I Carboidrati
I carboidrati sono macromolecole organiche molto abbondanti in natura, che servono a svolgere
funzioni sia metaboliche che strutturali. Dal punto di vista chimico, sono poliidrossialdeidi o
poliidrossichetoni, e in base alla loro complessità sono distinti in:
Monosaccaridi
Oligosaccaridi (fino a 10 unità)
Polisaccaridi (oltre le 10 unità)
Il glucosio rappresenta il monosaccaride più importante per la biochimica dei mammiferi, essendo
il combustibile principale. Il ribosio, il galattosio e il glicogeno sono altri zuccheri con rilevanza
biomedica.
La maggior parte dei carboidrati - detti anche saccaridi o glucidi - è riconducibile alla formula
chimica generale Cn (H2O)n. Hanno almeno un gruppo aldeico o chetonico e funzioni alcoliche
coniugate a ciascun atomo di carbonio. I monosaccaridi non possono essere idrolizzati a saccaridi
più semplici. Diverse unità di monosaccaride portano alla formazione di oligosaccaridi e
polisaccaridi. Le forme più diffuse in natura del glucosio (il monosaccaride biochimicamente più
importante) sono il beta-glucopiranosio (62%) e l'alfa-glucopiranosio (37%). La disposizione
equatoriale piuttosto che assiale dell'ossidrile anomerico giustifica la maggiore stabilità
dell'anomero beta rispetto all'alfa.
Il principale ruolo biochimico dei carboidrati è di fornire energia (circa 4 kcal per grammo), oltre
ad importanti ruoli strutturali. Dopo l'assorbimento intestinale, il glucosio è ridistribuito ai diversi
tessuti ed organi. Ormoni specifici, quali insulina e glucagone, intervengono per regolare i valori
glicemici e per mantenerli costanti; inoltre, sono molto importanti per il metabolismo glucidico e i
processi di trasporto transepiteliale: la molecola di glucosio passa dal lume intestinale
all'enterocita, poi al sangue ed infine alle cellule dell'organismo.
Nessun carboidrato è essenziale! In quanto possono essere comunque sintetizzati dall'organismo
a partire da altre molecole, tuttavia svolgono funzioni fondamentali nell'organismo. Per esempio,
il metabolismo neuronale si basa prevalentemente sul glucosio: infatti il cervello ne consuma
circa 120 grammi al giorno.
L'amido è il polisaccaride principale, si trova soprattutto nelle patate e nei cereali ed è costituito
solo da glucosio (lineare con legami α-1-4 glicosidici e ramificata, l'amilopectina, con legami α-1-6
glicosidici). La digestione dei carboidrati complessi inizia in bocca, grazie ad enzimi
idrolitici presenti nella saliva e si completa a livello intestinale per l'intervento di enzimi secreti
dal pancreas o presenti nella parete degli enterociti. La saliva è composta principalmente
da acqua, mucina (lubrificante per la masticazione) e l'enzima α-amilasi salivare; essa serve a
idratare i cibi secchi e a creare un ambiente idoneo per l'attacco enzimatico. L'azione
dell'amilasi viene interrotta nello stomaco a causa della forte acidità, infatti il succo gastrico non
effettua alcuna degradazione dei carboidrati. L'intestino tenue è la maggiore sede
dell'assorbimento dei prodotti della digestione.
Le sostanze idrosolubili, come gli zuccheri, vengono trasferite all'interno dell'organismo
attraverso il sistema portale epatico; quelle liposolubili raggiungono invece il sangue attraverso
i vasi linfatici ed il dotto toracico. Il passaggio dei nutrienti avviene prima nella membrana apicale
(fase 1), poi in quella baso-laterale (fase 2) e infine avviene l'ingresso nelle cellule dei vari tessuti.
Il trasporto attivo di glucosio è mediato dalla proteina SGLT (Sodium Glucose Transporters), essa
fa entrare nella cellula gli ioni di sodio. La pompa del sodio elimina lo ione consumando ATP. Per
facilitare il trasporto del glucosio esiste nel nostro organismo una "famiglia" di molecole chiamate
GLUT, che si trovano generalmente nella membrana apicale delle cellule, e poi effettuano il
trasporto entrando nel torrente circolatorio. Quando la glicemia è bassa, la captazione è favorita
da alcuni organi e tessuti, rispetto ad altri (la priorità nel consumo di monosaccaridi ce l'ha il
cervello). Il più importante trasportatore della famiglia dei GLUT è il GLUT 4: si trova nelle cellule
adipose, nel muscolo scheletrico e cardiaco, ed è attivato dall'insulina in condizioni di glicemia
(glucosio plasmatico) elevata; GLUT-4 incrementa fino a 20-30 volte la velocità d'ingresso del
glucosio in queste cellule.
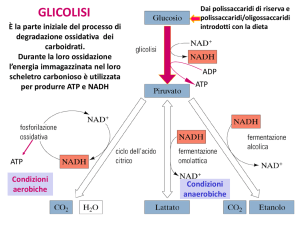
Le funzioni della glicolisi sono: la produzione di ATP e intermedi necessari per l'anabolismo; la
regolazione dipende da 2 fattori: Il fabbisogno cellulare di ATP e il fabbisogno degli intermedi
metabolici. La resa energetica della glicolisi anaerobica è di 2 molecole di ATP. In condizioni di
aerobiosi, invece, se ne formano 2 + 36 (2 derivate dall'ossidazione del glucosio a piruvato, e 36
grazie alla respirazione mitocondriale, ovvero fosforilazione ossidativa). Considerando la
variazione di energia libera ottenuta da ciascun legame fosfoanidride dell'ATP, si può calcolare
come il rendimento della glicolisi aerobia sia nettamente superiore (80% circa) rispetto a quello
della glicolisi anaerobia (20% circa).
La glicolisi la possiamo suddividere in 3 fasi: la prima fase dove avviene prima la fosforilazione del
glucosio in glucosio-6-fosfato (grazie all'enzima esochinasi, la reazione consuma una molecola di
ATP); successivamente, il glucosio-6-fosfato diventa fruttosio-6-fosfato che a sua volta si
"addiziona" con il fruttosio 1,6-difosfato (fino a questo punto la glicolisi invece di produrre
energia ne ha richiesta); adesso comincia la seconda fase in cui il fruttosio si "rompe" per formare
2 molecole più semplici: diidrossiaceton fosfato e gliceraldeide 3-fosfato (l'enzima che catalizza
questa reazione è l'aldolasi), in un secondo momento la molecola di diidrosssiaceton fosfato viene
trasformata in gliceraldeide 3-fosfato (avviene grazie al triosofosfato isomerasi),: solo ora il
gliceraldeide 3-fosfato si trasforma prima in 1,3-difosfoglicerato (l'enzima richiesto è la
gliceraldeide 3-fosfato deidrogenasi e l'agente ossidante invece è il NAD+), e poi in 3fosfoglicerato con produzione di 2 molecole di ATP e acido piruvico (piruvato); poi comincia
la terza fase, in cui il 3-fosfoglicerato trasferisce il gruppo fosfato dal C3 al C2 del glicerato (2fosfoglicerato), che a sua volta si trasforma in fosfoenolpiruvato, qui avviene una disidratazione
che porta alla liberazione di energia tramite una molecola di acqua creando un gruppo fosfato ad
alta energia (gluconeogenesi o neoglucogenesi, essa si può verificare nel fegato in condizioni
di ipoglicemia; il glucagone blocca la glicolisi inducendo la fosforilazione ed inattivazione di 2
enzimi, la fosfofruttochinasi 2 e la piruvato chinasi), ed infine il fosfoenolpiruvato viene trasferito
all'ADP per formare una molecola di ATP (l'enzima responsabile è la piruvato chinasi).
La gluconeogenesi è il processo opposto della glicolisi, ovvero la sintesi di glucosio a partire dalle
molecole organiche di natura non glucidica (protidi, lattato, glicerolo). Se la glicolisi e la
gluconeogenesi avvenissero contemporeanamente avverrebbe soltanto uno spreco di energia
chimica. Per la glucogenesi sono fondamentali gli amminoacidi (soprattutto alanina), il lattato e
il glicerolo. Per la neoglucogenesi gli enzimi che la regolano sono: piruvato carbossilasi,
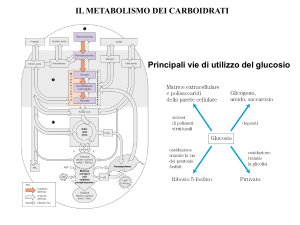
fosfoenolpiruvato carbossichinasi, fruttosio 1,6-bifosfatasi e glucosio-6-fosfatasi. Il glucosio-6fosfato può essere inoltre metabolizzato in un'altra via ossidativa, che avviene a livello citosolico,
definita via dei pentoso fosfati o shunt dei pentosi. Ha 2 funzioni:
1. Produrre pentosi, necessari per la sintesi di macromolecole quali acidi nucleici;
2. Produrre equivalenti riducenti sotto forma di NADPH, necessari per i processi anabolici
cellulari.
Nel complesso in questa via metabolica, chiaramente attraverso più cicli, il glucosio viene
ossidato a CO2, senza alcuna produzione di ATP. Questa via metabolica ha importanza
significativa nei tessuti (adiposo, fegato, corteccia surrenale, testicoli, e ghiandola mammaria) che
producono attivamente acidi grassi e colesterolo, oltre che negli eritrociti, dove il NAPH
contribuisce a mantenere la disponibilità di ferro (II).
Il rendimento della glicolisi dall'ossidazione del glucosio nell'ambito della glicolisi e del ciclo di
Krebs ha un rendimento elevato, vicino al 70%.
A cura di,
Gabriele Grassadonia