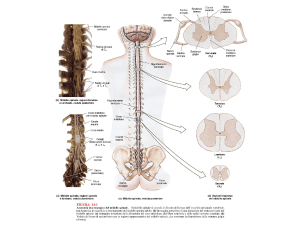
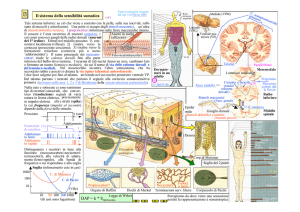
Prima di iniziare a vedere le caratteristiche di questo
sistema facciamo un cenno alla classificazione delle
fibre nervose. Questa che vediamo vale sia per le fibre
sensitive, sia per le fibre motorie. Le fibre sono
classificate in due gruppi principali A e C con
sottogruppi.
Fibre
A
C
Tipo
Proprietà
isolanti
Diametro
Velocità di
conduzione
A
mieliniche
13-20 m
80-120 m/s
A
mieliniche
6-12 m
35-75 m/s
A
mieliniche
1-5 m
5-30 m/s
amieliniche
0.2-1.5 m
0.5-2 m/s
Il sistema somato-sensoriale fornisce informazioni
riguardanti molteplici modalità, con recettori di diverso
tipo che segnalano sensazioni quali tatto, pressione,
temperatura e dolore. I recettori di questo sistema si
trovano distribuiti in tutte le parti del corpo e all’interno
di esso. Il sistema somato-sensoriale vedremo che
utilizza due vie nervose ascendenti per processare
l’informazione a livello del cervello. Tali vie
forniscono indicazioni che riguardano il tipo di
stimolo e la sua localizzazione a livello corporeo.
Le sensibilità somatiche si possono suddividere in
tre differenti tipi:
Sensibilità somatiche meccanocettive: sollecitazioni
meccaniche dei tessuti corporei (tatto, pressione,
solletico, vibrazione e senso di posizione)
Sensibilità dolorifiche: evocata da qualunque fattore
che danneggi i tessuti
Sensibilità termiche: per il caldo e per il freddo
Il tatto, la sensazione di pressione e quella vibratoria
sono sensazioni distinte che dipendono dallo stesso
tipo di recettori con queste differenze:
Il tatto dipende in genere da recettori della cute o
immediatamente sottostanti
La pressione è percepita da recettori più profondi
La vibrazione utilizza i due recettori appena detti,
specie quelli a rapido adattamento, ma in rapide
successioni.
Recettori tattili
Si conoscono almeno 6 tipi del tutto differenti di
recettori tattili. Ricordiamo:
1. Terminazioni nervose libere: si trovano nella
cute e in altri tessuti. Rilevano stimoli tattili
superficiali e profondi. Sono fibre A (5-30m/s)
2. Corpuscoli di Meissner: terminazione nervosa
incapsulata in rapporto con una grossa fibra
mielinica (A 30-70m/s). Tipici della cute glabra
e specie di labbra e polpastrelli. Sono fasici.
3. Dischi di Merkel: tipici delle regioni cutanee
sensibili, anche in zone provviste di peli. Sono
tonici. Permettono di avvertire il contatto continuo
di oggetti sulla cute. In rapporto con una fibra A.
4. Organo terminale pilifero: pelo + fibra nervosa
basale. Sono recettori fasici. Segnalano oggetti in
movimento sulla superficie corporea.
5. Organi terminali di Ruffini: terminazioni
incapsulate e pluriramificate. Sono recettori a
medio adattamento della cute profonda.
6. Corpuscoli del Pacini: tipici di cute e tessuti
profondi. Segnalano pressione e vibrazioni.
Sono a rapidissimo adattamento. Li vediamo
meglio.
cute glabra
fasici
Aδ
cute profonda
fasici
fasici
tonici
Aβ
Cute
con peli
Il corpuscolo del Pacini è un particolare neurone-
recettore (neuroni a T o pseudounipolari) con una
caratteristica struttura periferica per la captazione dello
stimolo. È un recettore fasico che presenta una parte
recettrice altamente specializzata e una parte assonica
con un rivestimento di mielina. La parte periferica è
formata da lamelle concentriche di connettivo separate
fra loro da un gel viscoso. Se sperimentalmente la parte
periferica è eliminata lasciando la fibra nuda, il recettore
diviene tonico.
Il potenziale generatore insorge perché lo stimolo pressorio
determina una deformazione della struttura a lamelle
concentriche facendo aprire canali del Na+ che
depolarizzano la membrana.
Sequenza di eventi
dall’insorgenza alla
scomparsa di uno
stimolo pressorio sul
corpuscolo del Pacini
Quasi tutte le informazioni della sensibilità somatica
entrano nel midollo spinale attraverso i gangli delle
radici posteriori (dorsali, DRG neurons) dei nervi
spinali. Dopo l’ingresso nel midollo e fino all’encefalo
questi segnali seguono due vie distinte: il sistema
dorsale o del lemnisco mediale e la via antero-laterale.
Questi due sistemi convergono nel talamo.
central
process
cell body
in DRG
peripheral
process
DRG (dorsal root ganglion)
neuron
pseudounipolar cell
Nel sistema somato-sensoriale, le vie sensoriali sono
formate da tre neuroni:
Neurone del primo ordine: i corpi cellulari di questi
neuroni sono localizzati nei gangli delle radici dorsali
del MS. La parte distale di questi neuroni è altamente
differenziata come abbiamo visto nei corpuscoli di
Pacini, di Merkel, etc….La parte prossimale entra nel
MS.
Neuroni del secondo ordine: questi neuroni sono in
contatto con i neuroni del I ordine e hanno il soma a
livello del MS (via antero-laterale) o del tronco
encefalico (via del lemnisco mediale). Le fibre dei
neuroni del II ordine in genere decussano, proiettando
quindi controlateralmente e poi ascendono a livello del
talamo dove entrano in contatto con il neurone del III
ordine.
Neurone del terzo ordine: proietta da varie aree del
talamo (nuclei sensoriali talamici) alla corteccia
sensoriale cerebrale. La stazione talamica esiste in
tutti i sistemi fatta eccezione per parte delle fibre del
sistema olfattivo in cui la stazione talamica è saltata e
le fibre proiettano direttamente in coteccia olfattiva.
Sistema dorsale o del lemnisco mediale: trasporta
segnali che seguono la via dei cordoni dorsali del MS
e più in alto quella del lemnisco mediale fino al talamo.
Fibre nervose a 30-110m/s. La conduzione lungo
questo sistema consente una fine percezione di forma,
peso e tessitura di un oggetto. La capacitá di
identificare un oggetto attraverso la sua forma è detta
STEREOGNOSI
Vie motorie
Vie sensoriali
3. Sistema colonne dorsali
3a fasciculus gracilis
3b fasciculus cuneatus
Sacrale
Lombare
Toracico
Cervicale
4. Tratto spino-cerebellare
4a posteriore
4b anteriore
5. Sistema antero-laterale
5a spinotalamico laterale
5b spinotalamico anteriore
6. Tratto spino-olivare
Il sistema del lemnisco mediale ha un grado molto
elevato di organizzazione spaziale delle fibre nervose
(organizzazione SOMATOTOPICA), mentre il sistema
antero-laterale ha un grado di organizzazione molto
inferiore. L’informazione che richiede di essere
trasmessa rapidamente e con alta fedeltà spaziale e
temporale prende la via del lemnisco mediale, mentre
per informazioni “più grezze” è imboccata la via
antero-laterale.
Via del lemnisco mediale
Decussazione
N.B.: esistono due divisioni principali delle colonne
dorsali, il fasciculus gracilis e il fasciculus cuneatus. Le
fibre sensitive entrano nelle colonne dorsali
ipsilateralmente. Alcune fibre danno luogo a riflessi
locali. Le rimanenti ascendono nei due tratti – il
fasciculus gracilis (fibre provenienti dalle estremità
inferiori sotto T7) e il fasciculus cuneatus (fibre
provenienti dalle estremità superiori sopra T7). I
neuroni del I ordine sinaptano con quelli del II nel
nucleo gracilis e cuneatus del bulbo.
Gli assoni
dei neuroni
del II ordine
si portano
al nucleo
ventrale
posterolaterale del
talamo dove contraggono sinapsi con i neuroni del
III ordine.
Antero-laterale
Lemnisco-mediale
Decussazione
Colonne
dorsali di
sostanza
bianca nel
MS
Soma del neurone
del I ordine
Sistema antero-laterale (tratto spino-talamico):
riceve segnali da nocicettori, meccanocettori,
termocettori. Le fibre entrano nel MS attraverso i
gangli della radici dorsali e ascendono per qualche
tratto (tratto di Lissauer); sinaptano con i neuroni del II
ordine nel MS, decussano a livello di MS e ascendono
nei cordoni anteriore e laterale della sostanza bianca
controlaterale per terminare nel tronco encefalico
(bulbo) e nel talamo. Fibre nervose a 40m/s.
Via antero-laterale
Sistema del lemnisco mediale
a.
Sistema antero-laterale
Sensibilità tattile altamente
a.
Sensibilità dolorifica
discriminata per la
b.
Sensibilità termica
localizzazione dello stimolo
c.
Sensibilità tattile
e per l’intensità
grossolana di pressione
b.
Sensibilità fasiche
d.
Sensazione di solletico
c.
Senso di posizione
e.
Sensazioni sessuali
d.
Senso di pressione
e.
Movimento sulla cute
Sistema del lemnisco mediale
Sistema antero-laterale
Le fibre nervose che servono particolari aree cutanee
si riuniscono insieme a formare i nervi e l’area servita
dai differenti nervi cutanei è mappabile. Ciascuna
area è la combinazione dei campi recettivi di tutte le
fibre che formano il nervo, cioè il campo recettivo del
nervo periferico. Nonostante alcune variazioni
individuali, i campi recettivi dei nervi sono abbastanza
costanti in localizzazione ed estensione da essere
usati per anamnesi clinica. Quando un nervo è
resecato, questo risulta in un’area di completa
assenza di sensazione, cioé in anestesia.
Anche i campi recettivi
delle fibre che
raggiungono una
radice dosale, coprono
un’area ben delimitata
che prende il nome di
dermatomero. Questa
organizzazione
somatotopica è una
caratteristica costante
del sistema nervoso
I segnali provenienti dalla periferia devono poi essere
integrati a livello di SNC per elaborare una risposta. Il
termine INTEGRAZIONE si riferisce al modo in cui il
sistema nervoso interpreta l’informazione sensoriale e
determina la risposta appropriata ad uno stimolo.
Alcune forme di integrazione hanno luogo a livello di
midollo spinale, per dare origine ai cosiddetti archi
riflessi. Altri segnali sono integrati a livello di tronco
spinale. L’integrazione piú complessa ha luogo a livello
di corteccia, dove una sensazione diviene coscienza e
attenzione.
Pain withdrawal reflex
Corteccia somestesica:
aree di Brodmann (inizi 1900)
La corteccia encefalica è
suddivisa in 50 aree corticali dette
aree di Brodmann suddivise sulla
base di differenze strutturali
istologiche rilevabili nelle colonne
di neuroni. Questa mappa si
utilizza come riferimento per le
diverse aree funzionali della
corteccia umana.
Solco centrale
Notare il solco centrale o scissura di Rolando che si estende
in direzione infero-superiore sulla faccia laterale di ciascun
emisfero. Separa il lobo frontale dal lobo parietale. In genere
le informazioni somatiche terminano nella corteccia
posteriore alla scissura di Rolando.
Lateral sulcus
Per quanto riguarda la corteccia somestesica è
localizzata subito dietro la scissura di Rolando e
principalmente nelle aree 1, 2, 3, 5, e 40 che
grossolanamente costituiscono il lobo parietale della
corteccia.
Le aree che si trovano anteriormente alla scissura di
Rolando sono coinvolte nella motilità somatica e in
alcuni aspetti del pensiero analitico.
Esistono due diverse aree alle quali giungono fibre
afferenti direttamente dai nuclei somestesici del
talamo: area somestesica I (area S-I) corrispondente
al giro post-centrale e area somestesica II (area S-II).
Nell’uomo l’area S-I comprende le aree 1, 2 e 3 di
Brodmann. Esiste in queste zone una netta
localizzazione spaziale delle zone deputate alla
ricezione di segnali nervosi provenienti dalle diverse
parti del corpo.
Esiste un classico schema che riporta le
rappresentazioni delle diverse parti del corpo nelle
differenti zone di S-I. Alcune zone hanno una
rappresentazione molto estesa, come le labbra, la
faccia, il pollice, mentre il tronco e tutto il corpo sono
rappresentati da aree relativamente piccole.
Si parla di homunculus sensitivus.
I neuroni di S-I sono disposti in colonne che
attraversano tutti gli strati della corteccia. Ciascuna
colonna è al servizio di un’unica modalità sensoriale e
le colonne sono frammiste le une alle altre.
Un’ampia escissione dell’area S-I causa:
Incapacità a localizzare le diverse sensazioni nelle
aree del corpo
Incapacità a discriminare diverse pressioni
Incapacità a valutare il peso degli oggetti
Incapacità a riconoscere al tatto tipo e forma degli
oggetti
Incapacità ad apprezzare la tessitura degli oggetti.
L’area S-II è meno estesa e comprende le aree 5 e 40.
Il grado di localizzazione delle diverse parti del corpo è
molto inferiore a S-I. L’area S-II riceve in ingresso
segnali provenienti dall’area S-I e da altre aree
sensoriali, funzionando come corteccia di
associazione. L’ablazione di S-II determina:
Incapacità a riconoscere al tatto oggetti di forma
complessa
L’ablazione monolaterale determina la perdita
della consapevolezza dello schema corporeo
controlaterale. Si parla di amorfosintesi.
La distruzione della corteccia somestesica nell’uomo è
seguita dalla perdita della sensibilità tattile più fine e
discriminativa , mentre si recupera un lieve grado di
sensibilità tattile grossolana. Il talamo quindi, seppure
in maniera lieve permette di avvertire sensazioni tattili.
La perdita della corteccia somestesica non ha invece
effetto sulla sensibilità al dolore e alla temperatura.
Dolore
Il dolore costituisce per l’organismo un meccanismo di
difesa. Esso insorge quando un tessuto sta subendo
un danno. Il dolore è classificato in due tipi:
Acuto: insorge entro 100ms dallo stimolo dolorifico.
Viene indicato anche come lancinante, pungente,
rapido, elettrico. Non è avvertito in genere nei tessuti
profondi. Fibre A
Lento: insorge dopo 1s o più. Viene anche detto
urente, pulsante, nauseante o cronico. Solitamente è
associato a distruzione di tessuti. Fibre C.
Confronto fra dolore acuto e dolore cronico: tratto spinotalamico
Dolore acuto
Dolore lento
Pungente, rapido
Tedioso, continuo
Fibre A
Fibre C amieliniche
Latenza breve
Insorgenza lenta
Ben localizzato
Diffuso
Breve durata
Lunga durata
Non sensibile alla morfina
Bloccato dalla morfina
Tratto neospinotalamico
Tratto paleospinotalamico
I recettori dolorifici sono tutti costituiti da terminazioni
nervose libere diffuse negli strati superficiali della
cute e anche nei tessuti profondi quali periostio,
pareti arteriose, superfici articolari.
Fra questi recettori ricordiamo:
nocicettori meccanici
nocicettori termosensibili
nocicettori chemosensibili (bradichinina, serotonina,
istamina, ioni K+, acidi, ACh ed enzimi proteolitici
sono sostanze stimolanti)
I recettori implicati nella trasduzione di segnali
dolorifici sono tutti a lentissimo o nullo adattamento e
questo è importante per continuare a segnalare la
presenza di uno stimolo doloroso. Si classificano in:
Recettori sensibili alla temperatura: al di sopra dei
43ºC il calore attiva canali del Na+ determinando una
depolarizzazione. Questi recettori sono attivati da
capsaicina, l’ingrediente fondamentale del chili
pepper.
Recettori pH-dipendenti: i canali responsabili della
depolarizzazione rispondono a pH inferiori a 6.5 in
seguito a danno tessutale. Sono identificati come
canali chemio-dipendenti .
Recettori sensibili all’ATP: le cellule danneggiate
rilasciano ATP che determina l’apertura o la chiusura
di canali determinando depolarizzazione e quindi un
segnale nocicettivo.
Recettori sensibili alla bradichinina: le chinine, inclusa
bradichinina e kallidina, sono peptidi prodotti a livello di
tessuti danneggiati. Derivano dall’azione degli enzimi
proteolitici rilasciati dalle cellule danneggiate su una
pre-proteina. Attivano recettori specifici B1 e B2 che
sono accoppiati ad un vasto numero di meccanismi di
trasduzione del segnale. Inducono vasodilatazione,
stimolano le cellule del sistema immunitario e stimolano
i recettori dolorifici.
Recettori sensibili al K+: il potassio é rilasciato da
cellule danneggiate e attiva i nocicettori limitrofi.
Esiste una via di trasmissione per il dolore acuto ed
una via per il dolore lento. I segnali del dolore rapido
utilizzano fibre afferenti di tipo A (6-30m/s), mentre
quelli del dolore lento sono trasmessi da fibre C
amieliniche (0.5-2m/s). Il dolore acuto ha in genere
una miglior localizzazione e può essere localizzato
esattamente se sono stimolati anche recettori tattili.
I neurone: DRG
II neurone: corna
ventrali del MS
(Lamina I, IV, V)
e prosecuzione
nel tratto spino-talamico
III neurone: Talamo (VPL)
Terminazione: (S I)
Tratto spinotalamico
decussazione
Commissura
bianca anteriore
Radice posteriore
Tratto spino-talamico
- In caso di lesione viene persa la
sensazione di dolore e di percezione
della temperatura nella parte
controlaterale al di sotto della lesione
Sensibilità termica
È possibile percepire diverse gradazioni di temperatura.
Queste sono recepite grazie ad almeno tre diversi tipi di
recettori termici posti circa 1mm sotto la pelle:
Recettori per il caldo: sono meno numerosi di quelli per il
freddo e si trovano sotto l’epidermide. Sono fibre C.
Recettori per il freddo: anch’essi sotto l’epidermide. Fibre
A.
Recettori dolorifici: eccitati solo da grandi estremi di caldo
o freddo.
Sono tutti a rapido adattamento e quindi segnalano bene
variazioni di temperatura.
I recettori termici hanno la massima localizzazione in
determinati distretti corporei.
•
Recettori per il caldo: polpastrelli, naso, gomiti
•
Recettori per il freddo: labbro superiore, naso,
mento, dita, petto