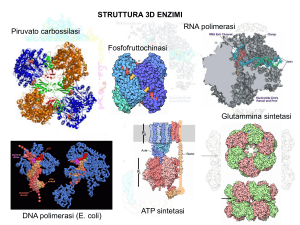
DETERMINAZIONE DEI PARAMETRI CINETICI
DEGLI ENZIMI - Vmax E Km
Saggio enzimatico in saturazione di S: V0 = K2 [ET]
permette di ottenere le U di attività catalitica relative ad un
enzima in un campione biologico: U.E. dipendenti direttamente
dalla concentrazione di E totale presente;
Determinazione dei parametri cinetici di un enzima: Vmax e Km:
fondamentale in biochimica applicata – Km: indice di affinità dell’enzima per il suo
substrato; Vmax: massima velocità alla [Et] data;
studio di inibitori e regolatori enzimatici; identificazione di isoenzimi;
consente di «seguire» le tappe di purificazione di un enzima.
Le applicazioni dello studio degli enzimi, oltre ad essere basilari nella ricerca
fondamentale biochimica, hanno importanti ricadute in diagnostica clinica e
in farmacologia.
ENZIMI
Gli enzimi sono i biocatalizzatori delle reazioni chimiche all’interno delle cellule.
Sono lo «strumento» principe attraverso il quale si realizzano:
metabolismo energetico, biotrasformazioni o processi fondamentali della vita
cellulare quali duplicazione del DNA, trascrizione dell’RNA e sintesi proteica.
Sono macromolecole biologiche, proteine.
Eccezione: i ribozimi, particolari RNA con attività catalitica nelle cellule.
Ipotesi interessante: RNA, prime molecole con funzione di catalizzatori biologici,
sostituite dalle proteine, strutturalmente più adatte ad esplicare questa funzione:
RNA
PROTEINE
Enzimi proteici:
proteine a singola catena polipeptidica;
proteine con struttura quaternaria;
isoenzimi (ex. LDH) con diversa localizzazione nei tessuti
o a livello sub-cellulare
Parte proteica apoenzima + parte non proteica
oloenzima
Parte non proteica: Gruppi prostetici: ex: eme nel Citocromo P450; Coenzimi,
Cofattori inorganici: metalli, (Zn, Mg, Cu, Fe..) in metalloenzimi
PROPRIETA’ DEGLI ENZIMI
VELOCITÀ DELLE REAZIONI: enzimi aumentano la velocità di reazione fino a
108-1012 volte la velocità in assenza del catalizzatore. Reagente/i = substrato/i.
SPECIFICITA’: Specificità degli enzimi è un criterio di classificazione:
enzimi a specificità verso il substrato assoluta (una molecola unica); di tipo
stereochimico (riconoscimento degli stereoisomeri); di gruppo (gruppo chimico;
ex. metiltransferasi); di legame (un legame chimico, ex. proteasi, legame
peptidico).
REGOLAZIONE: gli enzimi sono soggetti a regolazione per adattare l’attività
catalitica alle necessità fisiologiche e ai cambiamenti che si attuano nelle cellule.
inibizione a feed-back, inibizione da substrato o prodotto, fosforilazione reversibile,
induzione e regolazione dell’espressione genica.
Siti di regolazione - modulazione allosterica: siti di legame con molecole
che modulano l’attività enzimatica. Struttura quaternaria e reg. allosterica.
DOMINI E PORZIONI DELLE PROTEINE CON FUNZIONE DI ENZIMA:
Sito attivo o catalitico: “tasca” catalitica, dominio dell’enzima che riconosce il
substrato e si lega reversibilmente con questo, formando ES. Nel sito catalitico si
trovano alcuni residui di AA che sono coinvolti nel meccanismo di catalisi grazie
alla natura chimica dei loro gruppi –R. Complementarietà e/o adattamento sterico
tra E e S. La conformazione 3D del sito attivo – struttura 3D di tutta la molecola.
MECCANISMO DI INTERAZIONE ENZIMA-SUBSTRATO
CONSIDERAZIONI ENERGETICHE E CINETICHE
Termodinamica chimica: E libera G di reazione,
G(P) –G(R) = G
G = H – TS
G < 0, reazione esoergonica, spontanea da sinistra vs. destra; G > 0, reazione endoergonica,
spontanea da destra vs. sinistra; G = 0, all’equilibrio nei due sensi.
G0’: variazione di E libera standard, 1 M, a pH 7, Kcal mole-1
A+B
C+D
reazioni reversibili
G = - RT ln Keq + RT ln ([C] [D]/[A] [B])
G0’ = - RT ln Keq , a pH 7
La spontaneità di una reazione chimica o della direzione di un equilibrio chimico
dai reagenti ai prodotti e viceversa non ci dice però niente sulla cinetica e la
velocità della reazione stessa.
E di attivazione: soglia energetica di una reazione chimica;
Coordinate di reazione: percorsi e distanza delle specie chimiche in gioco ;
Equazione di Arrhenius : k = A e-Ea/RT
K = cost. di velocità della reazione
o vel. specifica di reazione
A= fattore di frequenza, geometria
delle molecole;
R= costante universale dei gas
T= temperatura in Kelvin
Teoria dello stato di transizione
Tanto minore è E att., tanto più la reazione raggiunge facilmente lo stato
di transizione e può procedere più velocemente nel verso termodinamicamente
favorito.
Quindi: una reazione può procedere o avvenire + veloc. non per il G tra R e P ma per
il valore di Eatt. Anche nella cellula valgono tali principi - le reazioni intracellulari –
isobare e isoterme – anche se hanno un G neg. procederebbero comunque troppo
lentamente; le reaz reversibili non potrebbero andare in una direzione o in un’altra
prontamente per rispondere alle esigenze cellulari: gli enzimi svolgono questa funzione.
E(att.) di una reazione
chimica con e senza l’enzima
Un enzima:
-accelera la reazione;
-non è né un R né un P;
-non viene consumato durante la reaz.;
-non sposta l’equilibrio di reaz.;
-non modifica il G complessivo R vs. P
-è presente in quantità minime,
non stechiometriche.
CINETICA ENZIMATICA
Le reazioni del metabolismo energetico possono essere spontanee o meno e, se
sono reversibili, avvengono in un verso o l’altro in base alle concentrazioni dei
reagenti o dei prodotti. Gli enzimi abbassano l’Eatt. delle reazioni senza modificare la
direzione della reazione, la necessità o meno di consumo energetico e gli equilibri.
Dalla cinetica chimica:
k1
E+S
K-1
K2 =Kcat.
ES
K-1
v0
S
k1
E+P
K-2
P
Vf = k1 [R], senza enzima
Eq. Henri-Brown
Cinetica in stato stazionario: si vede
come varia la vel. enzimatica a varie [S];
approccio semplificato per studiare la
cinetica delle reazioni catalizzate da E;
assunzioni teoriche:
- [E] << [S]
- velocità iniziale, V0:
- Stato stazionario:
[ES] = k
K1[E] [S] = k-1 [ES]
Alle condiz. fissate di:
temperatura, pH, forza ionica,
alla v0, la pendenza della tangente
alla curva [S] dt dip. da [ET ].
Iperbole di Michaelis & Menten
Vmax= K2[Et]
V0 = k2 [ET] [S]
Km + [S]
La V0 dipende da: [E], [S] e Km
b) V0 = K2 [ET]
1) Per [S] << Km
la V0 dipende da [S] e [Et]:
V0 = (Vmax/ [S])/Km
Reazione che segue una
cinetica del 1° ordine;
a)
V0 = k2[ET] [S]
Km
- 2) Per [S] = Km , V0 = Vmax/2
Km = K-1 + K2/ K+1
Situazioni limite:
a) [S] molto basse,
Lim [S]
0
V0 = k2[ET]/2
Reazione che dipende da [S] = Km
dell’ E in gioco e da [ET]
- 3) Per [S] >> Km, V0 = Vmax,
reazione di ordine cinetico = 0
La V0 dipende solo da [ET]
V0= K
b) [S] verso la
saturazione
CINETICA DEL 1° ORDINE
Situazione limite a)
1° tratto della M & M
Esempi generici
- dS/dt = K [S], retta,
Quando S << Km
[S] scompare nel tempo
secondo una curva esponenziale:
integrando - dS/dt= k [S]:
[A] = A0e-kt
[A]
-∫ A
0
d[A]/[A] = ∫
t
kdt
0
CINETICA DI ORDINE 0
Situazione limite b)
in saturazione
[A]
[A]=A0 -kt
dS/dt= K
[S] scompare nel tempo secondo
una retta con pendenza –k
integrando - dS/dt= k
[A]
tempo
-∫ A
0
d[A] =
t
∫0
kdt
Linearizzazione iperbole di Michaelis- Menten
L’iperbole si può linearizzare, rendendo l’analisi dell’attività E più immediata
e di più facile gestione
pendenza
Altre linearizzazioni: Hanes-Woolf; Eadie-Hofstee
Vmax e Km ADENOSINA DEAMINASI
+ H2O
+ NH3
idrolasi
Adenosina
Inosina
Come si realizza in vitro il saggio di
determinazione dei parametri cinetici di questa
trasformazione
Esistono diverse metodiche per determinare la velocità enzimatica di questa
deaminasi – saggio spettrofotometrico: Abs 260/265 nm.
A 260nm si misura l’Abs dell’adenosina; la velocità V0 dell’adenosina deaminasi,
ovvero l’ estinzione a tempo iniziale dell’adenosina per effetto della trasformazione
enzimatica in inosina, si misura invece a 265nm.
Questo viene fatto per eliminare l’interferenza del prodotto,
l’inosina, che assorbe a 260 ma non a 265 nm.
ESPERIMENTO: Vmax e Km dell’enzima Adenosina Deaminasi con metodo
spettrofotometrico UV
Si determina la V0 a varie concentrazioni di Substrato (adenosina), scelte al di
sotto e al di sopra della Km attesa (~40 mM): le concentrazioni di adenosina sono
comprese tra 5 e 80 mM.
S1: 5 mM
Soluzione stock
di partenza
Adenosina 300 mM
diluizioni
S2: 10 mM
S3: 20 mM
S4: 40 mM
S5: 60 mM
S6: 80 mM
a) Verifica concentrazione delle soluzioni preparate di adenosina:
Spettrofotometro: si legge a 260 nm (UV) per verificare [Adenosina] mediante la
e, coeff. di estinzione mmolare dell’adenosina = 0.015. A260nm/0.015= conc mM
b) Calcolo della V0 dell’adenosina deaminasi:
Si sposta la l di lettura da 260 a 265 nm e si aggiunge l’enzima Adenosina
Deaminasi (opportunamente diluito) alle soluzioni di adenosina preparate :
+ Adenosina Deaminasi
Reazione:
Adenosina
Inosina
In sostanza: per ogni soluzione di adenosina, si calcola la V0: -dS/dt, A/min, in 2 min
d’intervallo di tempo: lo strumento misura l’Abs a 265nm ogni 10 sec in 2 min.
A nel tratto lineare: A(iniziale) – A(finale) sulla retta tangente alla curva di estinzione
di A in funzione di t: si rapporta al min (A/min). Diluendo opportunamente l’E, il tratto sarà lineare nei 2 min.