Parte III - Metabolismo delle proteine e degli amminoacidi
Risposte e breve discussione
1)
2)
3)
4)
5)
6)
7)
8)
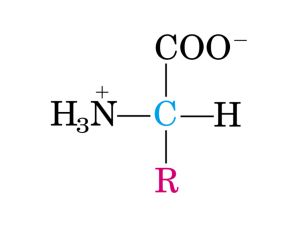
b. Gli amminoacidi essenziali per l'uomo sono quelli che non possono essere biosintetizzati
nell'organismo. Per questo motivo tali amminoacidi debbono essere introdotti con
l'alimentazione. Gli amminoacidi che rispondono a tali caratteristiche sono: treonina, lisina,
valina, leucina, isoleucina, metionina, fenilalanina e triptofano. Istidina e arginina sono
considerati semiessenziali ovvero amminoacidi la cui biosintesi è insufficiente a supportare
adeguatamente la crescita del bambino. Il fabbisogno giornaliero medio degli amminoacidi
essenziali è di circa 7-800 mg. Non è vero che gli amminoacidi strutturalmente più semplici
siano necessariamente essenziali né che da questi derivino tutti gli altri amminoacidi.
a. La prolina, unico imminoacido tra i venti che costituiscono le proteine, può essere
sintetizzata in tre passaggi a partire da glutammato. Questa via sintetica è inversa rispetto a
quella attiva nel catabolismo della prolina. La prolina può essere prodotta anche da
aspartato, serina e metionina ma ciò avviene attraverso un percorso fatto da un numero
cospicuo di reazioni. Dalla leucina non si può invece produrre prolina.
a. Le L-amminoacido ossidasi sono enzimi che catalizzano specificamente la deaminazione
ossidativa degli L-amminoacidi utilizzando FMN come coenzima. Le loro caratteristiche
cinetiche sono, tuttavia, inadeguate per un'efficiente deaminazione degli amminoacidi. In
particolare, questi enzimi possiedono una scarsa affinità di legame per il substrato (valori di
Km molto elevati) con valori di Vmax particolarmente ridotti. Non hanno quindi rilevanza
nell'ambito del catabolismo degli amminoacidi.
d. La carbammilfosfato sintetasi I è l'enzima che catalizza la biosintesi del carbammilfosfato
destinato alla produzione di urea all'interno del mitocondrio degli epatociti. La biosintesi
mitocondriale del carbammilfosfato avviene a partire da CO2 , NH3 e ATP ed è stimolata dal
N-acetilglutammato, un effettore allosterico positivo dell'enzima. Esiste anche una
carbammilfosfato sintetasi II, citosolica, che partecipa alla biosintesi delle basi
pirimidiniche. Quest'ultimo enzima utilizza glutammina come donatore di azoto e non è
attivato da N-acetilglutammato. La molecola di carbammilfosfato non presenta legami
estere.
c. I due atomi di azoto presenti nella molecola di urea derivano dal carbammilfosfato e
dall'aspartato. Il carbammilfosfato mitocondriale si genera attraverso una reazione che
impiega l'ammoniaca formata, per la maggior parte, dalla deaminazione ossidativa del
glutammato, catalizzata dalla glutammato deidrogenasi. Il secondo atomo di azoto si
aggiunge quando la citrullina, formata per condensazione del carbammilfosfato con
l'ornitina, reagisce con l'aspartato generando argininsuccinato. Il passaggio successivo
prevede la rottura di quest'ultima molecola con formazione di arginina e fumarato.
L'arginina, in una reazione catalizzata dall'arginasi, libera urea rigenerando
contemporaneamente ornitina. Questa può dare inizio ad un nuovo ciclo di reazioni mediante
condensazione con un'altra molecola di carbammilfosfato.
c. La leucina è l'unico amminoacido esclusivamente chetogenetico. Nel suo catabolismo si
producono acetacetato e acetil CoA. In tutti gli altri casi si genera almeno un composto da
cui è possibile sintetizzare glucosio. Il catabolismo della fenilalanina porta alla formazione
di acetacetato e fumarato. Quello dell'istidina ad α-chetoglutarato e imminoformile. La
serina può essere convertita in piruvato attraverso reazioni di deaminazione non ossidativa.
L'isoleucina, infine, viene catabolizzata a propionil CoA e acetil CoA.
a. Il formiato si libera nel catabolismo del triptofano. In particolare, nella reazione
catalizzata dalla chinurenina formilasi che porta da N-formil chinurenina a chinurenina.
e. La reazione tra i due amminoacidi arginina e glicina catalizzata dalla arginina-glicina
transamidinasi, porta alla formazione di ornitina e guanidinacetato. Da quest'ulimo si genera
9)
10)
la creatina attraverso una reazione di N-metilazione, dipendente da S-adenosilmetionina.
Aspartato e carbammilfosfato sono substrati dell'aspartato transcarbammilasi, enzima
implicato nella biosintesi delle basi pirimidiniche. Carbammilfosfato e ornitina si
condensano a formare citrullina nel ciclo dell'urea. Succinil CoA e glicina sono implicate
nella biosintesi di δ-amminolevulinato, un metabolita della biosintesi dell'eme.
a. La tappa limitante nella biosintesi dell'eme è quella catalizzata dalla ALA sintasi. La
presenza dello stesso eme determina infatti una diminuzione della sintesi dell'enzima con
associato un possibile effetto inibitorio per retroazione. Molti farmaci inducono un aumento
della sintesi dell'ALA sintasi a livello epatico. La maggior parte di questi farmaci è
metabolizzata nel fegato da un sistema che utilizza il citocromo P450. Durante il
metabolismo di questi farmaci si impiegano grandi quantità di eme per sintetizzare il
citocromo P450 e ciò determina una diminuzione dei livelli intracellulari dell'eme con
conseguente derepressione dell' ALA sintasi.
b. La bilirubina diretta è definita tale per il fatto di reagire direttamente con il diazoreagente
di Erhlich. Il primo ad osservare questo fenomeno fu Van den Bergh il quale, per errore,
dimenticò di aggiungere il metanolo alla miscela di reazione per il dosaggio dei pigmenti
biliari. La bilirubina diretta è quella coniugata con acido glicuronico, molecola che rende
maggiormente solubile la bilirubina favorendone l'eliminazione con la bile. Gli itteri
emolitici sono caratterizzati da un aumento prevalente della bilirubina indiretta che si
produce dalla trasformazione della biliverdina. Solo successivamente la bilirubina è
coniugata con glicuronato a livello del fegato. I danni epatici sono invece causa, piuttosto
che effetto, di un aumento della bilirubina nel siero.