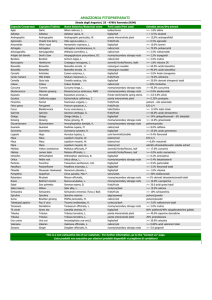
ORGANOGENESI: DIFFERENZIAMENTO
DEI PRIMORDI FOGLIARI
Le foglie si formano nello sviluppo post-embrionale
Cotyledons (embryonic leaves)
Embryo
inside seed
zygote
Germination
Embryonic
development
True leaves
Cotyledons (embryonic leaves)
Post-embryonic
development
Sono formate dal meristema apicale del germoglio (SAM)
SHOOT
APICAL
MERISTEM
True leaves
zygote
Germination
Leaf formation
Il SAM si forma durante l’embriogenesi
Shoot apical meristem
Apical
Basal
TOP DOWN
Shoot apical meristem
Laux, T, Jurgens, G. (1997) Plant Cell 9: 989-1000
Dopo la germinazione il SAM forma le foglie
Leaves
Shoot apex at germination
Post-embryonic
leaf formation
Leaves
Cotyledons
Cotyledons
Reprinted by permission from Macmillan Publishers, Ltd: NATURE. Long, J.A., et al., 379: 66-69, copyright 1996.
diversi stadi nello sviluppo dei primordi
Organogenesi: poche cellule negli strati L1 e L2 nella zona periferica acquistano
l’identità di iniziali (founder cells) della foglia. Cominciano a dividersi più rapidamente
delle cellule circostanti e formano una zona distinta dal resto del doma (primordio).
Sviluppo di differenti regioni nella foglia: regioni dei primordi acquisiscono l’identità
delle diverse parti della foglia. Tre assi di sviluppo: adaxiale/abaxiale; prossimale/distale;
mediale/laterale
Differenziamento di cellule e tessuti: Con la crescita della foglia cellule e tessuti
si differenziano: L1 epidermide; L2 mesofillo; L3 elementi vascolari e cellule della
guaina del fascio
DIVISIONI CELLULARI NELLA FORMAZIONE DEL PRIMORDIO (arabidopsis)
divisioni periclinali nello strato più interno della tunica
divisioni periclinali anche negli strati meno interni della tunica e divisioni meno orientate
divisioni anticlinali nelllo strato esterno della tunica per formare il protoderma
Le foglie in formazione hanno una loro polarità
intrinseca
Peripheral
Central
Leaf
Cot
Cot
Leaf
Assi di asimmetria nella foglia
La polarità è evidente fin dagli stadi iniziali
The adaxial side is towards
the center of the plant
Central
Peripheral
Adaxial
Abaxial
In quale posizione si formano sul germoglio?
(fillotassi)
Alternate
Opposite
Whorled
Spiral
Alternata
TEM of rice apex
Cross section of rice apex
One leaf at a time,
180° apart, as in rice
or other grasses.
Candela, H. et al. (2008) Plant Cell 20: 2073-2087; Itoh, J.-I., et al. (2000) Plant Cell 12:2161-2174
Opposta
Two at a time, 180° apart at
each node. Sometimes pairs
alternate by 90° at successive
nodes.
Verticillata
Three or more leaves at
each node, as in the
horsetail (Equisetum).
Photos courtesy of tom donald
Spiralata
In most plants,
such as this
succulent,
leaves form in
a regular spiral
pattern.
Photos courtesy of tom donald
spiralata
In plants with
spiral phyllotaxy,
leaves form at
about 137° apart.
137°
Spiral phyllotaxy
Spiral phyllotaxy
Spiral phyllotaxy
A line through
sequential
leaves makes a
spiral.
Spiral phyllotaxy
The NEXT leaf
to form is called
the Incipient
primordium (I1).
I1
Spiral phyllotaxy
The one that will
form after that is
called I2….etc.
I2
I1
Fillotassi spiralata in apice di tabacco
I1
Poethig , R.S. and Sussex ,I.M. (1985) The developmental morphology and growth dynamics of the tobacco leaf.
Planta 165: 158-169. Copyright (1985) Planta. Reprinted with kind permission of Springer Science+Business Media.
Cosa determina la posizione del primordio incipiente?
Surgical experiments
demonstrate that leaf
placement is
determined by preexisting primordia
Reinhardt, D., Frenz, M., Mandel, T., and Kuhlemeier, C. (2005) Development 132: 15-26. Reproduced with permission.
I2
This tomato apex shows the positions of several
primordia (P) and incipient primordia (I).
Reinhardt, D., Frenz, M., Mandel, T., and Kuhlemeier, C. (2005) Development 132: 15-26. Reproduced with permission.
P3
I1
P1
I2
P2
I2
This tomato apex shows the positions of several
primordia (P) and incipient primordia (I).
The expected position for I3 (*) can be found by
tracing the spiral.
Reinhardt, D., Frenz, M., Mandel, T., and Kuhlemeier, C. (2005) Development 132: 15-26. Reproduced with permission.
P3
I1
P1
P2
I2
I1 (shown in black) was
surgically isolated from
the rest of the meristem,
by cutting along the red
line.
Reinhardt, D., Frenz, M., Mandel, T., and Kuhlemeier, C. (2005) Development 132: 15-26. Reproduced with permission.
P3
Two days later, the apex
was examined.
I1
P1
P2
I2
Reinhardt, D., Frenz, M., Mandel, T., and Kuhlemeier, C. (2005) Development 132: 15-26. Reproduced with permission.
P3
Instead of emerging at its
expected position (star), I3
shifted towards I1.
I3
I1
P1
P2
I2
This experiment shows
that I1 influences I3
position.
Reinhardt, D., Frenz, M., Mandel, T., and Kuhlemeier, C. (2005) Development 132: 15-26. Reproduced with permission.
P3
I3
I1
P1
P2
I2
Positions of I2 and I3;
older leaves have been
cut away.
Reinhardt, D., Frenz, M., Mandel, T., and Kuhlemeier, C. (2005) Development 132: 15-26. Reproduced with permission.
P3
I3
I1
P1
P2
I2
The older primordia control
the placement of the incipient
primordia.
What kind of signal or information is involved?
L’auxina è coinvolta nella formazione del primordio
Wild-type Arabidopsis
shoot apex. The
meristem is covered
by the leaves it has
produced.
Reprinted from Current Opinion in Plant Biology, 8 (1), Byrne, M.E., Networks in leaf development , 5966, Copyright (2005), with permission from Elsevier .
L’auxina è coinvolta nel determinare la posizione del
primordio
meristem
The apex of the
pin1 mutant is
bare – it fails to
produce lateral
organs.
Wild-type Arabidopsis
shoot apex
pin1 shoot apex
Reprinted from Current Opinion in Plant Biology, 8 (1), Byrne, M.E., Networks in leaf development ,
59-66, Copyright (2005), with permission from Elsevier ; Reinhardt D et al., (2000) Plant Cell 12: 507518
O
OH
IAA
N
H
The pin1 mutant is
defective in the
transport of auxin.
Auxin transport
Auxin (IAA) is a weak acid. At the low pH of
cell walls, it is protonated and uncharged
(IAAH), allowing it to move through the
plasma membrane.
pH 5
pH 7
Cytoplasm
IAAH
Cell wall
Auxin transport
In the less acidic cytoplasm, it loses the
proton, becomes charged (IAA-), and cannot
exit the call by diffusion through the plasma
membrane.
pH 5
pH 7
Cytoplasm
IAAH
IAACell wall
Auxin transport
The PIN1 protein is an
auxin efflux carrier,
transporting charged auxin
back out of the cytoplasm.
pH 5
IAAH
pH 7
IAA-
IAA-
PIN1 protein
Auxin efflux
through PIN1
Auxin transport
The subcellular localization of PIN proteins can be polar and
coordinated between cells, causing directed auxin transport.
In this diagram, the accumulation of PIN1 to the right of each cell
causes a net flow of auxin towards the right.
pH 5
IAAH
pH 7
IAA-
Net flow of auxin
Un massimo localizzato di auxina è richiesto per
l’organogenesi
Applying a spot of
exogenous auxin
(shown as a red
blob) stimulates
outgrowth of
primordium in the
pin1 mutant.
Reinhardt D et al., (2000) Plant Cell 12: 507- 518
38 hours after application 4 days after application
Reinhardt D et al., (2000) Plant Cell 12: 507- 518
Conclusion - Auxin transport and a
local auxin maximum contribute to
organ initiation.
This conclusion is supported by imaging
PIN1 distribution in living plants.
pH 5
IAAH
pH 7
IAA-
Visualizing PIN1 localization
GFP
Excitatory light
Green fluorescent protein
(GFP) emits green light
when excited by blue light.
PIN1
Emitted light
GFP
A protein’s position within a
cell can be determined by
making a fusion protein of it
with GFP, and then looking for
GFP fluorescence.
Visualizing PIN1 localization
Reporter gene
in the nucleus
PIN1 GFP
mRNA
GFP
PIN1
PIN1pro
GFP
GFP
Insertion into
membrane
PIN1
Fusion protein
PIN1
Translation
Visualizing PIN1 localization
PIN1pro
PIN1 GFP
PIN1
Using a confocal laser
scanning microscope,
PIN1:GFP protein distribution
can be imaged in the shoot
apical meristem. In this image,
the green lines show the
position of PIN1:GFP at cell
membranes.
GFP
mRNA
Reproduced with permission - Development Gordon, S.P., Heisler, M.G., Reddy, G.V., Ohno, C., Das,
P., Meyerowitz, E.M. Development, 2007, 134 (19): 3539-3548.
La distribuzione di PIN1 è dinamica durante l’organogenesi
PIN1:GFP
Positions of primoridia
and incipient primordia
The orientation of PIN1 within cells is shown
by white arrows, and indicates auxin flow.
Auxin accumulates at I1 position
Reprinted from Current Biology 15: Heisler, M.G., Ohno, C., Das, P., Sieber, P., Reddy, G.V., Long, J.A., and
Meyerowitz, E.M. Patterns of auxin transport and gene expression during primordium development revealed by live
imaging of the Arabidopsis inflorescence meristem,1899-1911, Copyright (2005), with permission from Elsevier.
La polarità delle proteine PIN 1 determina un massimo di
auxina in I1
I1
I1
This observation is
consistent with the
emergence of a
primordium at the site
of auxin application
dopo l’inizio della formazione del primordio la distribuzione di
PIN1 cambia e orienta il flusso di auxina nel tessuto vascolare
P1
I1
P1
I1
P1
I1
TIME
Poethig , R.S. and Sussex ,I.M. (1985) The developmental morphology and growth dynamics of the tobacco leaf. Planta 165: 158-169. Figure 3 Copyright (1985) Planta.
Reprinted with kind permission of Springer Science+Business Media. Adapted by permission from Macmillan Publishers, Ltd: Nature Reinhardt D., Pesce, E.-R., Stieger, P.,
Mandel, T., Baltensperger, K., Bennett, M., Trass, J., Friml, J., Kuhlemeier, C. Regulation of phyllotaxis by polar auxin transport. Nature 426, 255-260; copyright (2003).
Una successiva inversione nella polarità di PIN1 cambia la
posizione del picco di auxina e specifica la posizione del
nuovo primordio
P3
P3
P1
P1
I1
I1
I2
P2
time
P2
Summary
• Organ initiation at the shoot apical meristem is
determined by auxin distribution and PIN1
• An auxin maximum is necessary and sufficient to specify
the site of primordium formation
• Primordia affect auxin distribution and so placement of
incipient primordia
• Auxin has been proposed to act as a morphogen – a
generator of form
Come viene acquisita l’identità di foglia?
The meristem is a
population of small,
undifferentiated, dividing
cells.
A leaf primordium is a
population of small,
undifferentiated, dividing
cells.
The differ in their
expression of critical
regulatory genes; the
meristem expresses
meristem-specific genes,
and the leaf primordium
expresses primordiumspecific genes.
Ruolo dei fattori di trascrizione
Poethig , R.S. and Sussex ,I.M. (1985) The developmental morphology and growth dynamics of the tobacco leaf. Planta 165: 158-169.
Figure 3 Copyright (1985) Planta. Reprinted with kind permission of Springer Science+Business Media.
Geni di identità meristematica
KNOX-1
Class I KNOX genes (KNOX-1)
•(KNOX means Knotted-like homeobox)
•Expressed in meristem
•Not expressed in incipient primordia
•Help maintain indeterminate growth
Class I KNOX genes
KNOX genes are Knotted-like
homeobox genes that encode
homeodomain transcription
factors.
Hao Yu, H., Yang, S.H., and Goh, C. J. (2000) DOH1, a class 1 knox gene, Is required for maintenance of
the basic plant architecture and floral transition in orchid. Plant Cell 12: 2143-2160.
Espressione di KNOTTED
KNOTTED (a KNOX-1
gene) mRNA accumulates
in the meristem but not
the leaf primordia
(arrows) of Zea mays.
Jackson, D., Veit, B., and Hake, S. (1994) Expression of maize KNOTTED1 related homeobox genes in the shoot apical meristem
predicts patterns of morphogenesis in the vegetative shoot. Development 120: 405–413. Reproduced with permission.
STM un gene di classe KNOX1 è necessario per la
formazione del meristema
Wild-type plant showing
leaf formation at the
shoot apex
The shootmeristemless
mutant (stm) fails to form a
shoot apical meristem during
embryogenesis; notice the
absence of leaf formation.
Reprinted by permission from Macmillan Publishers, Ltd: NATURE. Long, J.A., et al., 379: 66-69, copyright 1996.
Geni Primordio-specifici
ARP
ARP
ARP genes
•“ARP” is derived from three genes, ASYMMETRIC
LEAF1, ROUGH SHEATH2, and PHANTASTICA
•ARP genes encode MYB transcription factors
•Expressed in cells of leaf primordia
• Promote determinate growth and differentiation
ASYMMETRIC
LEAF1 (AS1) mRNA
is expressed in
cotyledons but not in
the meristem .
ARP
ARP
Reprinted by permission from Macmillan Publishers, Ltd: NATURE. Byrne, M.E., et al., 408: 967-971. Copyright 2000..
Wild
type
stm
mutant
In the stm
mutant, AS1 is
expressed in
the meristem
(arrow).
ARP
Reprinted by permission from Macmillan Publishers, Ltd: NATURE. Byrne, M.E., et al., 408: 967-971. Copyright 2000..
L’espressione dei geni KNOX nel meristema
reprime quella dei geni ARP
ARP
KNOX-1
L’espressione dei geni ARP reprime quella dei
geni KNOX
ARP
KNOX-1
La sovraespressione di KNOX-1aumenta la
complessità della foglia e la sua indeterminazione
Arabidopsis
WT
OX
Tobacco
WT
OX
Maize
WT
OX
Chuck G et al., (1996) Plant Cell 8: 1277-1289. Reprinted by permission from Macmillan Publishers, Ltd: NATURE
GENETICS 31: 121 – 122. Hake, S., and Ori, N. Plant morphogenesis and KNOX genes. Copyright (2002).
Mutazioni loss of funcion arp hanno fenotipo simile
alla sovraespressione di KNOX-1
Arabidopsis
Maize
Wild-type
as1
rs2
WT
rs2 WT
rs2
WT as1
Reprinted by permission from Macmillan Publishers, Ltd: NATURE 408: 967-971. Byrne, M.E., Barley, R., Curtis, M., Arroyo, J.M., Dunham, M., Hudson, A., and
Martienssen, R.A. Asymmetric leaves1 mediates leaf patterning and stem cell function in Arabidopsis. Copyright (2000). Reproduced with permission Development
Schneeberger, R., Tsiantis, M., Freeling, M., Langdale, J. Development (1998) 125: 2857-2865.
Nei primordi i geni ARP agiscono come
repressori trascrizionali dei geni di classe KNOX
ARP heterodimer
KNOX gene
Guo, M., et al. (2008) Plant Cell 20:48-58
Geni di confine (boundary) sono necessari per la
separazione degli organi
Boundary genes
•Ensure a sharp boundary between leaf and meristem
•Expressed at organ boundaries
•Loss-of-function leads to “jagged” or fused organs
GENI CUC
JAGGED LATERAL ORGAN (JLO) (LOBD gene family)
JLO expression
JAGGED LATERAL
ORGANS (JLO) is a
boundary gene.
Loss-of-JLO function
causes fused or
jagged organs.
JLO coordinates
KNOX-1 and PIN
activities.
Loss-of-function
phenotype
Summary
• A leaf acquires identity by turning OFF meristem genes
and turning ON leaf genes
• KNOX-1, ARP and boundary genes encode
transcriptional regulators that control expression of other
genes
• Precise control of cell fates involves tight control of
transcription by developmentally regulated activators and
repressors
COME VIENE ACQUISITA LA POLARITA’?
Polarità anatomica e funzionale
Most leaves have polarity –
they are functionally and
anatomically different on their
upper and lower surfaces
Adaxial surface –
light harvesting
CO2
O2
Abaxial surface transpirational water loss,
respiratory gas exchange
Juarez, M. T., Twigg, R.W., and Timmermans, M.C.P. (2004) Development 131:4533-4544. Reproduced with permission.
Peripheral
Central
Leaf
Leaf
Leaves have an inherent polarity because one side is more
central and one more peripheral.
Peripheral
Central
Leaf
Adaxial
Abaxial
The central side is adaxial, and peripheral is abaxial.
How does a leaf “know” which side is central?
The Sussex signal
In the 1950s, Ian Sussex showed
that a signal from the meristem is
required for proper leaf polarity.
Reprinted, with permission, from the Annual Review of Plant Physiology and Plant Molecular Biology,
Volume 49 (c) 1998 by Annual Reviews www.annualreviews.org
P3
I1
I3
P1
P2
Incipient primordia
were surgically
isolated from the
rest of the
meristem by a
small incision
I2
Reinhardt, D., Frenz, M., Mandel, T., and Kuhlemeier, C. (2005) Development 132: 15-26. Reproduced with permission.
P3
I1
I3
P1
P2
I2
The isolated
primordium lost
polarity (it became
entirely abaxialized)
and became
radially-symmetrical.
P3
I1
I3
P1
P2
I2
A more recent experiment showed that laser ablation
of only the epidermal cell layer is sufficient for the
primordium to lose its adaxial polarity.
QuickTime™ and a
TIFF (Uncompressed) decompressor
are needed to see this picture.
Reinhardt, D., Frenz, M., Mandel, T., and Kuhlemeier, C. (2005) Development 132: 15-26. Reproduced with permission.
•A signal from the
meristem moves
through the epidermis
into the incipient
primordium.
•The signal conveys
the adaxial positional
information.
•The nature of the
signal is not known.
Controllo genetico della polarità
The phantastica
mutant of Antirrhinum
(snapdragon) gives
important clues to the
basis of leaf polarity.
Wild-type
phan
Waites, R., and Hudson, A. (1995) Development 121: 2143 – 2154. Reproduced with permission.
Wild-type leaf
phan mutant leaf
The phantastica mutant has radially symmetrical leaves.
Waites, R., and Hudson, A. (1995) Development 121: 2143 – 2154. Reproduced with permission.
Wild-type leaf
phan mutant leaf
Mutant phan leaves are abaxialized, indicating that PHAN
is necessary for adaxial cell fate.
Tutte le foglie radialmente simmetriche sono abaxializzate?
P3
I1
I3
P1
P2
I2
Surgical isolation
phan mutant leaf
The phan mutant leaves resemble the surgically
isolated leaf primordia –
No: I mutanti Loss of function kanadi hanno foglie
radiali adaxiali
•A triple mutant kanadi
1,2 and 3 has radial,
adaxialized leaves
•KANADI genes
promote abaxial cell
fate
Eshed Y et al., Izhaki, A., Baum, S.F., Floyd, S.K., and Bowman, J.L. (2004) Development 131: 2997-3006. Reproduced with permission.
La perdita della identità adaxiale o abaxiale determina la
radializzazione
Wild type
phan mutant
kan mutant
Eshed Y et al., Izhaki, A., Baum, S.F., Floyd, S.K., and Bowman, J.L. (2004) Development 131: 2997-3006. Reproduced with permission.
Fenotipo del mutante Gain-of-function phb-1d
Gain-of-function phb-1d mutants
have radial, adaxialized leaves.
McConnell, J.R. and Barton, M.K. (1998) Development 125: 2935-2942. Reproduced with permission.
La mutazione The phb-1d mutation inluenza la
distribuzione del mRNA di PHB
In gain-of-function phb-1d
In wild-type plants, PHB
mutants, PHB is expressed
expression is restricted to
everywhere, resulting in
the adaxial side of the
adaxialized, radially
leaves
symmetric leaves.
Longitudinal section Cross section
SEM
Cross section
Reprinted by permission from Macmillan Publishers, Ltd: NATURE. McConnell, J.R., Emery, J.,
Eshed, Y., Bao, N., Bowman, J., and Barton, M.K. Nature 411: 709-713, copyright 2001.
PHB promuove l’identità adaxiale
Wild-type leaf:
PHB
expression
= Adaxial
No PHB
expression
= Abaxial
phb-1d leaf
PHB
expression
= Adaxial
Reprinted by permission from Macmillan Publishers, Ltd: NATURE. McConnell, J.R., Emery, J.,
Eshed, Y., Bao, N., Bowman, J., and Barton, M.K. Nature 411: 709-713, copyright 2001.
come PHB, PHV and REV promuovono l’identità adaxiale
Like PHB, REVOLUTA
(REV) and PHAVOLUTA
(PHV) are expressed in
the adaxial domain.
Reprinted from Current Biology 13, Emery, J.F., et al., Radial patterning of Arabidopsis shoots by Class III
HD-ZIP and KANADI genes, 1768–177, Copyright (2003), with permission from Elsevier.
come PHB, PHV and REV promuovono l’identità adaxiale
Loss of function triple
phb / phv / rev
mutant has radial,
abaxialized leaves
Reprinted from Current Biology 13, Emery, J.F., et al., Radial patterning of Arabidopsis shoots by Class III
HD-ZIP and KANADI genes, 1768–177, Copyright (2003), with permission from Elsevier.
La polarità richiede la corretta espressione di PHB
Too little PHB
Too much PHB
Borghi, L., et al.,(2007) Plant Cell 19:1795-1808.
La espressione di PHB è regolata da miRNA
AAAAAAA
miRNAs are short (~21-22
nt) RNAs that, in
association with
ARGONAUTE (AGO)
target specific mRNAs for
degradation (or interfere
with translation).
AAAAAAA
AAAAAAA
Controllo della espressione di PHB da parte di
miRNA
x
AAAAAAA
miR166
PHB mRNA
PHB-1D mRNA
AAAAAAA
In wild-type plants,
miR166 binds to the PHB
mRNA and degrades it
on the abaxial side of
the leaf primordium.
In phb-1d plants, base changes
in the PHB mRNA prevent
miR166 from binding to it,
allowing it to accumulate
throughout the leaf primordium.
Reprinted by permission from Macmillan Publishers, Ltd: NATURE. Kidner, C.A. and Martienssen, R.A. Nature 428: 81-84, copyright 2004.; McConnell, J.R.,
Emery, J., Eshed, Y., Bao, N., Bowman, J., and Barton, M.K. Nature 411: 709-713, copyright 2001.
I miRNA controllano la polarità della foglia
Additional support for role of miRNA
in leaf polarity comes from the fact
that the ago1 mutant has radial
leaves; AGO is needed for miR166
function.
AAAAAAA
In ago mutants, as in phb-1D
mutants, PHB mRNA
accumulates throughout the leaf
primordia.
ago mutant
phb-1D mutant
Reprinted by permission from Macmillan Publishers, Ltd: NATURE. Kidner, C.A. and Martienssen, R.A. Nature 428: 81-84,
copyright 2004; McConnell, J.R., Emery, J., Eshed, Y., Bao, N., Bowman, J., and Barton, M.K. Nature 411: 709-713, copyright 2001.
I miRNA contribuiscono alla determinazione della polarità
della foglia
These experiments demonstrate that in
Arabidopsis miRNAs contribute to leaf polarity,
by controlling the distribution of critical mRNAs.
AAAAAAA
Cosa fanno tutti questi geni?
ADAXIALIZING GENES
PHANTASTICA encodes a
MYB transcription factor;
PHB/ PHV/ REV genes
encode HD-ZIP III
transcription factors
ABAXIALIZING GENES
KANADI 1,2,3 encode GARP
transcription factors;
YABBY genes have similar
function and also encode
putative transcription factors
The genes
regulated by these
transcription
factors are not yet
known.
geni YABBY (FIL, YAB2, YAB3)
promuovono il differenziamento del lato abaxiale
delle foglie (SONO FATTORI DI TRASCRIZIONE ZINC –FINGER)
Mutanti yab3 (omozigoti) producono foglie lobate che esprimono KNOX1 e formano
meristemi ectopici e mostrano conversione abaxiale/adaxiale
Il fenotipo yab suggerisce che ci sia incompatibilità tra la funzione
KNOX e le funzioni che specificano l’identità ABAXIALE nella
foglia
Modello per l’acquisizione della polarità
miR166
KAN, YAB
PHAN or PHB/PHV/REV
Abaxial fate
Adaxial fate
Meristem-derived signal
Il patterning ad/abaxiale può influenzare tratti
agronomicamente importanti
Wild-type rice leaf.
Sclerenchymatous
tissue forms on the
abaxial surface and
supports the leaf in an
open form.
Zhang, G-H. et al., (2009) Plant Cell 21:719-735
SLL1 è una proteina GARP che influenza la
polarità e l’arrotolamento della foglia
Wild-type
sll1 mutant
In sll1 mutants, the supportive sclerenchymatous tissues on the
abaxial surface do not form, causing the leaf to roll inwards.
Zhang, G-H. et al., (2009) Plant Cell 21:719-735
Wild-type
sll1 mutant
Rice plants with
rolled leaves (like
sll1) can have
more erect leaves,
reduced water
loss by
transpiration and
higher yields.
Zhang, G-H. et al., (2009) Plant Cell 21:719-735
SUMMARY
• Leaves are initiated from cells in the shoot apical meristem
• Auxin gradients are important in leaf primordium initiation and
positioning
• Leaf identity is determined by a change in expression of
transcription-factor encoding genes
• Leaf polarity requires an unknown signal from the meristem
and the domain-specific expression of adaxial- and abaxialspecific transcription factors
GENI DI IDENTITA’ ADAXIALE/ABAXIALE
PHANTASTICA (anthirrinum)/ AS1 (arabidopsis)
mutazioni loss of function phan promuovono la conversione
adaxiale/abaxiale
(organi a simmetria radiale)
I trascritti sono localizzati nel lato adaxiale
Geni di identità adaxiale
I mutanti phan hanno anche difetti nel mantenimento del
meristema
Interdipendenza destino adaxiale / meristematico
mutazione phan
PHABULOSA, PHAVOLUTA, REVOLUTA
Homeodomain leucin zipper proteins
HD-ZIPIII
contenenti un dominio START che lega lipidi
Fattori di trascrizione specifici per il lato
adaxiale
PHB PHV REV Sono inizialmente espressi in maniera continua dal centro del SAM fino
ai primordi fogliari; successivamente la loro espressione si restringe al lato adaxiale
della foglia
Mutazioni loss-of-function nei geni PHB o PHV o REV determinano conversione
adaxiale/ abaxiale e incapacità di formazione o mantenimento del SAM
Le funzioni PHB, PHV, REV sembrano positivamente correlate alla funzionalità dei geni
KNOX
Interdipendenza destino adaxiale / meristematico
La localizzazione nella regione adaxiale dei trascritti HD-ZIP III
è regolata da:
Geni KANADI (KAN) espressi nella regione abaxiale
(mutazioni recessive KAN determinano adaxializzazione e espressione
ectopica di PHAV, PHAB, REV)
microRNA
(mutazioni dominanti PHB e PHV inibiscono il riconoscimento e la
degradazione dei trascritti genici ad opera di miRNA espressi
specificamente nella regione abaxiale)
Regolazione da micro RNA dei geni HD-ZIP III
miR165
miR166
Mutazioni in queste regioni danno luogo a fenotipi dominanti con foglie adaxializzate,
radiali e meristemi più grandi