
Canali per il K+
Tra tutti i tipi di canale ionici, quelli per il K+ sono certamente i più diversificati.
La spiegazione si trova naturalmente a livello molecolare.
Oggi sappiamo che:
a) hanno in genere struttura tetramerica (4
subunità proteiche, ognuna codificata da un
suo proprio mRNA), e possono essere non
solo omotetrameri, ma anche eterotetrameri
(anche se non tutte le combinazioni sono
possibili)
b) ogni organismo ha numerosi geni che codificano per subunità dei canali del K+
c) ogni gene può dar luogo ad isoforme diverse per splicing alternativo.
Il caso più noto è il gene Shaker di Drosophyla, che contiene 23 esoni.
Ogni subunità del canale Shaker si può presentare in 10 varianti, con una stessa regione
centrale (S1-S5, con regione P) e 5 diverse estremità N- e 2 diverse estremità C-terminali.
d) alle 4 subunitò alfa possono essere associati molti tipi di subunità
accessorie (Kvβ, SUR, mink, ecc.).
Il numero di tipi diversi di canali K+ descritti negli ultimi anni è cresciuto a tal
punto da rendere praticamente impossibile una classificazione su base
funzionale.
Oggi si preferisce raggrupparli, sulla base della loro struttura molecolare (n° di
segmenti transmembranari), in tre categorie.
Primo gruppo: le 4 subunità-α hanno 6 STM. E’ il modulo consueto dei VD ionic
channels, comune ai canali del Na+ e del Ca2+, con S4 sensori del voltaggio.
Questo gruppo comprende:
a) la famiglia dei KV (KV1-KV9, ognuno con diverse varianti)
→ KV1.4 è quello che assomiglia di più al “tipo-A” (ha inattivazione molto rapida).
→ KV3.1 è quello che assomiglia di più al “delayed rectifier” di H&H (inattivazione praticamente assente).
b) la famiglia dei canali del K+ Ca-dipendenti (SK e slo).
L’apertura di canali al K+ (→ ripolarizzazione o iperpolarizzazione) è un altro dei mirabolanti effetti dei canali del
Ca2+ (o della liberazione di Ca2+ dagli organuli intracellulari). Sono stati distinti in tre gruppi in base alla
conduttanza di singolo canale: a bassa γ (bloccati dall’Apamina), a elevata γ (canali “maxi”, bloccati dalla
Caridbotossina), ed a conduttanza intermedia.
c) altri, che interessano varie patologie cardiache, muscolari o nervose. Curiosità: il canale erg (herg nell’Uomo)
presenta la caratteristica di avere l’inattivazione più rapida dell’attivazione, per cui è sempre chiuso (inattivato)
quando la membrana è depolarizzata, e si apre quando la membrana viene iperpolarizzata).
Per molti anni si è pensato che il classico “delayed rectifier” (KV) dell’assone gigante, che
non presenta inattivazione, fosse l’unico canale K+ esistente.
Poi fu scoperto un canale al K+ con inattivazione molto rapida (quasi insensibile al TEA
ma sensibile alla 4AP); questo tipo fu chiamato “canale A” (KA) .
Ma anche il dualismo tra “delayed rectifier” e “tipo A” è caduto, mano a mano che si trovavano canali con
caratteristiche intermedie. Ad esempio, l’espressione in oociti di Xenopus di omotetrameri delle subunità prodotte
da 4 geni di Drosophila ha dimostrato che tra Shaker (il più simile al “tipoA”) e Shaw (il più simile al “delayed
rectifier”) abbiamo i prodotti di Shal e Shab.
La storia del gene Shaker è molto istruttiva. La Drosophila Shaker è un mutante che si riconosce per un caratteristico tremito delle
zampe durante anestesia con etere. L’elettrofisiologia ha dimostrato che l’anomalia stava in un canale del K+ delle fibrocellule
muscolari. La genetica classica ha dimostrato che il gene stava in una certa posizione sul cromosoma X. Allora è stato clonato (nel
“wild type”) quel tratto di DNA, e ci si è trovato dentro un gene per una subunità di canale al K+; questo è stato chiamato “Shaker”.
Altri 3 geni di Drosophila (Shab, Shal e Shaw) sono stati poi trovati per omologia, facendo lo screening dei cDNA ottenuti da una
libreria contenente tutto l’mRNA estratto dalla testa di D. Tutto questo è stato necessario perché i bloccanti dei canali al K+ (TEA,
4AP) non avevano l’affinità necessaria per marcare selettivamente la proteina (come era stato fatto con TTX o bungarotossina o DHP).
Le gates di inattivazione
A) In alcuni canali del K+: dovuta all’azione di “tappo” di un peptide che
costituisce l’estremità N-terminale della subunità a (una “ball”; in realtà 4: uno per
subunità), collegato alla restante parte della molecola da un tratto flessibile (una
“chain”): meccanismo del tipo “ball and chain”.
B) In altri canali del K il peptide è l’estremità N-terminale di una subunità
accessoria (beta).
Il peptide è dotato di cariche parziali
positive, che prenderebbero contatto con
cariche parziali negative speculari
posizionate all’imboccatura intracellulare del
canale.
NB: questo modello presuppone che il canale
prima si apra, e poi si inattivi.
Questo tipo di inattivazione, viene chiamato inattivazione di tipo-N.
NB I canali del Na+ e del Ca2+ hanno un modo di inattivarsi
concettualmente simile, solo che la gate (h) non è l’estremità N-terminale
della molecola, ma un’ansa intracellulare (III-IV per Na+, I-II per Ca2+.
Inattivazione lenta o di tipo C
Con le gates di inattivazione sono stati fatti
vari giochetti, ad esempio:
- la “ball” può essere rimossa <con
trattamento proteolitico o manipolazione
genetica> (l’inattivazione scompare, e può
essere ripristinata aggiungendo la “ball” nel
citosol);
- la “chain” può essere accorciata
(l’inattivazione diviene più rapida, perché la
“ball” impiega meno tempo a trovare
l’imboccatura del canale).
In alcuni canali si è visto che dopo rimozione della N-inattivazione, persiste una
lenta riduzione della corrente al perdurare della depolarizzazione. Inizialmente s’è
pensato che fosse dovuta ad un’analoga azione “di tappo” dell’estremità Cterminale.
Probabilmente l’inattivazione “di tipo C” è dovuta al movimento voltaggiodipendente della porzione esterna del segmento S6, che restringe il filtro di
selettività.
Caratteristiche elettrofisiologiche dei canali KA
Anche il canale KA, come il canale KDR (delayed rectifier), è selettivo
per il K+, ma ha le seguenti caratteristiche:
- Attiva a potenziali più negativi di Kv (attivazione precoce)
- Inattiva tanto più rapidamente quanto maggiore è la depolarizzazione
Correnti di K+ in voltage-clamp
KDR
KA
+20
+40
+40
-20
-20
+20
-20
-20
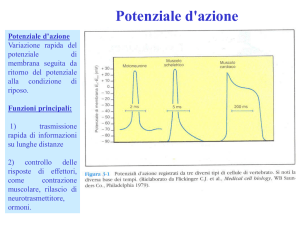
Se lo stimolo depolarizzante è duraturo
nel tempo, invece di un singolo
potenziale d’azione se ne può generare
una serie in sequenza:
scarica o treno di potenziali d’azione
Come fa un neurone a discriminare tra stimoli
di diversa intensità, dal momento che
l’ampiezza del p.d’a. non varia al variare di uno
stimolo sovraliminare (evento tutto o nulla)?
CODIFICAZIONE
Un esempio di codificazione
9 spk/1.2 s
15 nA
1.2 s
13 spk/1.2 s
20 nA
17 spk/1.2 s
30 nA
20 spk/1.2 s
45 nA
Nei neuroni il processo di
codificazione si realizza
variando la frequenza di
scarica al variare
dell’intensità dello stimolo
All’aumentare dell’intensità
dello stimolo aumenta la
frequenza di scarica
Affinchè il processo di codificazione si possa realizzare è necessario
l’intervento di un altro tipo di canale ionico denominato KA
I canali KA operano da freno
voltaggio-dipendente sulla
frequenza di scarica dei
potenziali d’azione
Infatti:
L’attivazione dei canali KA, che è piuttosto precoce,
contrasta l’azione depolarizzante dello stimolo
frena l’insorgere dello spike
ma:
l’inattivazione dei canali KA è tanto più veloce quanto
più intensa è la depolarizzazione
lo stimolo depolarizzante prevale sull’uscita di cariche K+
Esistono dei canali selettivi per il K+ che sono
attivati dal Ca2+ intracellulare
extra
intra
depolarizzazione
extra
intra
ADATTAMENTO
Il processo di adattamento si realizza
con una scarica di tipo fasico
Il processo di adattamento di realizza in seguito all’interazione di
almeno tre diversi tipi di canali ionici selettivi per il K+ (Kv, KA,
KCa) e di canali per il Ca2+ voltaggio-dipendenti
Neuroni diversi possono presentare modalità di
scarica diverse
Scarica tonica: la sua frequenza si
mantiene costantemente
proporzionale all’intensità dello
stimolo
Scarica fasica: la sua frequenza è
proporzionale alla variazione di
intensità dello stimolo
La scarica fasica contiene informazioni relative a variazioni
di intensità dello stimolo
Secondo gruppo: le 4 subunità hanno 2 STM, denominati M1 ed M2.
Questi canali vengono anche denominati “inward rectifier” (KIR) (o “anomalous
rectifier”), perché, quando presenti, sono responsabili della “rettificazione
entrante”.
I canali KIR sono equivalenti al “nocciolo” interno dei KV, quindi mancano del
sensore del voltaggio (S4).
Corrente
0
-120
-80
-40
0
Vm (mV)
40
Di conseguenza, la voltaggio-dipendenza di questi canali non può venire da un
processo di “gating” ma viene da un blocco VD della conduzione, esercitato sul
versante intracellulare da due tipi di “tappi” molecolari: il Mg2+ e molecole
organiche cariche positivamente, come le poliamine.
Domanda: gli IRK sarebbero utili nel pacemaker cardiaco ?
Domanda: cosa fanno gli IRK nel miocardio di lavoro ?
I KIR “classici” vengono chiamati IRK Sono presenti in moltissime cellule. Notare
che al pot. di riposo sono aperti; quindi lasciano passare una (debole) corrente
uscente. Quando presenti, sono i principali responsabili del leakage del K+
Il loro compito è quello di mantenere il pot. di riposo non troppo lontano dal pot. di
equilibrio del K+.
A) Se la m. viene iperpolarizzata, lasciano passare una corrente entrante di K+ che si oppone
all’iperpolarizzazione. B) Se la m. viene depolarizzata, lasciano passare una corrente
uscente di K+ che si oppone (ma sempre più debolmente) alla depolarizzazione.
Alcuni KIR (diagr. C) hanno la particolarità di
essere attivati da proteine-G trimeriche. Per
questa ragione vengono chiamati GIRK.
L’esempio si riferisce all’attivazione di GIRK1
(presente nel pacemaker cardiaco) da parte
dell’acetilcolina (che attiva recettori muscarinici m2, e
poi Gi, che tramite il dimero bg attiva il canale GIRK).
Quindi l’acetilcolina rallenta la frequenza cardiaca con due
meccanismi sinergici ….
I canali KATP, sono chiusi dall’aumento di ATP intracellulare (aperti
dall’aumento di ADP).
Assomigliano molto ai canali attivati dai nucleotidi ciclici (CNGC, attivati
dall’AMPc o dal GMPc) presenti in alcuni recettori sensoriali.
Terzo gruppo: subunità con 4 STM e due regioni P (da cui il nome di K2P).
Si pensa che ci siano due subunità
per canale. Sono i canali “senza
porta” del K+ che abbiamo
incontrato all’inizio (canali di
leakage) in quanto responsabili del
potenziale di riposo.










![Piano di marketing per [Nome prodotto]](http://s1.studylibit.com/store/data/000696763_1-6dc9bde362efff93156d58ddc841c6a8-300x300.png)