Rassegne
Polimorfismo C825T del gene GNB3 che codifica
la subunità β3 delle proteine G e rischio cardiovascolare
Michelangelo Sartori, Emanuela Parotto, Giulio Ceolotto, Italia Papparella, Livia Lenzini,
Lorenzo A. Calò, Andrea Semplicini
Hypertension is a common disorder of multifactorial origin that constitutes a major risk factor
for cardiovascular events such as stroke and myocardial infarction. The subunits of the heterotrimeric G proteins are attractive candidate gene products for both susceptibility to essential
hypertension and interindividual variation in blood pressure. A polymorphism (825C/T) in exon
10 of the GNB3 gene, that encodes for the β3 subunit, has recently been described. The 825T allele
is associated with alternative splicing of the gene and formation of a truncated but functionally
active β3 subunit. Carriers of the 825T allele appear to have an increased risk for hypertension,
obesity, insulin-resistance and left ventricular hypertrophy. Moreover, 825T allele carriers
respond with a stronger decrease in blood pressure to therapy with a thiazide diuretic and with
clonidine. GNB3 825T allele may be regarded as a potential genetic marker for a better definition of the risk profile of hypertensive subjects, but further studies are needed to precisely define
the impact of T allele on the prognosis of such patients.
(Ann Ital Med Int 2004; 19: 240-248)
Key words: Genetic predisposition; G proteins; Hypertension; Insulin-resistance; Left ventricular hypertrophy; Obesity.
Introduzione
e l’endotelina6. Le proteine G sono espresse ubiquitariamente e la differente combinazione tra le molteplici isoforme delle subunità ne determina la funzione7. L’attivazione delle proteine G da parte dei recettori provoca la rottura del legame tra la subunità α e il dimero βγ. In tal modo entrambi acquisiscono la capacità di attivare molteplici
proteine effettrici. Per esempio Gαq attiva la fosfolipasi Cβ,
responsabile della produzione di diacilglicerolo e inositolotrifosfato, i quali provocano, rispettivamente, l’attivazione della proteinchinasi C e l’aumento del Ca++ intracellulare; quest’ultimo è a sua volta responsabile della contrazione del muscolo liscio8.
Tra le numerose funzioni cellulari regolate dalle proteine
G, vi è anche la regolazione dell’antiporto Na+/H+9. Lo
scambio Na+/H+ è effettuato da una famiglia di proteine
transmembrana ubiquitarie, denominate NHE, che scambiano uno ione H+ intracellulare con uno ione Na+ extracellulare10. L’attività di NHE è aumentata nel 30-50% dei
pazienti ipertesi in diversi tipi di cellule circolanti (piastrine,
leucociti, eritrociti)11-14. Tuttavia non tutti i pazienti ipertesi hanno una maggiore attività di NHE, per cui il solo
aumento pressorio non è la causa della variazione dell’attività di questa proteina. I geni che codificano per le numerose isoforme di NHE non sono né mutati né “iperespressi” negli ipertesi15. Quindi la differente attività di
NHE, riscontrata negli ipertesi, può essere spiegata solo
da un’alterata regolazione dell’antiporto.
Studiando le isoforme delle proteine G eterotrimeriche
nei pazienti con elevata NHE, è stato scoperto un poli-
L’ipertensione è un fenotipo complesso causato dall’interazione di molteplici geni con lo spettro praticamente infinito delle possibili variazioni ambientali: risulta quindi
difficile risalire a cause molecolari precise1,2. Sono stati
studiati numerosi geni, i cui prodotti partecipano alla regolazione della pressione arteriosa attraverso fattori umorali, nervosi, emodinamici e renali; in particolare, ne è stata misurata l’associazione con i differenti fattori di rischio cardiovascolare3. Le mutazioni a carico di singoli nucleotidi dei geni codificanti le subunità delle proteine G
sembrano essere alcuni dei candidati più promettenti per
lo studio della componente genetica dell’ipertensione arteriosa4. Le proteine G, infatti, sono coinvolte nella regolazione della pressione arteriosa e nel rimodellamento
vascolare5.
Proteine G
Le proteine G, così chiamate per la capacità di legare la
guanosina trifosfato (GTP) quando attive, sono formate da
tre subunità differenti (α, β, γ) e costituiscono gli effettori
dei recettori a sette domini transmembrana, responsabili
dell’attivazione di molteplici vie di trasduzione del segnale;
in particolare, tali proteine mediano gli effetti intracellulari di molti ormoni vasocostrittori come l’angiotensina II
Clinica Medica 4 (Direttore: Prof. Achille Cesare Pessina), Dipartimento
di Medicina Clinica e Sperimentale, Università degli Studi di Padova
© 2004 CEPI Srl
240
Michelangelo Sartori et al.
morfismo del gene che codifica la subunità β316. Questo
gene (GNB3) è situato sul cromosoma 12p16. Siffert et al.16
hanno documentato che in alcuni pazienti ipertesi caratterizzati da NHE elevato, è presente una timina al posto
di una citosina nella porzione 825 del gene GNB3. La mutazione puntiforme C825T è associata a “splicing alternativo”, cioè ad un più corto RNA messaggero, responsabile della sintesi di una subunità β3 più piccola e più attiva rispetto alla subunità β3 nativa17.
nista α-adrenergico19 o dopo infusione dell’agente vasocostrittore metilergonovina maleato20 e una maggiore vasocostrizione nel territorio cutaneo dopo infusione di endotelina-1, di angiotensina II e di noradrenalina rispetto
ai soggetti con genotipo CC21. L’allele T sembra quindi
amplificare l’effetto di molte sostanze vasocostrittrici.
La subunità β3 è implicata anche nell’aggregazione
piastrinica stimolata da adrenalina. Ex vivo, infatti, le piastrine di pazienti con genotipo TT/TC presentano un’aggregazione maggiore, dopo stimolazione con adrenalina
e adenosindifosfato, rispetto a quelle dei pazienti con genotipo CC22.
Infine il dimero βγ è implicato nella regolazione della
risposta infiammatoria, scatenando la chemiotassi. Infatti
la chemiotassi dei neutrofili, stimolata da interleuchina823 o da lipopolisaccaridi24, risulta maggiore nei soggetti portatori dell’allele T rispetto ai soggetti con genotipo
CC. Parimenti la proliferazione linfocitaria, dopo stimolazione antigenica con il virus dell’epatite B, è maggiore
nei pazienti con genotipo CT/TT25.
Da tutti questi studi si può, quindi, concludere che il polimorfismo C825T non costituisce soltanto una curiosità
genetica ma assume anche una rilevanza biologica.
Il polimorfismo C825T del gene GNB3
Il polimorfismo C825T è presente in tutte le popolazioni
finora analizzate, ma la frequenza varia ampiamente a
seconda della zona geografica. Negli europei, l’allele T è
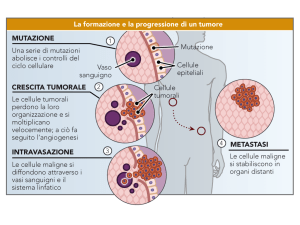
presente con una frequenza che si approssima al 30%, negli asiatici si avvicina al 50%, negli africani all’80% (Fig.
1)18. La ragione di questa diversa distribuzione dell’allele T non è chiara. Si ipotizza che questo allele possa essere stato scelto dai meccanismi evolutivi per migliorare
una specifica funzione che attualmente non si conosce.
Numerosi studi hanno dimostrato che la subunità β3 mutata, rispetto alla subunità nativa, è associata ad un’aumentata sensibilità delle cellule ad alcuni ormoni il cui sistema di trasduzione comporta l’attivazione della cascata delle proteine G4. Tale ipotesi è supportata da esperimenti in vitro che hanno evidenziato un aumento del legame del GTPγS, indice di attivazione delle proteine G,
nelle cellule di pazienti con allele T, indicando così un’aumentata attività delle proteine G in tali pazienti16. I pazienti
portatori dell’allele T presentano una maggiore vasocostrizione nel circolo coronarico dopo infusione di un ago-
Polimorfismo C825T ed ipertensione
Numerosi studi epidemiologici hanno indagato l’associazione dell’allele 825T con l’ipertensione arteriosa16,26-42.
L’associazione è stata studiata, inizialmente, attraverso studi caso-controllo, dove si confrontava la frequenza dell’allele T tra popolazioni di ipertesi e normotesi (Tab.
I)16,27,31,34,35-41 o dove si confrontavano i valori pressori tra
FIGURA 1. Frequenze dell’allele T in differenti aree geografiche.
241
Ann Ital Med Int Vol 19, N 4 Ottobre-Dicembre 2004
riscontrata nelle popolazioni asiatiche, in particolare nei
giapponesi36-38, nei cinesi39 e nelle popolazioni di neri afroamericani40 e di nativi canadesi41.
Le ragioni della discrepanza dei vari studi sono molteplici. La quasi totalità degli studi sono trasversali, di tipo
caso-controllo, affetti quindi da possibili bias di selezione. Gli effetti dell’allele T potrebbero essere stati coperti dall’effetto della terapia farmacologica e dalla selezione operata nella coorte di controllo. Essi hanno misurato
la frequenza dell’allele T in popolazioni di ipertesi e di normotesi ma i criteri di definizione di ipertensione erano differenti da studio a studio. Inoltre le popolazioni studiate
erano estremamente eterogenee: includevano infatti pazienti in età avanzata, pazienti in trattamento farmacologico, pazienti obesi e pazienti diabetici. In tutti questi
studi, quindi, i fattori ambientali potrebbero aver mascherato l’effetto dei fattori genetici.
Recentemente il nostro gruppo di ricerca ha realizzato
uno studio prospettico in una popolazione omogenea di giovani pazienti ipertesi con ipertensione di grado I42. La popolazione è stata scelta in modo tale da eliminare fattori
confondenti come l’età, la terapia farmacologica ed even-
pazienti con e senza allele T (Tab. II)26-28,32,34,35,38,39. I risultati sono stati contrastanti. Per quanto riguarda l’Europa
Occidentale, si è dimostrata un’associazione tra allele T
ed ipertensione arteriosa in popolazioni di origine tedesca16,26-28, spagnola29 e polacca30. Tuttavia uno studio in
popolazioni di origini francese e irlandese non ha documentato l’associazione31. Uno studio prospettico è stato
condotto in una popolazione finlandese ma non ha rilevato
alcuna associazione tra allele T e pressione arteriosa sia
all’inizio che alla fine di un follow-up durato 4.2 anni32.
Tuttavia la frequenza dell’allele T inaspettatamente bassa e l’eterogeneità della popolazione studiata non danno
allo studio la forza sufficiente per escludere tale associazione.
Uno studio condotto in una popolazione di pazienti austriaci affetti da ipertensione essenziale non ha inoltre
documentato alcuna associazione tra allele T e sviluppo
di crisi ipertensive33.
Per quanto riguarda le popolazioni extraeuropee, l’associazione tra allele T ed ipertensione è stata documentata
nei bianchi dell’Australia34 e nei neri inglesi di origine caraibica o africana35. Nessuna associazione è stata invece
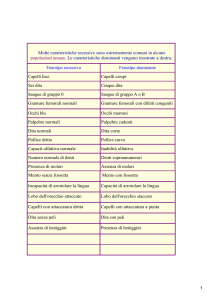
TABELLA I. Frequenza dell’allele T del gene GNB3 nei pazienti ipertesi e nei soggetti normotesi.
Autore
N. pazienti
al.16
Siffert et
Beige et al.27
Brand et al.31
Benjafield et al.34
Dong et al.35
Kato et al.36
Tozawa37
Ishikawa et al.38
Huang et al.39
Larson et al.40
Hegele et al.41
853
1479
1354
299
428
1424
359
352
1165
904
447
Nazionalità
Frequenza dell’allele T
Tedeschi
Tedeschi
Francesi
Bianchi australiani
Neri inglesi
Giapponesi
Giapponesi
Giapponesi
Cinesi
Afroamericani
Nativi canadesi
p
Normotesi
Ipertesi
0.25
0.28
0.31
0.25
0.76
0.50
0.56
0.52
0.52
0.75
0.50
0.31
0.35
0.35
0.43
0.87
0.49
0.63
0.52
0.52
0.74
0.57
< 0.01
< 0.01
NS
< 0.05
< 0.01
NS
0.04
NS
NS
NS
NS
TABELLA II. Valori della pressione arteriosa in base al polimorfismo (C825T) dell’esone 10 del gene GNB3.
Autore
N. pazienti
al.26
Schunkert et
Beige et al.27
Hengstenberg et al.28
Snapir et al.32
Benjafield et al.34
Dong et al.35
Ishikawa et al.38
Huang et al.39
608
1000
2658
908
299
428
352
1165
Pressione arteriosa (mmHg)
p
CC
CT
TT
143/88
167/103
138/83
135/89
166/105
130/84
123/74
150/95
145/90
169/105
139/84
136/89
176/109
136/86
124/74
148/95
142/91
176/105
139/84
136/88
187/128
135/87
125/73
149/95
Valori pressori “office”, cioè misurati in ambulatorio con sfigmomanometro.
242
< 0.05
NS
NS
NS
< 0.01
NS
NS
NS
Michelangelo Sartori et al.
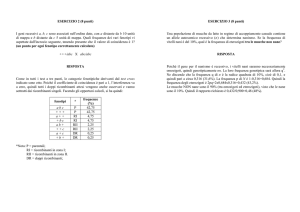
tuali patologie cardiovascolari. Durante un follow-up della durata media di 4.7 anni, i pazienti con allele T raggiungevano più velocemente ed in percentuale maggiore
un grado più severo di ipertensione arteriosa, necessitando quindi di una più precoce terapia farmacologica (Fig.
2)42. Lo studio prospettico e la giovane età della popolazione hanno quindi permesso di dimostrare l’effetto del polimorfismo C825T sullo sviluppo dell’ipertensione.
Come l’allele T favorisca lo sviluppo di ipertensione è
ancora materia di dibattito. Uno studio condotto in una popolazione tedesca ha evidenziato che i pazienti ipertesi portatori dell’allele T presentano un flusso plasmatico renale più elevato rispetto ai pazienti ipertesi con genotipo
CC43. Dal momento che le alterazioni dell’emodinamica
renale sono coinvolte nella patogenesi dell’ipertensione,
il risultato di questo studio suggerisce un possibile meccanismo attraverso cui la presenza dell’allele T potrebbe
contribuire alla regolazione dei valori pressori.
L’allele T non è solo implicato nello sviluppo dell’ipertensione arteriosa ma influenza anche la risposta alla terapia farmacologica antipertensiva. I pazienti neri e bianchi di origine non ispanica con genotipo TT, affetti da ipertensione essenziale, rispondono con un calo pressorio
maggiore alla terapia con idroclorotiazide rispetto ai pazienti con genotipo CC44. Nei giovani soggetti di sesso maschile portatori dell’allele T la clonidina riduce acutamente la pressione arteriosa sistolica e le resistenze periferiche totali in misura maggiore rispetto ai soggetti con
genotipo CC45.
Il polimorfismo C825T non è associato soltanto all’ipertensione arteriosa ma anche ad altre condizioni cliniche che
determinano un maggiore rischio di eventi cardiovascolari e cerebrovascolari. Wascher et al.46 hanno dimostrato la presenza di un’associazione tra allele T ed aterosclerosi carotidea in una popolazione tedesca. Un altro studio condotto recentemente in una popolazione tedesca
ha dimostrato che i portatori dell’allele T presentano un
rischio più elevato di sviluppare patologia coronarica47,
mentre Hengstenberg et al.28 non hanno documentato alcuna associazione tra presenza dell’allele T ed aumentato rischio di infarto miocardico in una popolazione germanica. Infine, in uno studio condotto in una popolazione bianca americana è stata dimostrata una significativa
associazione tra allele T ed ictus ischemico, indipendentemente dai valori pressori48.
Polimorfismo C825T e cardiopatia ipertensiva
Numerosi studi epidemiologici hanno dimostrato come
l’aumento della massa del ventricolo sinistro non sia solo associata all’ipertensione, ma costituisca un importante fattore di rischio cardiovascolare49,50. Nonostante la
pressione arteriosa sia il principale fattore in grado di influenzare la massa del ventricolo sinistro, altri fattori come il sesso, l’età, il peso corporeo e la razza concorrono
a determinarne le dimensioni. Nei grandi studi epidemiologici meno del 50% della varianza della massa cardiaca nella popolazione è spiegata da tali fattori. Una
quota di tale varianza non spiegata potrebbe essere attribuibile a fattori ereditari51.
La cardiomiopatia ipertrofica è una malattia geneticamente determinata, dovuta a mutazione di un singolo gene, tuttavia è difficile ipotizzare che una mutazione di un
singolo gene sia alla base dell’ipertrofia del ventricolo sinistro. Verosimile è la presenza di numerosi polimorfismi
in grado di favorire il rimodellamento del ventricolo sinistro in presenza di ipertensione.
Per quanto riguarda il gene GNB3, in piccole coorti di
pazienti ipertesi è stata documentata un’associazione tra
allele T e cardiopatia ipertensiva52-55. Uno studio tedesco
condotto in una piccola coorte di ipertesi non in trattamento
farmacologico ha dimostrato che l’allele T era associato
a disfunzione diastolica52. Olszanecka et al.53 hanno dimostrato l’associazione tra genotipo TT e ridotto rilasciamento ventricolare in una numerosa coorte di pazienti
di origine russa e polacca, mentre in una piccola coorte di
ipertesi spagnoli in trattamento farmacologico l’allele T
si presentava più frequentemente nei pazienti in cui era presente ipertrofia del ventricolo sinistro54. Semplicini et
al.55 hanno evidenziato che giovani ipertesi di origine
italiana portatori dell’allele T presentano un indice di
FIGURA 2. Probabilità di raggiungere un grado più severo di
ipertensione arteriosa (pressione arteriosa sistolica ≥ 160
mmHg e/o pressione arteriosa diastolica ≥ 100 mmHg, durante il primo anno di follow-up, o pressione arteriosa sistolica
≥ 150 mmHg e/o pressione arteriosa diastolica ≥ 95 mmHg, successivamente al primo anno di follow-up) in pazienti con ipertensione lieve a seconda della presenza (linea continua) o assenza
(linea tratteggiata) dell’allele T. La differenza tra le due curve,
stimate secondo l’algoritmo di Kaplan-Meier, raggiunge la significatività statistica42.
243
Ann Ital Med Int Vol 19, N 4 Ottobre-Dicembre 2004
massa ventricolare sinistra maggiore rispetto ai pazienti
omozigoti per l’allele C. Tali osservazioni sono state estese da Sartori et al.42 in uno studio condotto su 461 pazienti
con ipertensione di grado I, dove si è confermato che i portatori dell’allele T presentano un indice di massa ventricolare sinistra maggiore rispetto ai soggetti con genotipo
CC.
Tutti questi risultati sembrano contraddetti da uno studio trasversale compiuto su una numerosa popolazione tedesca56. In questa popolazione non è stato documentato
nessun rapporto tra geometria cardiaca e polimorfismo del
gene GNB3. Va detto però che la marcata eterogeneità dei
valori pressori, dell’età e del rischio cardiovascolare, nonché il disegno trasversale non permettono di escludere che
l’allele T abbia qualche effetto sulla geometria cardiaca.
ad un maggiore peso corporeo, ad un aumentato rischio
di sviluppare obesità e ad un maggiore spessore della plica tricipitale. Poston et al.63 hanno dimostrato che, in una
popolazione di neri africani e di neri americani, i soggetti portatori dell’allele T presentavano valori maggiori di
indice di massa corporea rispetto ai soggetti omozigoti CC,
indipendentemente dal livello di attività fisica; tale associazione, tuttavia, non raggiungeva la significatività statistica.
L’allele T, secondo uno studio condotto su 294 donne
americane, è associato ad un maggiore aumento di peso
corporeo durante la gravidanza64. Al contrario, uno studio caso-controllo su una popolazione giapponese non
ha documentato nessuna associazione65 e due studi casocontrollo su una popolazione di bianchi europei66 e australiani67 hanno documentato solo una tendenza all’aumento della frequenza del genotipo TT nelle classi di indice di massa corporea più elevato, ma tale tendenza non
raggiungeva la significatività statistica. Similmente, in
una popolazione americana composta da individui bianchi e neri, non è stata riscontrata nessuna associazione tra
il genotipo CT/TT e la massa corporea68.
I risultati contrastanti non sorprendono. Il genotipo
spiegherebbe solo una piccola parte della predisposizione genetica all’obesità e solo in assenza di attività fisica
e di influenze ambientali il polimorfismo predisporrebbe
all’obesità. L’eterogeneità degli stili di vita e delle abitudini alimentari delle popolazioni studiate potrebbero quindi spiegare i risultati positivi in alcuni campioni di popolazione e negativi in altri.
Polimorfismo C825T ed obesità
L’obesità è una malattia cronica, la cui presenza predispone all’insorgenza di diabete mellito, ipertensione, cardiopatie. L’obesità può essere considerata come il risultato dell’interazione di fattori ambientali con il substrato
genetico dell’individuo, in particolare con geni che ne conferiscono la suscettibilità. Circa il 40-70% della variabilità dei depositi di adipe del corpo sembra essere determinata geneticamente18.
Sono stati clonati molti geni responsabili della regolazione del deposito di grasso, ed in particolare quelli delle proteine G eterotrimeriche, che sono risultate importanti
anche per l’adipogenesi. È stato dimostrato che nei topi
la soppressione dell’attività della subunità Gαl2 determina insulino-resistenza57 mentre un aumento d’espressione di questa subunità ha effetti insulino-mimetici58. È
noto che l’ipertensione arteriosa è molto frequente nei soggetti obesi. Ad esempio, a 50 anni l’80% degli obesi ha
intolleranza al glucosio ed è iperteso ed il 67% degli ipertesi è allo stesso tempo diabetico ed obeso59. Il gene
GNB3, codificando per una subunità delle proteine G, è
quindi un “gene candidato” per la patogenesi dell’obesità
e dell’ipertensione. Nell’uomo è stato recentemente dimostrato come gli adipociti dei pazienti obesi con genotipo TT abbiano una ridotta lipolisi dopo stimolo con norepinefrina60, ma i risultati dei numerosi studi di associazione tra il polimorfismo C825T e l’obesità sono contraddittori.
Siffert et al.18 hanno documentato un’associazione tra
allele T ed obesità in soggetti di origine germanica, cinese ed africana. Similmente nei nativi canadesi Nunavut
Inuit, il genotipo TT era associato ad un aumento dell’indice di massa corporea61. Anche un recente studio epidemiologico, condotto da Brand et al.62 su una popolazione belga, ha dimostrato che il genotipo TT era associato
Polimorfismo C825T ed insulino-resistenza
Da anni è stata definita l’esistenza di una sindrome clinica (sindrome metabolica o dell’insulino-resistenza), caratterizzata da insulino-resistenza e iperinsulinemia, in
cui coesistono ridotta tolleranza glucidica o diabete di tipo 2, dislipidemia, ipertensione e obesità69. Poiché numerosi studi hanno dimostrato l’esistenza di un’associazione significativa tra il polimorfismo C825T ed ipertensione od obesità, è plausibile ipotizzare un’associazione
tra questo polimorfismo e l’insulino-resistenza. Poch et al.29
hanno dimostrato la presenza di un’associazione significativa tra insulino-resistenza e polimorfismo C825T in uno
studio condotto su una piccola popolazione di pazienti spagnoli con ipertensione essenziale. Brand et al.62 hanno riscontrato la presenza di insulino-resistenza con frequenza maggiore negli europei con genotipo TT rispetto ai soggetti portatori dell’allele C. Anche Wascher et al.46 hanno dimostrato nei maschi austriaci una maggiore insulino-resistenza nei portatori dell’allele T rispetto ai soggetti
con genotipo CC. Una correlazione statisticamente si-
244
Michelangelo Sartori et al.
gnificativa tra allele 825T e alterata sensibilità all’insulina è stata, infine, rilevata in un recente studio condotto in
una popolazione di pazienti spagnoli con diabete di tipo 270.
Al contrario uno studio condotto recentemente da Nürnberger et al.71 in una piccola popolazione di giovani tedeschi, analizzata per valutare gli effetti metabolici ed emodinamici del carico orale di glucosio, non ha confermato
l’associazione tra presenza dell’allele T e ridotta tolleranza
al glucosio.
so l’insufficienza renale78. Tuttavia, in due successivi
studi di associazione tali risultati non sono stati confermati,
poiché non è stato documentato nessun effetto dell’allele T sulla prognosi sia del trapianto che del ricevente79,80.
Recentemente uno studio in 281 pazienti trapiantati di
rene ha documentato che l’allele T non influisce sulla
precoce perdita del rene trapiantato, ma il genotipo TT interagisce con la pressione arteriosa per portare al declino
della funzionalità renale nel lungo periodo81.
Polimorfismo C825T e nefropatia diabetica
Conclusioni
La patogenesi della nefropatia diabetica è multifattoriale.
Numerosi sono i geni candidati per spiegare lo sviluppo
e la progressione della nefropatia diabetica. I primi geni
ad essere analizzati sono stati quelli deputati alla sintesi
e alla regolazione del sistema renina-angiotensina, dei
recettori e dei meccanismi intracellulari che regolano la
fisiologia della pressione arteriosa72.
Studi prospettici in popolazioni di diabetici hanno riscontrato un aumento di NHE nei pazienti che sviluppavano più facilmente nefropatia73. Poiché l’allele T era
stato inizialmente scoperto nei pazienti con NHE elevato, l’associazione con il gene GNB3 poteva spiegare perché i pazienti con diabete e nefropatia fossero caratterizzati da un’aumentata attività di NHE74.
Sono stati compiuti, quindi, numerosi studi per verificare se il polimorfismo C825T contribuisse allo sviluppo
della nefropatia diabetica, confrontando le frequenze
dell’allele T in pazienti in trattamento dialitico per nefropatia diabetica, in trattamento dialitico per altre nefropatie e in pazienti diabetici senza danno microvascolare75-77. Blüthner et al.75 hanno descritto un aumento
della frequenza dell’allele T nei pazienti tedeschi con
diabete di tipo 2 affetti da nefropatia diabetica terminale.
La frequenza dell’allele T era significativamente più elevata nei pazienti dializzati e diabetici. Questa associazione non è stata però confermata da studi successivi.
Non è stata documentata nessuna associazione nei pazienti americani76 e in una larga coorte di diabetici di tipo 1 di origine polacca e russa77. Nella stessa coorte l’allele T risultava, invece, associato all’ipertensione arteriosa.
Si è indagato anche il ruolo dell’allele T nella progressione verso l’insufficienza renale dei reni trapiantati, che
hanno un genotipo differente da quello del ricevente78-81.
Uno studio preliminare aveva dimostrato che l’allele T nel
donatore era associato ad una prognosi più sfavorevole del
rene trapiantato, confermando l’importanza del GNB3
sulla funzionalità renale78. Era stato infatti documentato
che, indipendentemente dal genotipo del paziente ricevente
il trapianto, i reni dei donatori con genotipo TT, rispetto
ai reni dei donatori CC, progredivano più rapidamente ver-
Nonostante numerosi studi epidemiologici abbiano indagato l’associazione dell’allele 825T con l’ipertensione
arteriosa o le condizioni cliniche ad essa associate, i risultati
sono stati talora contraddittori. Il polimorfismo C825T è
risultato associato ad ipertensione arteriosa ed a più frequente necessità di ricorrere alla terapia farmacologica antipertensiva16,26-30,34,35,42 nelle popolazioni europee, mentre l’associazione tra allele T ed ipertensione non è stata
confermata nelle popolazioni asiatiche, in particolare nei
giapponesi36-38, nei cinesi39, nelle popolazioni di neri
afroamericani40 e di nativi canadesi41. Anche per quanto
riguarda l’associazione tra allele T, obesità, insulino-resistenza, i risultati degli studi di associazione non sono univoci, per cui non vi sono sufficienti evidenze per considerare l’allele T un marker per la sindrome metabolica.
Le ragioni della discrepanza tra gli ormai numerosi studi sono molteplici. Il disegno trasversale e i possibili bias
di selezione possono spiegare i risultati contrastanti di alcuni studi caso-controllo. I criteri di definizione dell’ipertensione, o più in generale del fenotipo clinico, variano da
studio a studio, contribuendo a rendere non confrontabili i differenti studi di associazione. Tali differenze non sono comunque in grado di spiegare risultati contrastanti. Di
maggior rilievo è, invece, l’estrema eterogeneità delle
popolazioni studiate e la mancata definizione dei criteri di
inclusione utilizzati. Infine, nei differenti studi erano inclusi sia pazienti in trattamento farmacologico che in assenza di trattamento e, poiché il genotipo sembra influenzare la risposta alla terapia, gli effetti dell’allele T potrebbero essere stati mascherati. Tutto ciò ha contribuito
a diminuire il potere statistico degli studi sul polimorfismo
del gene GNB3, aumentando il contributo relativo dei
fattori ambientali quali l’età, l’abitudine al fumo, ecc.
Quando si è indagato se questo polimorfismo fosse un
marcatore di rischio per lo sviluppo delle complicanze
dell’ipertensione, ne è stata documentata l’associazione con
la massa indicizzata del ventricolo sinistro42,52-55 e con
l’aterosclerosi carotidea46, tutte condizioni associate ad un
maggiore rischio di eventi cardiovascolari. In uno studio
condotto in una popolazione bianca americana è stata,
245
Ann Ital Med Int Vol 19, N 4 Ottobre-Dicembre 2004
infine, documentata una maggiore associazione tra allele T ed ictus ischemico, indipendentemente dai valori
pressori48. Tuttavia, questi studi preliminari non sono ancora sufficienti per definire il polimorfismo C825T un fattore di rischio cardiovascolare.
Sono quindi necessari ulteriori studi prospettici per chiarire l’importanza dell’allele T sullo sviluppo di eventi cardiovascolari. Se confermato, tale polimorfismo potrebbe
consentire di identificare i pazienti a rischio di sviluppare
ipertensione grave e danno d’organo, individuando la scelta della terapia farmacologica più adatta.
05. Barandier C, Ming XF, Yang Z. Small G proteins as novel
therapeutic targets in cardiovascular medicine. News Physiol Sci
2003; 18: 18-22.
06. Sprang SR. G protein mechanisms: insights from structural
analysis. Annu Rev Biochem 1997; 66: 639-78.
07. Iiri T, Farfel Z, Bourne HR. G-protein diseases furnish a model
for the turn-on switch. Nature 1998; 394: 35-8.
08. Stryer L. Cascate di traduzione del segnale. In: Stryer L, ed.
Biochimica. IV ed. Bologna: Zanichelli, 1996: 401-10.
09. Avkiran M, Haworth RS. Regulatory effects of G protein-coupled receptors on cardiac sarcolemmal Na+/H+ exchanger activity: signalling and significance. Cardiovasc Res 2003; 57: 94252.
10. Wakabayashi S, Shigekawa M, Pouyssegur J. Molecular physiology of vertebrate Na+/H+ exchangers. Physiol Rev 1997;
77: 51-74.
Riassunto
11. Siffert W, Dusing R. Sodium-proton exchange and primary
hypertension. An update. Hypertension 1995; 26: 649-55.
L’ipertensione arteriosa è una condizione patologica
diffusa, di origine multifattoriale, che costituisce un importante fattore di rischio per lo sviluppo di eventi cardiovascolari come l’infarto miocardico e l’ictus. I polimorfismi dei geni che codificano le subunità delle proteine
G eterotrimeriche possono contribuire a spiegare la variabilità interindividuale della pressione arteriosa e la predisposizione allo sviluppo di ipertensione essenziale.
Recentemente è stato descritto un polimorfismo (C825T)
a carico dell’esone 10 del gene GNB3 che codifica la subunità β3 delle proteine G. L’allele 825T è associato a splicing alternativo e alla sintesi di una subunità β3 più piccola
ma più attiva. Vari studi hanno dimostrato che i portatori
dell’allele 825T sono caratterizzati da un aumentato rischio
di sviluppare ipertensione arteriosa, e sembrano rispondere
con un maggiore calo pressorio alla terapia con diuretico
tiazidico e con clonidina. Tuttavia i risultati degli studi di
associazione non sono univoci. L’ipotesi che l’allele 825T
del gene GNB3 sia un marcatore genetico in grado di definire il rischio cardiovascolare dei pazienti ipertesi non ha
avuto ancora la dimostrazione definitiva: sono, quindi,
necessari ulteriori studi per definire l’impatto dell’allele T
sulla prognosi del paziente iperteso.
12. Semplicini A, Ceolotto G, Felice M, et al. Kinetic properties of
erythrocyte Na+-Li+ and Na+-H+ exchange in hypertensive
patients. J Hypertens 1995; 13 (Part 2): 1566-70.
13. Canessa M, Morgan K, Goldszer R, Moore TJ, Spalvins A.
Kinetic abnormalities of the red blood cell sodium-proton exchange
in hypertensive patients. Hypertension 1991; 17: 340-8.
14. Rosskopf D, Fromter E, Siffert W. Hypertensive sodium-proton exchanger phenotype persists in immortalized lymphoblasts
from essential hypertensive patients. A cell culture model for
human hypertension. J Clin Invest 1993; 92: 2553-9.
15. Lifton RP, Hunt SC, Williams RR, Pouyssegur J, Lalouel JM.
Exclusion of the Na+-H+ antiporter as a candidate gene in
human essential hypertension. Hypertension 1991; 17: 8-14.
16. Siffert W, Rosskopf D, Siffert G, et al. Association of a human
G-protein β3 subunit variant with hypertension. Nat Genet
1998; 18: 45-8.
17. Rosskopf D, Koch K, Habich C, et al. Interaction of Gβ3s, a
splice variant of the G-protein Gβ3, with Gγ- and Gα-proteins.
Cell Signal 2003; 15: 479-88.
18. Siffert W, Forster P, Jockel KH, et al. Worldwide ethnic distribution of the G protein β3 subunit 825T allele and its association with obesity in Caucasian, Chinese, and Black African
individuals. J Am Soc Nephrol 1999; 10: 1921-30.
19. Baumgart D, Naber C, Haude M, et al. G protein β3 subunit 825T
allele and enhanced coronary vasoconstriction on α2-adrenoceptor activation. Circ Res 1999; 85: 965-9.
20. Meirhaeghe A, Bauters C, Helbecque N, et al. The human Gprotein β3 subunit C825T polymorphism is associated with
coronary artery vasoconstriction. Eur Heart J 2001; 22: 845-8.
Parole chiave: Genetica; Insulino-resistenza; Ipertensione arteriosa; Ipertrofia ventricolare sinistra; Obesità; Proteine G.
21. Wenzel RR, Siffert W, Bruck H, Philipp T, Schafers RF.
Enhanced vasoconstriction to endothelin-1, angiotensin II and
noradrenaline in carriers of the GNB3 825T allele in the skin
microcirculation. Pharmacogenetics 2002; 12: 489-95.
Bibliografia
22. Naber C, Hermann BL, Vietzke D, et al. Enhanced epinephrineinduced platelet aggregation in individuals carrying the G protein β3 subunit 825T allele. FEBS Lett 2000; 484: 199-201.
01. European Society of Hypertension-European Society of
Cardiology Guidelines Committee. 2003 European Society of
Hypertension-European Society of Cardiology. Guidelines for
the management of arterial hypertension. J Hypertens 2003; 21:
1011-53.
23. Virchow S, Ansorge N, Rosskopf D, Rübben H, Siffert W. The
G protein β3 subunit splice variant Gβ3-s causes enhanced
chemotaxis of human neutrophils in response to interleukin-8.
W Naunyn Schmiedebergs Arch Pharmacol 1999; 360: 27-32.
24. Virchow S, Ansorge N, Rubben H, Siffert G, Siffert W.
Enhanced fMLP-stimulated chemotaxis in human neutrophils
from individuals carrying the G protein β3 subunit 825 T-allele.
FEBS Lett 1998; 436: 155-8.
02. Beevers G, Lip GY, O’Brien E. ABC of hypertension: the
pathophysiology of hypertension. BMJ 2001; 322: 912-6.
03. Hilgers KF, Schmieder RE. Association studies in cardiovascular
medicine. J Hypertens 2002; 20: 173-6.
25. Lindemann M, Barsegian V, Siffert W, Ferencik S, Roggendorf
M, Grosse-Wilde H. Role of G protein β3 subunit C825T and
HLA class II polymorphisms in the immune response after
HBV vaccination. Virology 2002; 297: 245-52.
04. Siffert W. G proteins and hypertension: an alternative candidate
gene approach. Kidney Int 1998; 53: 1466-70.
246
Michelangelo Sartori et al.
26. Schunkert H, Hense HW, Doring A, Riegger GA, Siffert W.
Association between a polymorphism in the G protein β3 subunit gene and lower renin and elevated diastolic blood pressure
levels. Hypertension 1998; 32: 510-3.
polymorphism of the G-protein β3 subunit and antihypertensive
response to a thiazide diuretic. Hypertension 2001; 37: 739-43.
45. Nürnberger J, Dammer S, Mitchell A, et al. Effect of the C825T
polymorphism of the G protein β3 subunit of the systolic blood
pressure-lowering effect of clonidine in young, healthy male subjects. Clin Pharmacol Ther 2003; 74: 53-60.
27. Beige J, Hohenbleicher H, Distler A, Sharma AM. G-Protein β3
subunit C825T variant and ambulatory blood pressure in essential hypertension. Hypertension 1999; 33: 1049-51.
46. Wascher TC, Paulweber B, Malaimare L, et al. Associations of
a human G protein β3 subunit dimorphism with insulin resistance and carotid atherosclerosis. Stroke 2003; 34: 605-9.
28. Hengstenberg C, Schunkert H, Mayer B, et al. Association
between a polymorphism in the G protein β3 subunit gene
(GNB3) with arterial hypertension but not with myocardial
infarction. Cardiovasc Res 2001; 49: 820-7.
47. Von Beckerath N, Schusterschitz Y, Koch W, et al. G protein
β3 subunit 825T allele carriage and risk of coronary artery disease. Atherosclerosis 2003; 167: 135-9.
29. Poch E, Giner V, Gonzalez-Nunez D, Coll E, Oriola J, de la
Sierra A. Association of the G protein β3 subunit T allele with
insulin resistance in essential hypertension. Clin Exp Hypertens
2002; 24: 345-53.
48. Morrison AC, Doris PA, Folsom AR, Nieto FJ, Boerwinkle E.
G-protein β3 subunit and α-adducin polymorphisms and risk of
subclinical and clinical stroke. Stroke 2001; 32: 822-9.
30. Dzida G, Golon-Siekierska P, Puzniak A, et al. G-protein β3 subunit gene C825T polymorphism is associated with arterial
hypertension in Polish patients with type 2 diabetes mellitus. Med
Sci Monit 2002; 8: CR597-CR602.
49. Koren MJ, Devereux RB, Casale PN, Savage DD, Laragh JH.
Relation of left ventricular mass and geometry to morbidity and
mortality in uncomplicated essential hypertension. Ann Intern
Med 1991; 114: 345-52.
31. Brand E, Herrmann SM, Nicaud V, et al. The 825C/T polymorphism of the G-protein subunit β3 is not related to hypertension. Hypertension 1999; 33: 1175-8.
50. Levy D, Garrison RJ, Savage DD, Kannel WB, Castelli WP.
Prognostic implications of echocardiographically determined left
ventricular mass in the Framingham Heart Study. N Engl J
Med 1990; 322: 1561-6.
32. Snapir A, Heinonen P, Tuomainen TP, et al. G-protein β3 subunit C825T polymorphism: no association with risk for hypertension and obesity. J Hypertens 2001; 19: 2149-55.
51. Post WS, Larson MG, Myers RH, Galderisi M, Levy D.
Heritability of left ventricular mass: the Framingham Heart
Study. Hypertension 1997; 30: 1025-8.
33. Buchmayer H, Sunder-Plassmann G, Hirschl MM, et al. Gprotein β3 subunit gene (GNB3) polymorphism 825C—>T in
patients with hypertensive crisis. Crit Care Med 2000; 28:
3203-6.
52. Jacobi J, Hilgers KF, Schlaich MP, Siffert W, Schmieder RE.
825T allele of the G-protein β3 subunit gene (GNB3) is associated with impaired left ventricular diastolic filling in essential hypertension. J Hypertens 1999; 17: 1457-62.
34. Benjafield AV, Jeyasingam CL, Nyholt DR, Griffiths LR,
Morris BJ. G-protein β3 subunit gene (GNB3) variant in causation of essential hypertension. Hypertension 1998; 32: 10947.
53. Olszanecka A, Kawecka-Jaszcz K, Kuznetsova T, et al, for the
European Project on Genes in Hypertension (EPOGH) Investigators. Ambulatory blood pressure and left ventricular
structure and function in relation to the G-protein β3-subunit
polymorphism C825T in white Europeans. J Hum Hypertens
2003; 17: 325-32.
35. Dong Y, Zhu H, Sagnella GA, Carter ND, Cook DG, Cappuccio
FP. Association between the C825T polymorphism of the G protein β3-subunit gene and hypertension in blacks. Hypertension
1999; 34: 1193-6.
54. Poch E, Gonzalez D, Gomez-Angelats E, et al. G-Protein β3 subunit gene variant and left ventricular hypertrophy in essential
hypertension. Hypertension 2000; 35 (Part 2): 214-8.
36. Kato N, Sugiyama T, Morita H, Kurihara H, Yamori Y, Yazaki
Y. G protein β3 subunit variant and essential hypertension in
Japanese. Hypertension 1998; 32: 935-8.
55. Semplicini A, Siffert W, Sartori M, et al. G protein β3 subunit
gene 825T allele is associated with increased left ventricular mass
in young subjects with mild hypertension. Am J Hypertens
2001; 14: 1191-5.
37. Tozawa Y. G protein β3 subunit variant: tendency of increasing susceptibility to hypertension in Japanese. Blood Press
2001; 10: 131-4.
56. Sedlacek K, Fischer M, Erdmann J, et al. Relation of the G protein β3-subunit polymorphism with left ventricle structure and
function. Hypertension 2002; 40: 162-7.
38. Ishikawa K, Imai Y, Katsuya T, et al. Human G-protein β3 subunit variant is associated with serum potassium and total cholesterol levels but not with blood pressure. Am J Hypertens 2000;
13: 140-5.
57. Moxham CM, Malbon CC. Insulin action impaired by deficiency of the G-protein subunit Giα2. Nature 1996; 379: 840-4.
39. Huang X, Ju Z, Song Y, et al. Lack of association between the
G protein β3 subunit gene and essential hypertension in Chinese:
a case-control and a family-based study. J Mol Med 2003; 81:
729-35.
58. Chen JF, Guo JH, Moxham CM, Wang HY, Malbon CC.
Conditional, tissue-specific expression of Q205L Giα2 in vivo
mimics insulin action. J Mol Med 1997; 75: 283-9.
40. Larson N, Hutchinson R, Boerwinkle E. Lack of association of
3 functional gene variants with hypertension in African
Americans. Hypertension 2000; 35: 1297-300.
59. Cerasola G, Semplicini A. Ipertensione e diabete. A cura della
Società Italiana dell’Ipertensione Arteriosa. Ipertensione arteriosa: il contributo della ricerca italiana. Milano: Editrice Kurtis,
2000: 411-29.
41. Hegele RA, Harris SB, Hanley AJ, Cao H, Zinman B. G protein β3 subunit gene variant and blood pressure variation in
Canadian Oji-Cree. Hypertension 1998; 32: 688-92.
60. Ryden M, Faulds G, Hoffstedt J, Wennlund A, Arner P. Effect
of the (C825T) Gβ3 polymorphism on adrenoceptor-mediated
lipolysis in human fat cells. Diabetes 2002; 51: 1601-8.
42. Sartori M, Semplicini A, Siffert W, et al. G-protein β3-subunit
gene 825T allele and hypertension: a longitudinal study in
young grade I hypertensives. Hypertension 2003; 42: 909-14.
61. Hegele RA, Anderson C, Young TK, Connelly PW. G-protein
β3 subunit gene splice variant and body fat distribution in
Nunavut Inuit. Genome Res 1999; 9: 972-7.
43. Zeltner R, Delles C, Schneider M, Siffert W, Schmieder RE. Gprotein β3 subunit gene (GNB3) 825T allele is associated with
enhanced renal perfusion in early hypertension. Hypertension
2001; 37: 882-6.
62. Brand E, Wang JG, Hermann SM, Staessen JA. An epidemiological study of blood pressure and metabolic phenotypes in relation to the Gβ3 C825T polymorphism. J Hypertens 2003; 21:
729-37.
44. Turner ST, Schwartz GL, Chapman AB, Boerwinkle E. C825T
63. Poston WS, Haddock CK, Spertus J, et al. Physical activity does
247
Ann Ital Med Int Vol 19, N 4 Ottobre-Dicembre 2004
not mitigate G-protein-related genetic risk for obesity in individuals of African descent. Eat Weight Disord 2002; 7: 68-71.
73. Ng LL, Davies JE, Siczkoski M, et al. Abnormal Na+/H+
antiporter phenotype and turnover of immortalized lymphoblasts
from type 1 diabetic patients with nephropathy. J Clin Invest
1994; 93: 2750-7.
64. Dishy V, Gupta S, Landau R, et al. G-protein β3 subunit 825C/T
polymorphism is associated with weight gain during pregnancy. Pharmacogenetics 2003; 13: 241-2.
74. Matteucci E, Giampietro O. Sodium/hydrogen exchange activity in type 1 diabetes mellitus: the never-ending story. Diabetes
Nutr Metab 2001; 14: 225-33.
65. Ohshiro Y, Ueda K, Wakasaki H, Takasu N, Nanjo K. Analysis
of 825C/T polymorphism of G protein β3 subunit in obese/diabetic Japanese. Biochem Biophys Res Commun 2001; 286:
678-80.
75. Blüthner M, Schmidt S, Siffert W, Knigge H, Nawroth P, Ritz
E. Increased frequency of G-protein β3-subunit 825 T allele in
dialyzed patients with type 2 diabetes. Kidney Int 1999; 55: 124750.
66. Benjafield AV, Lin RC, Dalziel B, Gosby AK, Caterson ID,
Morris BJ. G-protein β3 subunit gene splice variant in obesity
and overweight. Int J Obes Relat Metab Disord 2001; 25: 77780.
76. Fogarty DG, Zychma MJ, Scott LJ, Warram JH, Krolewski
AS. The C825T polymorphism in the human G-protein β3 subunit gene is not associated with diabetic nephropathy in type I
diabetes mellitus. Diabetologia 1998; 41: 1304-8.
67. Hinney A, Geller F, Neupert T, et al. No evidence for involvement of alleles of the 825-C/T polymorphism of the G-protein
subunit β3 in body weight regulation. Exp Clin Endocrinol
Diabetes 2001; 109: 402-5.
77. Shcherbak NS, Schwartz EI. The C825T polymorphism in the
G-protein β3 subunit gene and diabetic complications in IDDM
patients. J Hum Genet 2001; 46: 188-91.
68. Rankinen T, Rice T, Leon AS, et al. G protein β3 polymorphism
and hemodynamic and body composition phenotypes in the
HERITAGE Family Study. Physiol Genomics 2002; 8: 151-7.
78. Beige J, Engeli S, Ringel J, Offermann G, Distler A, Sharma AM.
Donor G protein β3 subunit 825TT genotype is associated with
reduced kidney allograft survival. J Am Soc Nephrol 1999;
10: 1717-21.
69. Third Report of the National Cholesterol Education Program
(NCEP) Expert Panel on Detection, Evaluation, and Treatment
of High Blood Cholesterol in Adults (Adult Treatment Panel III)
final report. Circulation 2002; 106: 3143-421.
79. Wuthrich RP, Cicvara S, Booy C, Widmer U, Binswanger U.
The 825C/T polymorphism of the G-protein subunit β3 does not
influence blood pressure and renal function in kidney transplant
recipients. Nephrol Dial Transplant 2000; 15: 1663-6.
70. Fernandez-Real JM, Penarroja G, Richart C, et al. G protein β3
gene variant, vascular function, and insulin sensitivity in type
2 diabetes. Hypertension 2003; 41: 124-9.
80. Hocher B, Pleschka A, Yang F, Kraus R, Neumayer HH, Filler
G. G protein β3 subunit 825T genotype is not associated with
differing outcome in pediatric renal transplant recipients. Pediatr
Transplant 2002; 6: 141-6.
71. Nürnberger J, Dammer S, Philipp T, Wenzel RR, Schafers RF.
Metabolic and haemodynamic effects of oral glucose loading in
young healthy men carrying the 825T-allele of the G protein β3
subunit. Cardiovasc Diabetol 2003; 2: 7.
81. Beige J, Kreutz R, Tscherkaschina I, et al. Matrix analysis for
the dissection of interactions of G-protein β3 subunit C825T
genotype, allograft function, and posttransplant hypertension in
kidney transplantation. Am J Kidney Dis 2002; 40: 1319-24.
72. Remuzzi G, Schieppati A, Ruggenenti P. Clinical practice.
Nephropathy in patients with type 2 diabetes. N Engl J Med
2002; 346: 1145-51.
Manoscritto ricevuto il 12.2.2004, accettato il 21.6.2004.
Per la corrispondenza:
Prof. Andrea Semplicini, Clinica Medica 4, Università degli Studi, Policlinico Universitario, Via Giustiniani 2, 35126 Padova.
E-mail: [email protected]
248