondario
Cellule del fellogeno e del felloderma
Floema II (spessore massimo 1 mm)
Zona cambiale (estremamente sottile)
Cellule parenchimatiche e dei raggi midollari
(associate al legno) negli anelli di crescita più
periferici
I cambi vascolari ed i tessuti
secondari della radice e del fusto si
continuano gli uni con gli altri.