I recettori sensoriali
Recettori
Strutture di natura
proteica poste sulla
membrana o nel citoplasma
Cellule specializzate per
la trasduzione di stimoli
di varia natura in segnali
elettrici
Vista
Sensi
speciali
Udito,
equilibrio
Olfatto
Gusto
Sensi
somatici
Tatto,Dolore,
Temperatura
Posizione
del corpo
Proprio
cettiva
Recettore:
struttura
specializzata
stimolata da modificazioni ambientali
esogene ed endogene
E’ in grado di trasformare diversi tipi di energia in impulsi nervosi che
vengono inviati al Sistema Nervoso
Centrale
L’informazione viene qui decodificata
ed
analizzata,
determinando
la
sensazione
Stimolo adeguato: variazione ambientale
che, con minima incidenza energetica
specifica, genera l’impulso nel recettore
Meccanismi di codificazione
dello stimolo
Codificazione della qualità e della
posizione
dello stimolo: ogni modalità
sensoriale possiede una via specifica (“via
privata”)
sia nel midollo spinale che
nell’encefalo (densità recettoriale)
Codificazione dell’intensità dello stimolo:
modificazione della frequenza di scarica
e della popolazione di recettori attivati
Sintonizzazione delle frequenze: gli
inputs sensoriali sono proiettati in più
aree encefaliche
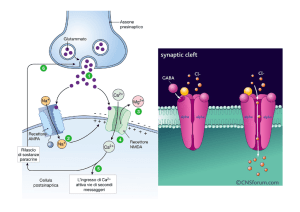
Trasduzione dello stimolo
in impulsi nervosi
La stimolazione del recettore determina
l’apertura dei canali del Na+ e del K+ (potenziale
recettoriale graduato); quando il potenziale
recettoriale raggiunge il valore soglia (potenziale
generatore) si ingenera il potenziale d’azione. Se
lo stimolo è sotto-liminale, l’evento elettrico non si
propaga ma influenza elettrotonicamente le
regioni vicine
Relazione stimolo-risposta: la maggior parte dei
recettori funziona entro un ampio range di intensità
di
stimolazione.
L’entità
della
risposta
è
proporzionale al logaritmo dell’intensità della
stimolazione
Trasmissione degli inputs sensoriali
Fibre nervose: A,B,C (diverse per ∅ e
velocità di conduzione)
-Propriocettori e recettori tattili: fibre di tipo A
-dolore acuto: fibre A;
-dolore persistente: fibre C
Recettori tattili: fibre Aβ
(Stimolazioni pressorie grossolane fibre C)
Afferenze cinestesiche (capsule articolari
e legamenti): fibre A
Afferenze termiche: terminazioni nervose
libere non mielinizzate. Il numero di recettori è
scarso; sono pertanto necessari notevoli
fenomeni di sommazione spaziale
Arco riflesso
Interneurone
(a livello spinale o encefalico)
Neurone afferente
Neurone efferente
Struttura recettoriale
Effettore
Circuiti nervosi presenti
nei mammiferi
Divergente: un neurone influenza
molti altri neuroni
Convergente: un neurone riceve
informazioni da un’ampia gamma
di altri neuroni
Parallelo: un singolo impulso può
avviare una serie di impulsi in
altri neuroni
Esempi di recettori sensoriali
strutture specializzate osservabili a a livello
superficiale (cute,sottocute) o in profondità
(muscoli, articolazioni,tendini, visceri) con
proprietà di trasformare differenti tipi di
energia in impulsi elettrici
Classificazione dei recettori
in base al tipo di stimolo a cui sono sensibili
Segnalano deformazioni
meccaniche del recettore
o delle cellule adiacenti al
recettore stesso
Meccanocettori
Termocettori
Nocicettori
Segnalano variazioni della
temperatura (alcuni sensibili al freddo e altri al caldo)
Segnalano un danno tissutale
sia di natura fisica che chimica
(recettori dolorifici)
Recettori elettromagnetici
Chemocettori
Segnalano stimoli luminosi a livello della retina
Segnalano stimoli di natura
chimica (gustativi, olfattivi,
pO2 e pCO2, osmolarità
dei liquidi corporei).
Meccanocettori
Sensibilità tattile della cute
Terminazioni nervose libere
Terminazioni ad estremità espansa
Dischi di Merkel
Terminazioni arboriformi
Corpuscoli di Ruffini
Terminazioni capsulate
Corpuscoli di Meissner
Terminazioni nervose annesse ai peli
Sensibilità profonde
Terminazioni nervose libere
Terminazioni ad estremità espansa
Terminazioni arboriformi
Corpuscoli di Ruffini
Terminazioni capsulate
Corpuscoli di Pacini
Terminazioni muscolari
Fusi neuromuscolari
Organi tendinei del Golgi
Udito
Recettori cocleari
Equilibrio
Recettori vestibolari
Pressione arteriosa
Barocettori dei seni carotidei e aortici
Sensibilità tattile della cute
Sensibilità profonde
Sensibilità profonde
Fuso
neuromuscolare
Organo tendineo di Golgi
Udito
Equilibrio
Pressione arteriosa
Termocettori
Rispondono alla temperatura
FRIGOCETTORI
CALOCETTORI
Dubbi sulla identità anatomica:
Corpuscoli di Krause (freddo)
Terminazioni libere (caldo)
Temperatura di attivazione:
•FREDDO tra 12 e 37°C (max 32). Poi riprendono a scaricare tra 45 e 50°C
Freddo paradosso: immersi in una vasca con acqua
a 42°C, se viene messa acqua a 47°C si avverte
un freddo intenso
•CALDO tra 20 e 47°C (max tra 38 e 43°C)
Sensazioni trasportate da fibre Aδ e C (lente)
Se si tocca un oggetto freddo e
lo si allontana rimane la sensazione
di freddo
La percezione della temp. è
influenzata da quella della cute
Nocicettori
Terminazioni nervose libere
Nella cute sono suddivisi in due classi:
MECCANONOCICETTORI rispondono solo a
stimoli meccanici
NOCICETTORI PLURIMODALI rispondono a
stimoli dolorifici di varia natura (termici, chimici ecc.)
Sono in grado di generare un
incremento della risposta quando
lo stimolo viene prolungato nel
tempo iperalgesia
Chemocettori
Gusto
Gemme gustative
Olfatto
Recettori dell’epitelio olfattivo
Concentrazione di O2 e CO2 nel sangue
Osmolarità dei liquidi organici
Classificazione dei recettori
in base alla localizzazione
Propriocettori
Esterocettori
Enterocettori
Si trovano nei muscoli, tendini,
articolazioni e danno luogo ad
informazioni che riguardano i
movimenti e la posizione
del corpo nello spazio.
Sono rappresentati da tutti i recettori cutanei (tattili, dolorifici,
termici) e danno informazione dei
cambiamenti che avvengono nelle
immediate vicinanze della cute.
Inoltre, a questo gruppo appartengono anche i recettori specifici
della retina, dell’orecchio interno,
della mucosa nasale: essi percepiscono a distanza le variazioni
dell’ambiente lontano dall’animale.
Trasmettono impulsi originati dagli
organi interni (visceri cavi, cuore,
ecc.).
Attivazione dei recettori sensoriali
Si ha in seguito all’incontro con uno stimolo
specifico detto stimolo adeguato: questo è
caratteristica unica di ogni recettore.
Es: fotorecettori→luce
recettori auditivi→onde sonore
In realtà in alcuni casi il recettore può essere attivato
anche da stimoli non specifici ma la risposta non è funzionale. Es: forte compressione sul globo oculare crea una
sensazione luminosa.
Come fanno tipi differenti di recettori
a rilevare tipi differenti di stimoli?
stimoli
La specificità del recettore è dovuta a
- caratteristiche molecolari della membrana
- struttura del recettore
- particolare organizzazione del recettore
con altre cellule accessorie
Ognuno dei principali tipi di sensazioni di cui
possiamo avere esperienza (dolore, tatto,
visione, suoni ecc.) si definisce
MODALITA’ SENSORIALE
… ma le fibre nervose trasportano solo
impulsi elettrici …
come è possibile che vengano
trasmesse differenti modalità
sensoriali?
Ciascuna via nervosa afferente va a terminare in un’area specifica del S.N.C e il tipo
di sensazione che viene avvertita è deter-
minato da quella particolare area del
S.N.C alla quale giunge la via nervosa
Fantaipotesi: facendo terminare il nervo ottico dove
termina l’acustico e viceversa si potrebbe vedere il
tuono e sentire il fulmine.
Relazione tra intensità dello stimolo e
potenziale recettoriale
L’ampiezza aumenta rapidamente all’inizio, ma
successivamente, a più alte intensità di stimolo,
aumenta progressivamente sempre meno.
Uno stimolo molto debole di solito riesce a generare almeno qualche impulso, mentre una stimola
zione molto intensa causa incrementi progressivamente minori della frequenza dei potenziali
d’azione
Il recettore è in grado di discriminare le variazioni di intensità di stimoli deboli assai meglio
di quelle di stimoli molto forti.
Ciò permette al recettore di avere un campo
di intensità di risposta estremamente
ampio a livelli molto bassi
Meccanismi di insorgenza dei
potenziali recettoriali
c Deformazione meccanica del recettore
d Azione di una sostanza chimica sulla membrana
e Modificazioni di temperatura della membrana
f
Effetti di radiazioni elettromagnetiche
Modificano le caratteristiche della
membrana del recettore e permettono il
flusso di ioni attraverso i suoi canali
Corpuscolo di Pacini
E’ situato prevalentemente sotto la cute.
E’ formato da una terminazione nervosa amielinica
incapsulata da diversi stra
ti di tessuto connettivo
(tipo cipolla).
La terminazione appartiene ad una fibra nervosa
mielinica.
Lo stimolo adeguato per il corpuscolo di Pacini
è costitutito da pressioni profonde applicate alla
cute.
Queste pressioni agiscono sulla capsula di tessuto
connettivo e quindi sulla terminazione nervosa determinando nella membrana di quest’ultima l’apertura di canali del sodio.
Il conseguente ingresso di ioni Na+ depolarizza la
membrana stessa e produce il potenziale gene-
ratore.
Se questo è di sufficiente ampiezza, potrà depolarizzare la membrana del primo, più vicino nodo di
Ranvier sino alla soglia dando inizio ad un poten-
ziale d’azione.
L’intensità dello stimolo è codificata
come frequenza di scarica dei
potenziali d’azione
CODICE DI FREQUENZA DELL’INTENSITA’ DELLO STIMOLO
Il numero dei potenziali
d’azione insorgenti nella
unità di tempo è funzione
dell’intensità della pressione (aumenta il numero
dei canali Na+ aperti).
Altro meccanismo:
CODICE DI POPOLAZIONE
L’aumento di pressione
determina l’attivazione
di un maggior numero di
corpuscoli di Pacini, in
quanto l’aumento di pressione va ad interessare
un area cutanea più vasta.
Il sistema nervoso risponde a
stimoli ripetitivi con
meccanismi di adattamento
Spesso quando uno stimolo viene applicato in
modo continuo, l’encefalo gradatamente ne
perde coscienza.
Ciò avviene ad esempio a fronte di un rumore
di fondo (ticchettio di un orologio): dopo un
certo tempo non lo si avverte più.
Questo fenomeno è denominato adattamento sensoriale e può avvenire sia con un meccanismo cerebrale che con variazioni insorgenti a livello recettoriale.
Esistono:
RECETTORI A RAPIDO ADATTAMENTO
Esempio: Corpuscolo di Pacini
RECETTORI A LENTO ADATTAMENTO
Esempio: Fuso e recettori articolari
Come si spiega l’esistenza di recettori
a lento e a rapido adattamento?
Occorre ammettere che i recettori a lento
adattamento siano in grado di mantenere
costante,
per
tutta
la
durata
dell’applicazione dello stimolo un certo
grado di depolarizzazione, mentre nei
recettori
a
rapido
adattamento
il
potenziale precipita rapidamente.
L’intimo
meccanismo
che
porta
all’adattamento
sembra
legato
alle
specifiche proprietà dei recettori, ma il
problema per il momento non può dirsi
risolto.
Le modalità di adattamento sono caratteristiche
specifiche di ciascun tipo di recettore.
Tra i meccanocettori, quello meglio studiato è il
corpuscolo di Pacini.
Quando si applica una forza deformante questa viene
trasmessa ed evoca il potenziale recettoriale.
Dopo pochi centesimi di secondo il liquido extracel-
lulare all’interno del corpuscolo (struttura viscoelastica)
si ridistribuisce e la pressione risulta dissipata per
cui il potenziale recettoriale non viene più generato, nonostante la compressione continui.
Quando la forza deformante viene rimossa si producono
eventi opposti.
Il corpuscolo di Pacini genera segnali all’inizio e
alla fine della compressione
Il tatto
La pelle è particolarmente sensibile a ogni
forma di contatto.
Possono essere distinte sensazioni tattili
di diversa natura, come il caldo, il freddo,
la pressione e il dolore grazie ai vari tipi di
recettori
Nella fisiologia classica il tatto viene annoverato tra i “sensi minori”.
In realtà questa è una semplificazione
erronea:
ciascun movimento, soprattutto l’afferrare e
il tenere in mano gli oggetti, si realizza grazie
alla collaborazione finemente coordinata di
sensibilità e motilità
Cellule di Merkel: sono i più semplici sensori di tatto.
Sono localizzati nello strato basale della epidermide.
Trasmettono gli impulsi che ricevono ad una
singola fibra nervosa (sensori proporzionali).
Registrano la pressione esercitata sulla cute.
Fondamentali per la sensibilità fine.
Esempio: discriminazione lettere in rilievo
Corpuscoli di Meissner:Al confine tra epidermide e
derma. Costituiti da una decina di cellule incolonnate tra le quali decorre una fibra afferente. Non
reagiscono a pressione persistente ma solo a variazioni (sensori differenziali).
Corpuscoli di Ruffini: Nel derma. Ammassi cellulari appiattiti, contengono un fitto groviglio di fibre nervose
dotate di numerose giunzioni. Somigliano agli organi del
Golgi e anch’essi misurano la tensione del tessuto. Reagiscono maggiormente agli stimoli veloci rispetto a quelli lenti ma non si adattano mai
completamente (sensori proporzionali-differenziali).
Corpuscoli di Pacini: Ancora più in profondità.
Ciascun corpuscolo è composto da un guscio che
contiene fino a 60 strati di cellule appiattite
che circondano una grossa terminazione nervosa centrale. Essa scarica solo all’inizio e al termine di una sollecitazione, per tale ragione i corpuscoli
sono particolarmente sensibili alle vibrazioni, in cui
direzione e velocità dello stimolo variano continuamente.
Terminazioni nervose annesse ai peli. Molti mammiferi
possiedono, vicino agli occhi e alla bocca, peli
particolarmente sviluppati (vibrisse ) che possiedono
funzione sensoriale. Molto sviluppati negli animali
notturni e in quelli che vivono in tunnel sotterranei
Su ogni centimetro quadrato di pelle mediamente circa 130 recettori tattili
Fusi neuro-muscolari
Presentano 2-10 fibre intrafusali (in
parallelo con le fibre extrafusali) ancorate
ad un’estremità all’endomisio di quest’ultime
ed innervate da motoneuroni Aγ. La
distensione
delle
fibre
extrafusali
determina la deformazione della zona
equatoriale
e
la
stimolazione
delle
terminazioni anulospirali (fibre Aα) che
attivano i motoneuroni per le fibre
extrafusali (lunghezza statica)
Ai lati delle anulospirali possono essere presenti le terminazioni a fiorame (fibre Aβ)
con soglia di attivazione più elevata;
stimolano i motoneuroni dei flessori ed
inibiscono i motoneuroni degli estensori
ipsilaterali. L’effetto è opposto sull’arto
controlaterale (lunghezza dinamica).
Fuso neuro-muscolare
Innervazione motrice e sensitiva
del fuso-neuromuscolare
Organo tendineo del Golgi
e fuso neuro-muscolare
Organi muscolo-tendinei del Golgi
Terminazioni nervose situate fra le
lacinie del tendine e pertanto in serie
rispetto alle fibre muscolari
Mediante fibre sensitive Ib inviano
inputs ad un interneurone inibitorio che
sinapta con il motoneurone per lo stesso
muscolo, inibendolo; è anche presente la
stimolazione dei motoneuroni per i
muscoli antagonisti
Vengono stimolati sia dallo stiramento
passivo
del
muscolo
(anche
se
scarsamente) che,soprattutto, dalla sua
contrazione, durante la quale scaricano
continuamente al fine
di evitare la
lacerazione delle fibre muscolari