Gluconeogenesi
Il glucosio può essere sintetizzato a partire da precursori più semplici,
i precursori possono essere vari a seconda della specie vivente, nei
mammiferi sono: piruvato, lattato, amminoacidi, glicerolo.
Fegato e reni
È operativa in condizioni di digiuno estremo (dopo le 24 h) → forte ipoglicemia e
dopo deplezione delle riserve di glicogeno endogeno.
La gluconeogenesi condivide con la glicolisi tutte le reazioni reversibili
le quali procedono in direzione opposta quando utilizzate nella sintesi
Le reazioni irreversibili della glicolisi sono
aggirate attraverso l’azione di enzimi differenti
PIRUVATO COME PRECURSORE GLUCONEOGENICO
2 piruvato + 2 NADH + 4 ATP + 2 GTP + 6 H2O + 2 H+ →
Glucosio + 2 NAD+ + 4 ADP + 2 GDP + 6Pi
Bicarbonato
Piruvato
carbossilasi
+
piruvato
a
Ossalacetato
Il piruvato prodotto dalla glicolisi è
immediatamente trasportato nel
mitocondrio dove può subire diversi
destini metabolici.
Se è necessario utilizzarlo per la isntesi
di glucosio deve essere carbossilato a
ossalacetato
Dentro il mitocondrio può
alimentare il ciclo di Krebs
o essere utilizzato per
sintetizzare glucosio
Ossalacetato
NADH + H+
NAD+
H
Malato
deidrogenasi
mitocondriale
Malato
l
OH
citosol
Malato
deidrogenasi
citosolica
NAD+
NADH + H+
Si genera una scorta di
NADH citosolici che sono
necessari per il proseguimento
della gluconeogenesi
Ossalacetato
gluconeogenesi
Reazioni inverse della
glicolisi
(stessi enzimi e stessi
intermedi metabolici)
Il piruvato è riconvertito a
PEP (fosfoenolpiruvato)
attraverso una via alternativa
(un bypass alla piruvato
chinasi)
glicolisi
L’ossalacetato nel citosol è convertito in fosfoenolpiruvato
dalla FOSFOENOLPIRUVATO CARBOSSICHINASI
Ossalacetato
2-fosfoglicerato
Enolasi
Fosfoenolpiruvato
Il glucosio neosintetizzato è liberato dagli epatociti nel torrente circolatorio per
raggiungere i tessuti extraepatici che sono in carenza di glucosio (in particolare il
cervello)
gluconeogenesi
Reazione alternativa
Glucosio 6-fosfato
Reazione alternativa
glicolisi
Il lattato prodotto nei muscoli in esercizio può essere un precursore
gluconeogenico nel fegato (Ciclo di Cori).
Vari amminoacidi sono precursori del glucosio durante il digiuno, per es.
l’alanina, che partecipa al Ciclo Glucosio/Alanina con cui il piruvato
prodotto nei tessuti extraepatici raggiunge il fegato per essere indirizzato
verso la gluconeogenesi
Piruvato
Glutammato
ALANINA
Transamminazione extra-epatica
α-chetoglutarato
Alanina trasportata al fegato
attraverso il circolo sanguigno
ALANINA
α-chetoglutarato
Piruvato
Glutammato
Transamminazione epatica, il piruvato
può essere utilizzato per sintetizzare
glucosio
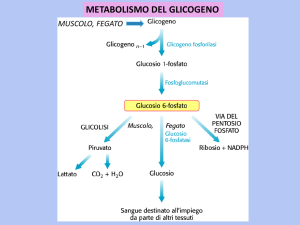
DEGRADAZIONE DEL GLICOGENO
Glicogeno fosforilasi
Scinde i legami glicosidici α(1→4) attraverso una fosforolisi a partire
dalle estremità non riducenti e liberando glucosio 1-fosfato
Glicogeno n unità
cofattore: piridossal-fosfato (PLP), che aiuta
l’enzima a scindere il legame C-O)
Glucosio 1-fosfato
Glicogeno n-1 unità
si ferma a 4 residui dal punto di
ramificazione lasciando una
destrina limite
Interviene l’enzima bifunzionale
deramificante
Scinde il legame α(1→6) liberando
glucosio non fosforilato
Gli ultimi 3 residui del ramo sono
trasferiti sull’estremità libera della
catena principale
La fosfoglucomutasi converte il glucosio 1-fosfato in glucosio 6-fosfato
Glucosio 6-fosfato
Glicolisi
Via dei pentoso-fosfati
Nel fegato viene defosforilato a glucosio e liberato
nel circolo sanguigno per essere trasportato ai
tessuti che lo richiedono
Glucosio 6-fosfatasi
Glucosio
Sintesi del glicogeno
Fegato e muscoli nei vertebrati
Glucosio 6-fosfato
Fosfoglucomutasi
Glucosio 1-fosfato
UDP-glucosio
pirofosforilasi
Il glucosio 1-fosfato deve essere attivato
per essere incorporato nella molecola di
glicogeno. Viene convertito in UDPglucosio
Donatore di glucosio all’estremità nonriducente delle catene di glicogeno
Glicogeno sintasi
Aggiunge, una alla volta, unità di α-D-glucosio alla
catena polisaccaridica crescente
Richiede un innesco (catena di 7 residui di
glucosio legati da legami α(1→4)
L’innesco con la sua estremità
riducente è attaccato ad un
residuo di Tyr della proteina
GLICOGENINA
Estremità non riducente
del Glicogeno (n unità)
Glicogeno (n + 1 unità)
La glicogenina è sia l’impalcatura su cui si
costruisce la molecola di glicogeno, sia l’enzima che
catalizza la sintesi dell’innesco.
GLICOGENINA
la glicogenina ha attività glucosil-trasferasica
GLICOGENINA
la glicogenina estende la catena ripetendo 6 volte la
reazione
Ramificazioni
Interviene l’enzima ramificante [amilo-(1,4→1,6)
transglicosilasi]
Rimuove un frammento di almeno 6 residui dall’estremità non riducente della catena
principale e lo riattacca a questa con legame α(1→6) nel punto di ramificazione.
I rami devono essere distanziati almeno 4 residui, si creano numerose estremità non
riducenti che contribuiscono ad aumentare la velocità di degradazione o di
allungamento del glicogeno
La via dei pentoso fosfati
Sfrutta il glucosio per produrre ribosio 5-fosfato
e equivalenti riducenti in forma di NADPH
è la via con cui i pentosi introdotti con la dieta entrano
nel metabolismo
Ribosio 5-fosfato
NADPH
Sintesi di RNA, DNA, ATP…, NAD+, NADP+, FAD,
FMN, Coenzima A,
Biosintesi di: ac. grassi, colesterolo, neurotrasmettitori,
nucleotidi.
Detossificazione: riduzione del glutatione ossidato;
citocromo p450 monoossigenasi
si realizza in due fasi: OSSIDATIVA (genera NADPH)
NON-OSSIDATIVA (interconversione degli zuccheri)
Fase ossidativa: produce NADPH
Glucosio-6-fosfato + 2 NADP+ + H2O → ribulosio 5-fosfato + 2 NADPH + CO2 + 2H+
glucosio-6-fosfato
deidrogenasi
ossidazione del C-1
del G6P: formazione
di un legame estere
intramolecolare
6-fosfo-gluconolattonasi
idrolisi del legame
estere
6-fosfo-gluconato
deidrogenasi
ossidazione del C-3 e
decarbossilazione
La [NADP+] ha un ruolo fondamentale nel determinare il destino del glucosio 6-fosfato
se nella cellula la
[NADPH] è molto
maggiore della [NADP+]:
NADPH compete con NADP+ per il sito
attivo della Glucosio 6-fosfato
deidrogenasi inibendo l’enzima.
bassa attività degli enzimi della fase
ossidativa della Via dei Pentoso-fosfati
(manca l’accettore di e-)
il Glucosio 6-fosfato è indirizzato verso la
glicolisi o la sintesi di glicogeno
La produzione di NADPH è strettamente associata al suo utilizzo nelle reazioni di
biosintesi (es.: sintesi degli acidi grassi)
Fosfopentoso
isomerasi
Ribosio 5-fosfato
Se sono necessarie grandi
quantità non solo di NADPH
ma anche di ribosio 5-fosfato
per la sintesi di nucleotidi il
ribulosio 5-fosfato è
isomerizzato a ribosio 5fosfato nella fase nonossidativa
Fase non ossidativa: rappresenta un raccordo con la glicolisi,
smaltisce i pentoso-fosfati formati nella prima fase
produce intermedi della glicolisi, dai quali si può riottenere il
glucosio 6-P
(ogni 6 molecole di glucosio 6-fosfato che vengono ossidate, nella
via dei pentoso-fosfati, se ne riproducono 5)
Ribulosio 5-fosfato
isomerasi
Ribosio 5-fosfato
epimerasi
xilulosio 5-fosfato
Fruttosio 6-P
Gliceraldeide 3-P
transchetolasi
transaldolasi
Sedoeptulosio 7-P
(zucchero a 7 atomi
di carbonio)
Eritrosio 4-P
transchetolasi
Gliceraldeide 3-P
Fruttosio 6-P
Sono utilizzati come intermedi della glicolisi o della gluconeogenesi: il
Glucosio 6-fosfato rigenerato con la gluconeogenesi può essere
utilizzato per alimentare di nuovo la via dei pentoso-P
sono tutte reazioni REVERSIBILI che collegano la via dei
pentoso-fosfati alla glicolisi/gluconeogenesi e vicerversa
La via dei pentoso-fosfati è versatile
e può avere diverse finalità
glucosio
6-P
fruttosio
6-P
2NADP+
glucosio
6-P
fruttosio
1,6-bis-P
Diidrossiacetone
fosfato
2NADPH
Ribulosio
5-P
Gliceraldeide
3-P
Fabbisogno di Ribosio 5-P maggiore
rispetto a quello di NADPH:
- la glicolisi è dominante rispetto alla
via dei pentoso-P,
- una buona parte della gliceraldeide
3-P e del Fruttosio 6-P sono utilizzati
per produrre ribosio 5-P con le
reazioni inverse della fase nonossidativa
Tymoczko et al. Principi di Biochimca. Zanichelli editore S.p.A 2010
Fabbisogno equivalente di
Ribosio 5-P e di NADPH:
- è preferita la fase ossidativa
della via dei pentoso fosfati che
termina con la conversione del
ribulosio 5-P in ribosio 5-P
glucosio
6-P
2NADP+ 2NADPH
2NADP+ 2NADPH
fruttosio
6-P
ribulosio
5-P
glucosio
6-P
ribosio
5-P
fruttosio
6-P
fruttosio
1,6-bis-P
Diidrossiacetone
fosfato
ribulosio
5-P
ribosio
5-P
fruttosio
1,6-bis-P
Gliceraldeide
3-P
Diidrossiacetone
fosfato
Fabbisogno di NADPH maggiore
rispetto a quello di Ribosio 5-P:
- la via dei pentoso-fosfati avviene
completamente
- il ribosio 5-P è riciclato come glucosio
6-P attraverso la via non-ossidativa e
parte della via gluconeogenica
(fegato).
Tymoczko et al. Principi di Biochimca. Zanichelli editore S.p.A 2010
Gliceraldeide
3-P
2 ATP
PIRUVATO
Fabbisogno equivalente di NADPH e di
ATP:
- il ribosio 5-P ottenuto dalla via dei
pentoso-fosfati è trasformato in
piruvato attraverso la via non-ossidativa
e parte della via glicolitica.
Non tutte le cellule hanno le stesse esigenze di NADPH e ribosio 5fosfato,
es.: fegato, tessuto adiposo, ghiandole mammarie che mantengono
attiva un’intensa biosintesi di acidi grassi hanno necessità di grandi
quantitativi di NADPH; cellule che si riproducono molto velocemente
hanno necessità di grandi quantità di ribosio 5-fosfato.
TESSUTI IN CUI E’ ATTIVA LA VIA DEI PENTOSO-FOSFATI
Ghiandola surrenale
Fegato
Ovaio/Testicoli
Tessuto adiposo
Ghiandola Mammaria
Eritrociti
sintesi steroidi
sintesi ac. grassi e colesterolo
sintesi steroidi
sintesi ac. grassi
sintesi ac. grassi
mantenimento del glutatione
ridotto
il NADPH prodotto dalla via dei pentoso-fosfati ha un ruolo fondamentale nel
sistema di omeostasi redox che serve a proteggere le cellule dagli attacchi di
agenti ossidanti (ROS) prodotti in condizioni di stress ossidativo
H2O2
Glutatione
reduttasi
Glutatione perossidasi
All’interno degli eritrociti il sistema glutatione-perossidasi/glutatione-reduttasi
è l’unico sistema di difesa dai ROS mantenendo l’ambiente intracellulare in uno
stato riducente.
il sistema GSH/GSSG controlla il livello di meta-Hb (la formazione di metaemoglobina può avvenire a causa della presenza di idroperossidi, che si originano
spontaneamente in presenza di alte concentrazioni di O2).
il sistema GSH/GSSG mantiene nello stato ridotto i gruppi sulfidrilici (-SH) della
emoglobina,
>>> in condizioni ossidanti si possono formare derivati ossidati delle cisteine e
ponti disolfuro fra molecole di Hb, che causano la formazione di grossi aggregati
(Corpi di Heinz) i quali vanno a depositarsi sulla membrana eritrocitaria
deformandola e danneggiandola insieme all’azione dei ROS > ANEMIA
EMOLITICA.
La carenza di glucosio 6-fosfato deidrogenasi (G6PDH) può causare
anemia emolitica in condizioni di stress ossidativo (per es.: con
assunzione di farmaci che producono ROS, favismo), perché gli eritrociti
non producono abbastanza NADPH per mantenere il glutatione nel suo
stato ridotto