A
Mario Graziano
LA MENTE
DEL CONSUMATORE
INTRODUZIONE AL NEUROMARKETING
L'opera usufruisce del contributo dei fondi relativi al Progetto di Ricerca
PRIN anno 2007 dal titolo “Filosofia e naturalizzazione del mentale.
Nuove prospettive sulla conoscenza e la natura umana"
(Unità locale dell'Università degli Studi di Messina).
Copyright © MMX
ARACNE editrice S.r.l.
www.aracne–editrice.it
info@aracne–editrice.it
via Raffaele Garofalo, 133/A-B
00173 Roma
(06) 93781065
ISBN
978-88–548–3201–5
I diritti di traduzione, di memorizzazione elettronica,
di riproduzione e di adattamento anche parziale,
con qualsiasi mezzo, sono riservati per tutti i Paesi.
Non sono assolutamente consentite le fotocopie
senza il permesso scritto dell’Editore.
I edizione:Lussografica, Caltanissetta 2008
II edizione: febbraio 2010
Indice
Introduzione
p.
7
I. Scienze cognitive e neuroeconomia
1.1. Cosa sono le scienze cognitive
11
1.1.1. La neuropsicologia classica, p. 12 - 1.1.2.Le tecniche di
visualizzazione cerebrale, p. 17 - 1.1.3. I limiti delle tecniche, p. 19
1.1.4. La modularità, p. 22 - 1.1.5. Il cervello automatico, p. 25
1.2. La neuroeconomia
28
1.2.1 Utilità e substrato cerebrale umano, p. 31
II. Il Neuromarketing
2.1. Le neuroscienze del consumatore
2.2. La preferenza e il neuromarketing
35
37
2.2.1.La preferenza per le automobili, p. 42 - 2.2.2. Non solo oggetti:
la preferenza elettorale, p. 45
.
2.3. Vendesi emozioni
2.4. Il ruolo della corteccia prefrontale
52
62
III. Il marketing
3.1.Cosa manca al neuromarketing: il marketing
3.2.La percezione
67
69
3.2.1. La percezione subliminale e il marketing, p. 72
3.3. I concetti
75
3.3.1. Domini e frames, p. 78
3.4. Concetti e frames nel marketing
5
82
6
Indice
IV. La ricerca motivazionale
4.1. I bisogni dei consumatori
4.2. Le scale per misurare gli atteggiamenti
4.3. Misure esplicite e misure implicite
89
91
95
4.3.1.Le misure implicite nel marketing, p. 99
4.4. Il potere predittivo delle metafore
103
4.4.1.La metafora nella linguistica, p. 105 - 4.4.2.La teoria
comparativa e quella interattiva, p. 107 - 4.4.3.La metafora
come evento del pensiero, p. 109
4.5. Manager o psicoterapeuta?
114
V. Una possibile sintesi
5.1.Teorie normative e descrittive della decisione
5.2.La teoria dei giochi
5.3.Teoria dei giochi e neuroscienze
5.4. Le emozioni
119
122
125
127
5.4.1. Scienze cognitive ed emozioni, p. 129 - 5.4.2. La valutazione
dell’emozione, p. 131 - 5.4.3. L’ipotesi di polarità, p. 136
5.5. Il ruolo delle emozioni nelle decisioni economiche
5.6.Teoria dei giochi e teoria della mente
139
143
5.6.1. Il test di falsa credenza, p. 145
5.7. ToM e Neuroeconomia
151
Bibliografia
157
Introduzione
Il sogno ricorrente del responsabile marketing di un’azienda è sempre
stato quello di riuscire a prevedere, con precisione, il comportamento
del consumatore di fronte ad una nuova campagna pubblicitaria, ad un
nuovo prodotto; sapere se il nuovo spot ha in sé gli elementi giusti per
funzionare, riuscire ad indirizzare l’azienda senza errori, raggiungere
gli obiettivi preposti con la minima dispersione di risorse. Negli anni
Ottanta, molti autori hanno studiato le condizioni che fanno sì che
un’azienda possa raggiungere con successo questi obiettivi. Ad esempio, Tom Peters e Robert Waterman hanno intervistato 43 imprese con
lo scopo di individuare i fattori determinanti del loro successo. I risultati ottenuti sono stati successivamente presentati nel libro Alla ricerca dell’eccellenza, il libro di marketing più venduto di tutti i tempi.
I due autori riscontrarono come principio determinante, alla base del
successo delle imprese studiate, la precisa attitudine a motivare i propri dipendenti a considerare la qualità ed il valore dei propri clienti.
Nel 1986, Frank Rodgers, direttore centrale marketing dell’IBM, pubblicò il libro Il metodo IBM, in cui viene descritto come questa importante azienda traduca in pratica il principio secondo cui “il cliente è
re”. Tutto ciò che ha formato oggetto della pubblicistica aziendale in
quegli anni negli Stati Uniti, la formulazione delle strategie, lo sviluppo di sistemi informativi per il management, le tecniche di realizzazione, si dimostrò di rilevante importanza poiché si continuò a tenere
conto di una condizione fondamentale: l’orientamento al cliente. Da
questo punto di vista diviene fondamentale chiedersi come i consumatori rispondano ai diversi stimoli di marketing che un’impresa è in
grado di proporre loro.
Un’impresa, infatti, in grado di capire e prevedere correttamente
come i propri consumatori possano rispondere alle differenti versioni
del prodotto, ai diversi livelli di prezzo, a campagne pubblicitarie alternative, riuscirà ad avere un rilevante vantaggio competitivo sui
concorrenti. Nonostante queste considerazioni, nel nostro Paese
l’attenzione verso il comportamento del consumatore è stata posta
solo di recente e decisamente dopo la nascita del marketing propriamente inteso. Mentre negli Stati Uniti, dove è nata questa disciplina,
esiste un certo equilibrio fra gli studi di marketing e quelli sui consu7
8
Introduzione
matori, in Italia in ambito scientifico-disciplinare, questa pratica non è
ugualmente diffusa. Ciò che risulta abbastanza curioso è rilevare come, in Italia, i primi esempi di studi di marketing risalgono ai primi
anni Trenta, mentre le prime trattazioni organiche e sistematiche sul
comportamento del consumatore sono comparse solo nei primi anni
Sessanta. Una spiegazione potrebbe essere data dal fatto che
l’assimilazione delle teorie del comportamento del consumatore, negli
studi di marketing, risulta piuttosto difficile per chi proviene da una
cultura economica-aziendale. Infatti, le radici teoriche delle teorie del
comportamento del consumatore sono abbastanza distanti da una prospettiva d’impresa: antropologia, psicologia, filosofia e scienze cognitive contribuiscono a costruire le basi su cui si innesta un disegno
complessivo che non prende necessariamente in considerazione il
punto di vista delle imprese, quale prospettiva privilegiata. Proprio
partendo da questa istanza, nei primi anni Sessanta, John A. Howard
[Howard, 1963] ha aperto la strada a una disciplina che si è progressivamente affrancata dalla sua subordinazione storica e teorica al marketing management e ha cominciato a produrre teorie e modelli propri,
indipendentemente dal fatto che risultassero immediatamente o necessariamente funzionali o utili per la funzione marketing all’interno
delle imprese. Questa traiettoria sembra culminare nei contributi di alcuni studiosi [Holbrook e Hirscheman, 1982; Hirschman e Holbrook,
1995] che negano espressamente la necessità di finalizzare i risultati
dello studio del comportamento del consumatore ad eventuali applicazioni di marketing: infatti, studiare i consumatori avrebbe un valore in
sé e non solo come supporto per il marketing.
Dagli inizi del Duemila, l’incontro teorico tra ricercatori di diverse discipline, come neuroscienza, psicologia, filosofia, antropologia ed
economia, ha permesso la nascita di due nuove discipline: la neuroeconomia e il neuromarketing. Entrambe si rifanno ai paradigmi e alle
conoscenze delle neuroscienze cognitive e hanno per oggetto lo studio
dei processi mentali espliciti ed impliciti dei consumatori in dei diversi contesti economici, concernenti anche attività di valutazione, di presa di decisione, di memorizzazione o di consumo. Scopo generale,
pertanto, di questo libro è quello di indagare, attraverso la descrizione
e spiegazione dei processi mentali (affettivi e cognitivi), i comportamenti risultanti osservabili negli individui in azione in dei contesti di
Introduzione
9
natura commerciale e consumistica. Il volume è così articolato. Dapprima, nel capitolo I, viene delineato il quadro epistemologico di riferimento nell’ambito del quale si pone la neuroeconomia.
Un’attenzione particolare è stata dedicata alla questione metodologica,
specificando i meriti ed i limiti delle tecnologie di visualizzazione cerebrale. Nel capitolo II segue, attraverso una rassegna di alcuni tra i
lavori empirici più significativi, un’introduzione relativa al dibattito
attorno il neuromarketing. Vogliamo a questo proposito specificare
che quando parliamo di neuromarketing non ci riferiamo ad una versione “debole” che consiste in una semplice appropriazione di metodologie e di tecniche obiettive che mirano a quantificare e visualizzare
dei fenomeni cognitivi, ma al contrario, aderiamo ad una versione
“forte”, secondo la quale è il contesto globale e paradigmatico delle
neuroscienze ad applicarsi a dei contesti particolari circoscritti
dell’attività umana. In questo senso, il neuromarketing contribuisce
alla raccolta di dati sulla conoscenza riguardo la relazione comportamento-cervello, conservando parallelamente una sua finalità pratica
nel mondo degli affari e della pubblicità.
A ciò fa seguito, nel capitolo III, una rassegna delle principali aree
del sapere psicologico applicabili al marketing, in modo da fornire al
lettore una più adeguata comprensione di tutti i fattori che con forza
sfociano in un determinato comportamento d’acquisto. A tal proposito, ci siamo concentrati sulle variabili affettive e motivazionali che
attengono all’individuo, sulle molteplici influenze semantiche, sociali,
antropologiche, valoriali e, più in generale, culturali che, nell’insieme,
connotano i processi d’acquisto. Sono successivamente introdotti, nel
capitolo IV, alcune teorie e modelli che pur trovando origine in altri
ambiti del sapere hanno successivamente trovato collocazione nelle
teorie di marketing. Infine, nel capitolo V, viene riassunto il quadro epistemologico e metodologico di riferimento nell’ambito del quale si
pone la teoria della decisione, ovvero quell’area del sapere che ha per
oggetto lo studio scientifico di come si prendano le decisioni, di come
si possa cercare di renderle ottimali o per lo meno soddisfacenti e di
quali fattori possono influire su di esse. Viene delineato, inoltre, attraverso la trattazione di alcuni concetti della “teoria dei giochi”, come i
vissuti emotivi e le dinamiche strategiche di interazione sottoforma di
10
Introduzione
cooperazione e conflitto possano influire sul processo decisionale individuale.
Questo libro è stato la tappa di un viaggio di conoscenza, di verifica e di confronto, durante il quale tante persone mi hanno accompagnato ed aiutato. In particolar modo dato l’ineguagliabile sostegno
che, in questi anni, ha dato al mio lavoro tengo a ringraziare Pietro
Perconti che ha avuto anche la pazienza di leggere e commentare
l’intero manoscritto. Ringrazio, inoltre, Ninni Pennisi e Franco Lo Piparo per l’influenza che hanno avuto sul mio pensiero con i loro contributi e le loro critiche. Devo rivolgere altresì il mio ringraziamento ai
professori Francesco Parisi, Andrea Velardi, Alessandra Falzone, Daniele Schilirò, Valentina Cardella, Alessio Plebe, per avermi fornito
preziosi commenti e utili stimoli culturali. Ho inoltre beneficiato della
discussione sui temi qui affrontati con molteplici colleghi-amici, tra
cui: Elvira Bruni, Piero Drago, Domenica Bruni, Caterina Scianna,
Sebastiano Nucera, Maria Primo, Emiliano La Licata, Cristina Puleo.
Un debito ugualmente profondo ho con la mia famiglia per avermi
sopportato e sostenuto nei momenti difficili.
I. Scienze cognitive e neuroeconomia
1.1. Cosa sono le Scienze cognitive
Quando si parla di scienza cognitiva s’intende quella confluenza di ricerche multidisciplinari aventi come oggetto di studio i processi cognitivi. Nella scienza cognitiva confluiscono studiosi provenienti da
diversi ambiti quali, ad esempio, filosofia, neuroscienze, psicologia,
linguistica, intelligenza artificiale, per citare solo gli ambiti disciplinari che hanno fornito i maggiori apporti. I settori sono, dunque, molti
ed eterogenei, tanto che si è ritenuto opportuno, probabilmente a ragione, parlare di scienze cognitive, al plurale.
Infatti, da un lato, i vari studiosi pur provenendo da settori diversi
hanno in comune l’interesse per l’analisi della cognizione; dall’altro, a
seconda della provenienza di ogni singolo studioso, tale analisi viene
affrontata con metodi talvolta molto differenti. Questo pluralismo metodologico viene visto da alcuni studiosi come un fattore positivo di
scambio, dialogo e incontro; di contro, secondo altri questo fantomatico pluralismo non è altro che motivo di confusione e di preoccupazione. Cercheremo di affrontare alcune di queste difficoltà epistemologiche ed in particolare ci soffermeremo sull’idea che più di ogni altra è
stata al centro di discussione: cioè l’ipotesi per cui sarebbe possibile
mettere in relazione specifiche regioni del cervello con particolari aspetti del comportamento cognitivo.
Alcuni studiosi, partendo dall’osservazione di sintomi estremamente selettivi a seguito di lesioni cerebrali, hanno postulato che alcune capacità potessero essere realizzate da un insieme complesso di
meccanismi, operanti in modo relativamente autonomo e tali da risultare separabili e singolarmente lesionabili. In questo senso, lo studio
della cognizione e dei suoi rapporti con il cervello si è venuto progressivamente a configurare da un lato come un tentativo di scomporre il
comportamento nei suoi meccanismi più elementari (componente funzionale), dall’altro come un tentativo di individuare regioni distinte
sotto il profilo anatomo-fisiologico nell’organizzazione del cervello
11
12
Capitolo I
(componente neurale), nella speranza di poter individuare omologie
fra gli uni e gli altri.
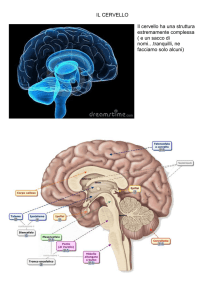
La rilevanza di questo genere di questioni emerge soprattutto nei
confronti dei nuovi paradigmi sperimentali di cui si avvalgono le neuroscienze cognitive per correlare comportamento e cervello, in primis
le metodologie di brain imaging, considerate da gran parte degli studiosi come una delle strategie più promettenti per cercare di restringere reciprocamente l’architettura funzionale dei processi cognitivi e
l’anatomia funzionale del cervello umano. Le ipotesi tradizionali, emerse dai paradigmi sperimentali classici, circa la corrispondenza fra
componenti funzionali ed aree di elaborazione nel cervello, hanno trovato nei metodi di brain imaging un efficace strumento di conferma e
di eventuale revisione.
1.1.1.La neuropsicologia classica
La neuropsicologia, ovvero lo studio dei rapporti fra lesioni cerebrali e
disfunzioni comportamentali, ha rappresentato, a partire dal XIX secolo, il terreno più fertile per lo studio dell’organizzazione funzionale
del cervello umano. Alle origini del paradigma neuropsicologico stava
l’idea che lesioni di particolari regioni della corteccia cerebrale generavano deficit specifici di particolari capacità. La possibilità, quindi, di
identificare delle disfunzioni selettive che lasciavano inalterato
l’insieme delle rimanenti facoltà, ha dato origine alla concezione del
cervello come sistema costituito da organi autonomi specializzati nella
realizzazione di diverse funzioni.
La neuropsicologia si avvaleva, pertanto, di due classi di fenomeni: da un lato anomalie specifiche ricorrenti in un soggetto nell’atto di
svolgere alcune attività (come il nominar un oggetto percepito, il ripetere parole ascoltate, il leggere ad alta voce sequenze scritte di caratteri), dall’altro la presenza nello stesso soggetto di una lesione circoscritta di una determinata area cerebrale. Il primo requisito
dell’indagine neuropsicologica era quello di capire come il deficit osservato fosse interpretabile in maniera significativa rispetto al comportamento normale. La sindrome patologica isolata non dice, infatti, nulla se non in rapporto ad una teoria del funzionamento normale della
capacità considerata. Si richiedeva come prima cosa, pertanto, un mo-
Scienze cognitive e neuroeconomia
13
dello del funzionamento normale di una determinata capacità: soltanto
rispetto ad essa il sintomo poteva essere interpretato come disfunzione
circoscritta di una specifica struttura funzionale. È questo il principio
noto come “assunzione di trasparenza”. Questo principio stabilisce
che schemi comportamentali deficitarii possano essere interpretati solo a patto che rimangano inalterate le restanti capacità cognitive [Caramazza, 1986]. Questa assunzione, in altre parole, richiede che il sistema cognitivo di un paziente che presenta una lesione cerebrale sia
essenzialmente lo stesso di quello di un soggetto normale, eccezion
fatta per una modifica locale del sistema. Viene cioè respinta la possibilità che una lesione cerebrale comporti la creazione di nuove operazioni cognitive, tali da dare luogo nel soggetto ad un diverso sistema
cognitivo che abbia una relazione non trasparente con il sistema originario. Schemi di risposta devianti sarebbero di scarso valore teorico se
fossero privi di restrizioni, se cioè fossero devianti su tutte le funzioni
interessate. L’assunzione di trasparenza postula, quindi, che una lesione cerebrale non alteri complessivamente l’organizzazione dei sistemi
cognitivi, fornendo in tal modo una cornice teorica per la classificazione dei disturbi nonché per la previsione di deficit non ancora osservati. Per meglio esplicitare questa problematica riportiamo, a titolo di
esempio, il modello classico di Ludwig Lichtheim [Lichteim, 1885]
teso a ricostruire l’architettura della facoltà del linguaggio e le procedure legate alla comprensione ed alla riproduzione degli stimoli linguistici:
Figura 1. Modello dell’architettura della facoltà del linguaggio [Lichtheim,
1885]
14
Capitolo I
Questo modello individua l’insieme delle operazioni e delle procedure
richieste dai diversi tipi di compiti linguistici che siamo abituati a
compiere, quali ripetere ad alta voce parole udite, trascriverle e via dicendo. Lo schema di sinistra, infatti, descrive i “centri di elaborazione” del linguaggio e i percorsi dell’informazione di essi (a descrive
l’input sensoriale, m l’output motorio dell’elocuzione verbale, A per il
centro delle rappresentazioni motorie delle parole, O per il centro delle
rappresentazioni visive, B per il centro semantico, E per il centro
d’innervazione degli organi deputati della scrittura). Lo schema di destra, viceversa, riporta i vari punti in cui il sistema è suscettibile di lesioni tali da produrre deficit comportamentali. Come scrive Andrè
Ombredane [Ombredane, 1951], questo modello ha consentito la prima descrizione unitaria delle diverse sindromi descritte nella letteratura afasiologica (ad esempio, da una lesione che colpisce il centro
dell’articolazione motoria M discende la sindrome afasica descritta da
Broca o- secondo la definizione di Wernicke -afasia motoria, tale che
viene abolita la parola spontanea -BMm-, la ripetizione di parole aAm-, la lettura ad alta voce OABMm-).
Il modello di Lichtheim risulta particolarmente interessante per il
nostro studio poiché stabilisce una diretta corrispondenza fra capacità
deficitarie e lesioni selettive di componenti funzionali: l’autore evidenzia, infatti, le connessioni dirette tra i vari centri sottostanti al processo linguistico, ritenendo pertanto che questo fosse il risultato della
loro azione congiunta, e che le diverse forme di afasia fossero in tal
modo causate sia dalla distruzione dei centri, sia dall’interruzione delle vie di connessione tra i vari centri. Naturalmente il modello è stato
oggetto di numerose critiche. John Morton [Morton, 1984], ad esempio, ne dà una sommaria rassegna, a partire da Freud e Head, circa
l’inadeguatezza strutturale dei suoi modelli di spiegazione delle afasie.
In realtà, osserva Morton, il valore dell’opera di Lichtheim sta nel suo
tentativo di separare descrizioni della funzione da descrizione
dell’anatomia, nella sua intenzione di isolare un dominio indipendente
di ricerca, teso a studiare la pura articolazione funzionale dei processi
linguistici alterati in seguito a lesioni. La mistificazione operata dai
denigratori di Lichtheim deriverebbe, sempre secondo Morton,
dall’aver quindi interpretato l’associazione lesione-sindrome come
un’associazione lesione-componente funzionale, dall’aver, pertanto,
Scienze cognitive e neuroeconomia
15
caricato di valenze “localizzazioniste” ipotesi che non erano affatto
nell’intenzione dell’autore.
Ciò che comunque questo modello evidentemente non contempla
è la possibilità di spiegazioni molteplici dello stesso deficit. Tim Shallice [Shallice, 1988] osserva, a tal proposito, il fatto che pur essendo
possibile che alcuni componenti funzionali possa operare in modo
normale, questo non ci impedisce di postulare che il modo in cui essi
interagiscono non lo sia: che non lo sia cioè l’insieme delle procedure
attraverso cui i vari componenti realizzano dati compiti. Shallice suggerisce, quindi, di individuare dei criteri più stringenti per identificare
singoli componenti deficitarii ed evitare in questo modo la costruzione
di modelli artefatti: criteri individuati da Shallice nella strategia delle
“doppie dissociazioni”.
Figura 2. Schema esplicativo del fenomeno della doppia dissociazione [Shallice,
1988]
16
Capitolo I
Ciò che questo modello mette in evidenza è il fatto che il riscontro, rispettivamente, in due soggetti che riportano una lesione cerebrale di
(1) una capacità A deficitaria di fronte a una capacità B illesa e (2)
della stessa capacità A illesa accanto alla capacità B deficitaria, possa
assumersi generalmente sufficiente a garantire l’indipendenza dei due
sistemi responsabili dei deficit osservati. Come risultato degli studi
fatti con il metodo delle doppie dissociazioni, la neuropsicologia cognitiva potrebbe essere descritta come un esercizio di scomposizione
di una funzione cognitiva nelle sue componenti essenziali.
Come sottolinea sempre Shallice, le difficoltà cognitive riscontrate
nei pazienti neurologici devono essere interpretati in termini di un
normale sistema dell’informazione con alcuni sottosistemi isolati o vie
di trasmissione che operano in modo sbagliato. Pertanto questi modelli, non più strettamente legati al substrato anatomico, sono considerati
come caratteristiche schematiche o formalizzate del normale processo
di elaborazione cognitiva dell’uomo, e risultano estremamente utili
per lo studio di pazienti che hanno subito deficit cognitivi a causa di
un danno cerebrale.
La strategia delle doppie dissociazioni ha goduto di tanta stima
negli studi neuropsicologici, rappresentando lo strumento teorico più
comune per la costruzione di modelli normativi di date capacità cognitive. Tuttavia il problema centrale del modello di Shallice, così come
di tutti i modelli che mirano a spiegare capacità complesse, è che se
pur costruito in maniera tale da dar ragione del funzionamento in termini astratti di una data capacità e dei suoi potenziali deficit, sembra
necessitare di un ulteriore requisito e cioè che i processi descritti a livello di architettura funzionale siano ad un certo livello di analisi confrontabili con i processi effettivamente realizzati nel cervello. La neuropsicologia classica per rispondere a questo requisito assumeva che
dovesse esistere almeno una regione cerebrale che fosse la realizzazione fisica di ogni singolo componente funzionale specificato in un
modello e che le regioni fossero tra loro connesse in modo tale da implementare i processi descritti dal modello; questa concezione che
proiettava il tutto sull’anatomia cerebrale è stata oggetto di aspre critiche. In seguito a queste considerazioni anche la moderna neuropsicologia deve continuare a porsi il problema dell’adeguatezza implementazionale dei propri modelli? Un filone consistente di studiosi dichia-
Scienze cognitive e neuroeconomia
17
ra, infatti, di non volersi sbilanciare in attesa di conoscenze più precise
da parte neurologica sulle strutture che realizzano i processi cognitivi.
Esamineremo a tal proposito come le ipotesi fin qui esaminate abbiano
trovato nei metodi di brain imaging un efficace strumento di conferma
o di eventuale revisione.
1.1.2. Le tecniche di visualizzazione cerebrale
I progressi compiuti nella comprensione delle strutture cerebrali si sono fatti significativamente più rapidi a partire dagli anni Settanta, attraverso l’introduzione delle sofisticate tecniche di imagery cerebrale.
L’impiego, infatti, di metodi non invasivi per la visualizzazione
dell’attività cerebrale, quali in primis PET (Positron Emission Tomographi) e fMRI (Functional Magnetic Resonance Imaging ), ha permesso di osservare la sede e l’estensione dei danni cerebrali nonché la
visualizzazione delle modifiche dell’attività cerebrale in seguito allo
svolgersi dei diversi compiti mentali.
La PET (Tomografia per emissione di positroni), è uno strumento
di registrazione indiretta dell’attività cerebrale (mentre le tecniche
come, ad esempio, EEG e MEG, così come le altre tecniche elettrofisiologiche sono definite dirette in quanto misurano direttamente il segnale elettrico o un segnale ad esso riconducibile come quello magnetico), che rivela variazioni di un indice dell’attività cerebrale, nella fattispecie del debito sanguigno dei tessuti neurali. La validità del metodo riposa, sull’assunzione che variazioni dell’attività cerebrale siano
accompagnate costantemente da variazioni dell’irrogazione sanguigna
dei tessuti. Utilizzando un tracciante radioattivo a decadimento rapido,
preliminarmente iniettato nel paziente, è possibile individuare le aree
in cui il debito sanguigno aumenta conseguentemente ad una aumento
di un’attività cognitiva; localizzando, in seguito, le zone in cui il tracciante decade, la PET permette di identificare le aree in cui si è sviluppato il debito sanguigno in seguito ad un’attività neurale.
Il sistema di rilevazione della PET è costituito da una serie di sensori disposti a corona attorno alla testa del soggetto: dal momento che
il decadimento dell’isotopo radioattivo è accompagnato dall’emissione
simultanea di due fotoni in direzione opposte, il rilevamento del loro
incontro simultaneo con i rilevatori permette di risalire alla traiettoria
18
Capitolo I
della loro emissione (rilevamento per coincidenza). Dall’intersezione
delle diverse traiettorie individuate in corrispondenza di altrettante
coppie di fotoni è possibile risalire ulteriormente alle regioni da cui
esse provengono. Una elaborazione digitale di questi dati permette infine di visualizzare tridimensionalmente le regioni d’emissione e di
correggere le eventuali distorsioni dovute all’assorbimento dei tessuti.
La risoluzione spaziale della PET è dell’ordine del millimetro: più che
sufficiente, quindi, a rilevare fenomeni di attivazioni cerebrali di larga
scala. Sotto il profilo temporale, di contro, la PET risulta essere una
tecnica piuttosto povera, dal momento che i tempi necessari per
l’acquisizione delle immagini risultano notevolmente più lunghi della
durata dei processi che essi intendono misurare: l’attività in cui il soggetto è impegnato deve di conseguenza essere mantenuta o ripetuta
per tutta la durata temporale richiesta dagli strumenti ai fini
dell’acquisizione dei dati. Il limite maggiore consiste però nel fatto
che la PET richiedendo l’impiego di traccianti radioattivi risulta essere
una tecnica molto invasiva, con la conseguenza che il numero di esperimenti reiterabili per soggetto è notevolmente limitato.
L’fMRI (Risonanza Magnetica Nucleare funzionale) è, invece,
basata sul fenomeno della risonanza magnetica nucleare, che sfrutta le
proprietà nucleari di certi atomi in presenza di campi magnetici. La
tecnica è entrata in uso negli anni Settanta allo scopo di ottenere immagini dettagliate dell’anatomia cerebrale. Attraverso tecniche di rilevamento ultrarapido dei dati, è divenuta possibile l’acquisizione di
immagini in tempi talmente ridotti (dell’ordine del centesimo di secondo) da permettere di seguire nel loro svolgimento alcuni aspetti del
metabolismo. Applicato alla fisiologia del cervello, l’fMRI ha permesso di visualizzare su una scala temporale estremamente fine le variazioni dell’ossigenazione delle regioni corticali, variazioni che si considerano siano in stretta relazione con il grado di attività delle regioni
stesse. Si sono sfruttate a questo scopo le proprietà magnetiche di cui
godono le molecole di emoglobina, proprietà che differiscono leggermente a seconda che questa sia legata o meno all’ossigeno. Si registrano dunque immagini che si ipotizza rappresentino fedelmente le
variazioni dell’attività neurale regionale laddove si evidenzia un contrasto fra regioni ricche in ossiemoglobina, in cui cioè il flusso sanguigno risulta accresciuto, e regioni dal flusso sanguigno normale.
Scienze cognitive e neuroeconomia
19
Se il segnale, rispetto alla PET, rilevato è più debole, minore la
sensibilità e peggiore di conseguenza il rapporto segnale/rumore, i
vantaggi rispetto a quest’ultima sono molteplici. Oltre alla risoluzione
temporale accresciuta, va sottolineato in primo luogo che l’fMRI si
basa su un segnale intrinseco che non richiede alcun tipo di tracciante.
Inoltre, rispetto alla PET, le misurazioni ottenute per fMRI
dell’attività legate al compito svolto dal soggetto in sede sperimentale
possono essere rapportate direttamente alle immagini anatomiche tridimensionali del soggetto ricavate con la stessa tecnica. L’insieme di
queste tecniche, unitamente a quelle di analisi microstrutturale e agli
studi di lesione, ha aperto uno spazio di indagine estremamente ampio
per studiare l’organizzazione ed il funzionamento del cervello: è oggi
possibile misurare tramite tecniche diverse l’attività neurale dal livello
sinaptico fino a quello convoluzionare, così come studiare fenomeni
che si collocano sotto il profilo temporale intorno a una durata
dell’ordine del millisecondo (le interazioni tra neuroni) fino a fenomeni relativamente lenti, come quelli dell’apprendimento.
1.1.3. I limiti delle tecniche
Con l’introduzione di queste sofisticate tecniche, i modelli della neuropsicologia si sono dovuti confrontare con le varie mappe anatomofunzionali del cervello, divenendo così possibile un raffronto fra teorie, conferme o confutazioni delle stesse. Come le altre metodologie
che mirano a spiegare i complessi rapporti fra cervello e comportamento, anche il brain imaging annovera, all’interno del suo apparato
teorico, una serie di metodi atti a rendere più trasparenti possibile i dati osservativi e a contenere la possibile proliferazione di spiegazioni
dei fenomeni esaminati. A dire il vero nel caso in questione ciò si presenta di fondamentale importanza per il semplice fatto che vengono
applicati modelli comportamentali all’interpretazione di dati di natura
neurofisiologica. La metodologia più accettata in sede neuroscientifica
è quella nota con il nome di “sottrazione cognitiva”. Per capire il ruolo
dei metodi sottrattivi bisogna fare un passo indietro nella storia delle
scienze cognitive fino agli studi di Frans Cornelis Donders di psicologia sperimentale e dei tempi di reazione dei processi mentali. In questi
studi Donders aveva sviluppato una rudimentale tecnica sottrattiva che
20
Capitolo I
nelle intenzioni dell’autore avrebbe permesso di isolare le differenti
operazioni compiute da un soggetto nel corso di una determinata attività cognitiva.
La logica di base del metodo di Donders consisteva nell’idea che
la durata di una fase di elaborazione può essere misurata mettendo a
confronto il tempo necessario per risolvere una versione di un particolare compito (ad esempio premere un pulsante dopo il riconoscimento
di un particolare stimolo visivo), con una seconda versione del compito che differisce dal primo solo perché viene omessa la fase di elaborazione (la pura reazione allo stimolo visivo). La differenza nel tempo
necessario per risolvere le due versioni rappresentano il tempo che è
stato speso nello stadio di elaborazione preso in considerazione. Il metodo sottrattivo di Donders fu ripreso e perfezionato un secolo più tardi da Saul Stenberg con il metodo dei “fattori additivi”. Stenberg
[Stenberg, 1969], infatti, dimostrò che i tempi di reazione dei processi
mentali erano suscettibili di variazioni a seguito di manipolazioni di
alcune variabili: ad esempio, nel caso del tempo necessario ad un soggetto per stabilire se un numero appartenga ad una lista preliminarmente scelta, variabili influenti possono essere la chiarezza con cui il
numero viene presentato visivamente (quindi la chiarezza del segnale),
la ricerca nella memoria attiva della lunghezza della lista memorizzata, l’attivazione della risposta dalla sua compatibilità, ecc.
Quindi assumendo che un task richieda una sequenza di operazioni, è possibile valutare, sostiene Stenberg, in che misura, manipolando
le variabili, ne risenta la durata delle singole operazioni.
L’importazione del metodo sottrattivo di Donders e dei fattori additivi
di Sternberg negli studi di brain imaging è dovuta alla collaborazione
fra il neurologo Marcus E. Raichle e lo psicologo Michael Posner [Posner e Raichle, 1994]. Negli studi dei due autori la sottrazione viene
impiegata nella ricostruzione dei fattori che generano l’attività neurale
rilevata: il principio adottato da Posner ricalca, infatti, quello di Donders e di Stenberg e consiste nel sottrarre, nelle mappe di attivazione
rilevate dalla PET, i valori relativi allo stato di controllo da quelli relativi allo stato attivato.
Ad esempio, sottraendo dai valori ottenuti chiedendo ad un soggetto di fissare uno stimolo visivo, i valori di controllo registrati nello
stesso soggetto mentre tiene gli occhi chiusi, si può ipotizzare, secon-
Scienze cognitive e neuroeconomia
21
do Posner, di poter isolare l’attività cerebrale correlata alla sola fissazione passiva dello stimolo visivo. Quello che vogliamo pertanto affermare è che non si deve ritenere che lo studio della fisiologia o
dell’anatomia funzionale fornisce dati per così dire indipendenti rispetto, ad esempio, ai modelli psicologici di partenza sull’architettura
dei processi cognitivi: in altre parole si vuole avanzare ben più che il
semplice sospetto che gli esperimenti scientifici partano con un modello neuropsicologico ben chiaro.
In complesso, si è riconosciuto che la grande innovazione apportata dall’impiego di tecniche come la PET e l’fMRI, rispetto ad esempio
alle tecniche classiche, consiste nel fatto che, pur non fornendo che
una misurazione indiretta dell’attività cerebrale, esse hanno permesso
di visualizzare in modo tridimensionale e con una notevole definizione spaziale il flusso sanguigno cerebrale, indicatore della variazione
locale dell’attività del cervello. In particolare, le tecniche di brain imaging hanno rappresentato il primo potente strumento non invasivo
per la localizzazione di funzioni cognitive nel cervello umano e sono
state, di conseguenza, un ponte teorico fra gli studi dettagliati sugli animali e le eterogenee informazioni neuropsicologiche sulla specializzazione funzionale del cervello dell’uomo. Le forti aspettative che
molti studiosi nel campo delle scienze cognitive hanno mostrato nei
confronti del brain imaging sono intimamente legate all’idea che esso
potesse per la prima volta fornire una misura integrata dell’attività
globale del cervello umano in relazione a un determinato compito o, in
altre parole, una visualizzazione precisa delle diverse aree cerebrali
coinvolte nella realizzazione di un compito. Al tempo stesso le grandi
riserve sono dovute all’argomento che avanza più di un dubbio
sull’idea avanzata da alcuni studiosi per cui ad ogni procedura cognitiva (modulo funzionale) corrisponda un centro localizzato nel cervello (modulo anatomico).
L’importanza di questa distinzione sta a nostro avviso nel fatto
che una definizione architettonica di modulo deve essere scevra da ingerenze relative alla sua realizzazione neurale: da questa distinzione
dipende il rigore di ipotesi di correlazione e la possibilità di applicare
vincoli comportamentali all’identificazione di aree cerebrali e viceversa. D’altronde, le conoscenze complessive di come le strutture macroscopiche del cervello siano in relazione con gli aspetti del comporta-
22
Capitolo I
mento cognitivo, sono ancora ad uno stadio embrionale. A questo proposito, le scienze cognitive non dispongono che di informazioni locali
provenienti da diversi paradigmi sperimentali: dati circa la risposta di
particolari regioni cerebrali a classi di stimoli sensoriali (evidenza elettrofisiologia), nozioni relative ai percorsi principali all’interno della
corteccia (studi anatomo-fisiologici sulla connettività), evidenza clinica e comportamentale relativa a disfunzioni selettive indotte da lesioni
localizzate (evidenza neuropsicologica), conoscenze comparative
sull’organizzazione cerebrale di specie filogeneticamente affini
all’uomo (studi su primati) oppure informazioni globali sul coinvolgimento di varie regioni e sulle variazioni di parametri metabolici in
corrispondenza di determinati compiti (studi elettrofisiologici su larga
scala e metodologie di brain imaging).
Il problema cardine della modellizazione dei sistemi cerebrali su
larga scala è quello di come impiegare contestualmente e confrontare
reciprocamente questa massa eterogenea di dati sperimentali. Infatti,
lo scarto che intercorre nel rapporto teoria/osservazione fra la neuropsicologia e le neuroscienze non comporta difficoltà rilevanti finché si
resta ad un livello strettamente intrametodologico. Nel momento stesso in cui, però, si passa ad un livello di confronto interdisciplinare in
mancanza di un tessuto teorico comune, diventa fondamentale
l’individuazione di criteri espliciti per il confronto fra teorie ed osservazioni di natura eterogenea. In realtà, se lasciamo cadere alcune delle
assunzioni che governano i singoli metodi di indagine, assunzioni non
sempre universalmente accettate (si pensi, ad esempio, all’assunzione
di trasparenza nel caso della neuropsicologia, o alla pura additività nel
caso del brain imaging), vengono automaticamente a cadere quei vincoli da cui, singolarmente presi, essi dipendono.
1.1.4. La modularità
L’ipotesi che il cervello sia costituito da sistemi operanti in maniera
autonoma e indipendente gli uni dagli altri ha assunto vigore negli ultimi anni del Novecento non solo sotto la spinta delle indagini neuroscientifiche (e quindi dei metodi sottrattivi), ma argomenti a sostegno
sono stati avanzati anche dall’indagine psicologica. Di questi tentativi,
il più rappresentativo è certamente quello di Jerry Fodor [Fodor, 1983]
Scienze cognitive e neuroeconomia
23
che ha esposto le caratteristiche in virtù delle quali alcune proprietà
architettoniche riescano a determinare sottosistemi isolati all’interno
di facoltà più ampie (ad esempio, la facoltà della visione o la facoltà
del linguaggio). Fodor [Fodor, 1985] definisce come “modulo” un
componente di un sistema più complesso caratterizzato da una serie di
proprietà. Tra queste proprietà la più importante è la specificità di dominio, vale a dire quella caratteristica che intende valutare la destinazione di un sistema nella elaborazione dell’informazione proveniente
da una sola modalità sensoriale. Questa caratteristica esclude, pertanto, che lo stesso componente funzionale possa trattare input diversi dal
dominio per cui è designato.
Strettamente legata alla specificità di dominio vi è anche una altra
proprietà, l’idea di Fodor più forte della nozione di modulo, e cioè
l’icapsulatezza o “ impenetrabilità cognitiva”. Un sistema si definisce
“ incapsulato” se non riceve informazioni da altri sistemi, ovverosia se
i processi che svolge non sono modulati dall’insieme di informazioni
di cui l’organismo dispone, ma dipendono bensì soltanto
dall’informazione di livello più basso (i cosiddetti transducers) o
dall’informazione contenuta nel sistema stesso. L’esempio riportato
dallo stesso autore è quello della visione, in cui si può cercare di stabilire in che misura gli stadi iniziali della percezione visiva non siano interessate da conoscenze preacquisite, ma svolgono autonomamente la
loro elaborazione degli stimoli. In altre parole, le operazioni di un
modulo si avvalgono unicamente di informazioni altamente specifiche
contenute al suo interno, senza mai ricorrere a conoscenze di tipo più
generale disponibili al resto del sistema cognitivo. Il modulo fodoriano è pertanto computazionalmente autonomo, vale a dire riesce a portare a termine il processo di cui è responsabile in modo completamente autonomo, senza condividere cioè risorse con altri componenti o
meccanismi neurali. Oltre alla specificità di dominio, l’accessibilità
limitata e l’incapsulamento informativo, altre proprietà definiscono un
modulo fodoriano: ad esempio, i moduli sono veloci (cioè non subiscono interferenze da altri sistemi tali da rallentare il processo), sono
obbligati (cioè che non è possibile, ad esempio, percepire una frase
come una pura sequenza di suoni), mostrano disfunzioni peculiari (caratteristica legata alla specificità di dominio e di fondamentale impor-
24
Capitolo I
tanza per l’indagine psicologica), e seguono nel loro sviluppo un ritmo
e una sequenza caratteristica.
Queste caratteristiche sono una conseguenza diretta del modo in cui
i moduli sono concepiti, cioè come meccanismi di elaborazione di informazione puramente sintattica, tarati, quindi, sul particolare formato
dei propri dati e non sul contenuto. Impostando in questo modo il concetto di modularità, rimangono fuori dalla spiegazione di Fodor stati
mentali complessi (come, ad esempio, credenze e desideri) che non
possono essere ricondotti facilmente ad una singola funzione mentale,
ma che sono invece emergenti, attribuibili olisticamente alla mente
nella sua interezza e non a un suo sottosistema: in opposizione alle capacità implementate localmente dai moduli mentali, Fodor tratta questi stati come globali. La sensibilità di essi al contenuto, oltre che alla
forma, lo spinge a vederli come non implementabili all’interno di un
modulo e, quindi, non trattabili da un punto di vista computazionale.
Contro questa ipotesi si sono schierati i sostenitori
dell’interpretazione massiva del modularismo mentale, secondo i quali
la cognizione può essere spiegata facendo appello all’idea di un funzionamento coordinato dei numerosi moduli specializzati, deputati
ciascuno di essi a trattare problemi di un dominio specifico, mentre le
proprietà della cognizione generale sarebbero da ricondursi
all’interazione tra i moduli [Pinker, 1997; Sperber, 1994].
Entrambe le interpretazioni del modularismo sostengono, quindi,
che le informazioni sono elaborate dai moduli da un punto di vista puramente sintattico, secondo dei processi perfettamente replicabili da
una macchina di Turing. Per Fodor e per i sostenitori della teoria modulare classica della mente, tuttavia, i dati da elaborare ricalcano le entità teoriche della psicologia del senso comune e sono codificati in un
linguaggio interno (il mentalese) simile ad un linguaggio naturale, in
maniera tale che la base di conoscenze a fondamento dei moduli ricalca in senso forte un processo logico-deduttivo di calcolo. Secondo i
teorici della modularità massiva, al contrario, le informazioni sono
presenti nei moduli in un formato subsimbolico [Smolensky, 1988],
dando in questo modo la possibilità ai moduli di interagire strettamente tra loro: infatti, sebbene essi trattino forme di informazioni altamente specifiche, la codifica di queste nei circuiti neurali può dar vita a dei
Scienze cognitive e neuroeconomia
25
meccanismi di corrispondenza e di associazione che ignorano la specificità dei dati e del loro formato di rappresentazione.
Steven Pinker, a questo proposito, scrive: “i moduli mentali non
hanno bisogno di essere rigidamente isolati l’uno dall’altro, di comunicare solo tramite poche e strette condutture..[…] la metafora del
modulo mentale è insomma un pò approssimativa; una migliore è
quella di Chomsky, di organo mentale che è una struttura fatta su misura per svolgere una particolare funzione” [Pinker, 1997: 9]. Da questo deriva che non è necessario ipotizzare, in maniera alquanto ingenua, l’esistenza di moduli per ogni argomento che gli uomini possono
trattare, ma, al contrario, qualsiasi fenomeno di distribuzione sociale
dell’informazione può utilizzare i preesistenti moduli.
Argomentazioni a favore della modularità massiva della mente
provengono dallo studio delle patologie psicologiche: l’autismo, per
citare un solo caso, è stato recentemente interpretato come un danneggiamento locale dei moduli deputati all’interpretazione degli stati
mentali altrui (e propri) in termini di credenze e desideri e
all’attribuzione di questi stati ad altri (e a se stessi) che, tuttavia, lascia
intatti altri tipi di abilità cognitive [Baron-Cohen e Swettenham,
1997]. Più dubbio è il discorso riguardo all’evidenza sperimentale:
l’ipotesi della modularità è in effetti confermata da analisi
dell’attivazione neurale per quanto riguarda alcune attività cognitive,
quali la percezione [Zeki, 2000], ma per le attività generalmente definite come “centrali”, l’interpretazione dei dati empirici è più controversa [Carruthers, 2003]. Infatti, sebbene, per una definizione minima,
i moduli devono essere pensati come distinti e specializzati, la loro
funzione dipende in larga misura dalla posizione rispetto a sistemi di
moduli più ampi e infine al macrosistema rappresentato dal cervello
umano, la cui complessa topografia siamo ben lontani dal conoscere
con sicurezza.
1.1.5. Il cervello automatico
Per molto tempo si è pensato, in modo implicito, che gli esseri umani
avessero capacità cognitive generali e che queste potessero essere applicate a qualsiasi tipo di problema. Inoltre si assumeva che le persone
si comportavano in maniera equivalente nei confronti dei problemi
26
Capitolo I
che si presentavano loro con le stesse caratteristiche. L’automaticità
del cervello, evidenziata da alcuni studi neuroscientifici, suggerisce, al
contrario, che il modo di agire delle persone dipenderà, in maniera
critica, da come può essere valutato un particolare problema da un
modulo neuronale specializzato e che ben si adatta a quel tipo di analisi. Pertanto, quando esiste un modulo neuronale specializzato a risolvere un particolare compito, l’elaborazione avviene rapidamente ed
il compito viene svolto relativamente senza sforzi (è quanto avviene,
ad esempio, per i processi automatici implicati nella visione). Sapere,
quindi, fino a che punto i nostri giudizi, le nostre decisioni o i nostri
comportamenti sono sotto il controllo del pensiero cosciente o se sono
frutto di processi automatici è tra le questioni al centro di un acceso
dibattito. Inizialmente era dibattuta l’esistenza stessa di questi particolari processi. Oggi, invece, la discussione si focalizza sulle circostanze
nelle quali l’uno o l’altro di questi processi (controllati o automatici)
sono attivi.
I “processi controllati” vengono definiti come stati di atti mentali
che vengono iniziati volontariamente, che necessitano pertanto di uno
“sforzo” e che si possono controllare. Definire, invece, i “processi automatici” è più arduo. In una accezione minima essi sono i contrari dei
processi controllati, operano in parallelo alla coscienza, non necessitano di alcuno “sforzo intenzionale” e sono abitualmente vissuti come
percezioni [Lieberman, 2003]. Bargh e Chartrand [Bargh e Chartrand,
1999] distinguono due forme di processi mentali “non coscienti”. Entrambi operano senza sforzo ma il primo sistema necessita del comando della coscienza per iniziare la sua azione (ad esempio, guidare una
automobile per la centesima volta), mentre l’altro evolve completamente in parallelo. Il modo di operare dei processi controllati e automatici è chiaramente interattivo anche se, naturalmente, i processi automatici essendo più rapidi fanno sempre valere la loro “opinione”,
occupando in tal modo larga parte delle basi del comportamento [Camerer et al., 2003]. Di fronte ad un compito o un problema da risolvere, i due processi possono arrivare ad una diversa soluzione. Ciò è stato dimostrato da Stevan Sloman [Sloman, 1996], il quale evoca delle
forme di credenza che possono essere simultaneamente contraddittorie
come viene provato dall’illusione visiva e l’errore di giudizio delle
due linee dell’illusione ottica di Muller-Lyer.
Scienze cognitive e neuroeconomia
27
Figura 3. L’illusione di Muller-Lyer
Pur essendo le linee della stessa lunghezza (cambia solo il senso) la
percezione visiva e la comprensione astratta forniscono due risposte
differenti. Solo la conoscenza astratta delle regole della geometrica ci
permettono di comprendere che la percezione visiva induce all’errore.
Un altro errore di giudizio che ci permette di illustrare la coesistenza
di processi controllati e automatici è conosciuto con il nome di “errore
di congiunzione” [Tversky e Kahneman, 1983] e si rifà al più noto e
semplice tra i principi statistici, ovvero quello che la probabilità di una
congiunzione è inferiore alla probabilità del verificarsi di ciascuna delle sue componenti ( la formula è: P (A e B) < P (A) e P (a e B) < P
(B)).
Tversky e Kahneman presentando l’esperimento ai partecipanti
premettono loro che:
Linda ha 31 anni, è molto intelligente ed ha ottenuto una laurea in filosofia. Inoltre, è impegnata socialmente contro le discriminazioni ed
ha partecipato a delle dimostrazioni anti-nucleari.
In seguito i due studiosi domandarono ai partecipanti di classificare in
ordine di probabilità alcune opzioni di risposte, tra cui:
1) Linda è attiva nel movimento femminista (A)
2) Linda è cassiera in una banca (B)
3) Linda è cassiera in una banca ed è attiva nel movimento femminista
(A e B).
La stragrande maggioranza dei partecipanti, anche quelli che avevano delle ottime conoscenze dei principi statistici, violò la regola
di congiunzione: essi non la applicarono semplicemente perchè si trovavano fuori da un contesto di un corso normale di statistica. La coe-
28
Capitolo I
sistenza di queste due forme di valutazione è, secondo gli autori, la
prova dell’esistenza di due processi, uno di deliberazione attentiva e
l’altra fatta su base associativa.
Studi neuroscientifici, nei quali si è utilizzato l’fMRI, hanno permesso di identificare alcuni corollari cerebrali dei processi controllati
e automatici. Il neuroscienziato Philip Lieberman classifica i processi
in due categorie: il “sistema C” ed il “sistema X”. Il “sistema C” è
composto da tre regioni cerebrali: la corteccia cingolata anteriore, la
corteccia prefrontale ed il lobo temporale mediale (incluso
l’ippocampo). Il lobo temporale mediale permette la memoria cosciente. La corteccia prefrontale (chiamata anche regione esecutiva) utilizza
le informazioni provenienti dalle altre zone del cervello per formulare
obiettivi a breve e lungo termine, pianificando le azioni conformemente a questi obiettivi [Lieberman, 2003; Shallice e Burgess, 1998; Camererer et al., 2003]. La corteccia cingolata anteriore si attiva per diversi conflitti cognitivi [Eisenberger et al., 2003]. Questa regione può
“allertare” la corteccia prefrontale quando i processi automatici non
raggiungono il loro obiettivo [Botvinick et al., 1999]. Benché queste
regioni fanno parte dei processi controllati non si può classificarli
troppo rigidamente poiché alcuni processi automatici hanno ugualmente corso in queste strutture cerebrali. Per quanto riguarda il “sistema X”, Lieberman, individua tre strutture neuronali: l’amigdala, il
giro centrale (gangli della base) e la corteccia laterale temporale.
L’amigdala reagisce spontaneamente a degli stimoli di differenti forme (espressioni di paura o presenza, ad esempio, di animali che incutono timore, ecc.) dando vita a movimenti motori di avvicinamento o
di fuga. Il giro centrale risponde a differenti forme di ricompensa anticipata, mentre la corteccia temporale laterale permette il riconoscimento dell’identità, attributi e comportamenti di oggetti sociali e non
sociali.
1.2. La neuroeconomia
Gli economisti sono stati i primi, proponendo il termine neuroeconomia, a cogliere l’esigenza di comprendere i processi di decisione degli
agenti economici rifacendosi agli approcci della psicologia cognitiva e
Scienze cognitive e neuroeconomia
29
delle neuroscienze [Tversky e Kanheman, 1974]. La neuroeconomia è
una branca dell’economia comportamentale (behavioral economics)
che cerca di indagare il ruolo dei meccanismi psicologici dell’analisi
economica senza, tuttavia, rigettare di colpo il paradigma neoclassico,
che fornisce una cornice teorica basata sulla massimalizzazione
dell’utilità, degli equilibri e dell’efficienza [Camerer e Loewenstein,
2002]. Questi concetti sono stati rimpiazzati, comunque, da altre ipotesi giudicate più realiste, al fine di tenere conto dei limiti umani in
materia di capacità di calcolo, volontà, egoismo. È in questo contesto
che la neuroeconomia è stata definita come lo stato in cui
l’utilizzazione dei processi cerebrali permette di trovare dei nuovi
fondamenti per le teorie economiche [Camerer, 2004]. L’economia
sperimentale, la psicologia e le neuroscienze possono, pertanto, convalidando o falsificando il modello dell’utilità, permettere d’arricchire
la comprensione del comportamento economico. Nel modello
dell’utilità, si suppone che la scelta presa abbia una più grande utilità
rispetto le sue alternative concorrenti. L’utilità può, dunque, essere
concepita come un sistema mentale in cui si effettua una valutazione
soggettiva, delle differenti alternative, con l’ipotesi della massimizzazione. Per Jeremy Bentham, è la dimensione edonistica dell’utilità, il
cuore della presa di decisione. La scelta tende ad orientarsi verso la ricerca del piacere e l’evitabilità del dispiacere. Di recente, sono stati attivati dagli studiosi di neuroscienze nuovi campi di studio. Queste ricerche, collegate alla memoria, ai processi di decisione e di risoluzione dei problemi, fanno appello ad alcuni concetti proposti
dall’economia, in primis l’utilità [Camerer e Loewenstein, 2002].
L’utilizzazione del concetto di utilità per spiegare le scelte ed i comportamenti complessi ha quindi permesso di stabilire alcuni ponti tra
neuroscienza ed economia.
L’ipotesi standard di una funzione d’utilità consiste nell’idea che
l’utilità sia marginale e decrescente, vale a dire che U<0 e U>0. Un
esperimento di Platt e Glimcher [Platt e Glimcher, 1999] fatto su delle
scimmie, ha permesso di illustrare come il cervello possa realmente
codificare questo genere di funzione. Infatti, vi è una certa similitudine tra come le scienze economiche intendono questo concetto di base
e i dati che sono state registrati a partire dal cervello delle scimmie. Lo
studio sperimentale di Platt e Glimcher [Platt e Glimcher, 1999] ha
30
Capitolo I
permesso, utilizzando il concetto di speranza dell’utilità e la tecnica di
misurazione di un solo neurone, di dimostrare che la regione laterale
intraparietale del cervello delle scimmie, possa allo stesso tempo codificare sia le probabilità sia il valore associato ad una ricompensa.
L’esperimento consisteva nell’insegnare alle scimmie a scegliere tra
due punti luminosi (che apparivano su degli schermi piazzati a destra
o a sinistra rispetto al punto di osservazione in cui si trovavano le
scimmie). Quando, con un movimento della testa, le scimmie effettuavano la scelta giusta, ottenevano una ricompensa (del cibo). Per rendere massima la sua utilità, una scimmia doveva ricordare le probabilità
anteriori associate alla sua scelta ed ovviamente anche il valore della
ricompensa. Il compito sperimentale, veniva eseguito su blocchi di
100 compiti. In alcuni di questi blocchi, la probabilità del movimento
verso la parte destra si stabilivano all’ 80% ed al 20% verso la sinistra.
In altri blocchi, queste probabilità erano invertite. In questo modo, le
probabilità anteriori e posteriori, a ognuno di questi blocchi dovevano
essere continuamente stimate dal cervello della scimmia e non essere,
quindi, concordanti.
Figura 4. L’esperimento di Platt e Glimcher [Platt e Glimcher, 1999]
L’obiettivo era quello di variare le probabilità di ricompensa mantenendo costanti gli stimoli visivi e i movimenti motori, in maniera tale,
da verificare se l’attivazione dei neuroni della regione laterale intrapa-
Scienze cognitive e neuroeconomia
31
rietale era correlata, in un modo o in un altro, con le probabilità anteriori o posteriori. Mantenendo costanti gli stimoli e i movimenti delle
scimmie, gli autori constatarono che le probabilità di ricompensa ed i
loro cambiamenti erano correlati all’attivazione neuronale. Quindi, il
cervello delle scimmie, codificava un segnale non sensoriale e non
motore. Platt e Glimcher interpretarono questo segnale come prova
che le scimmie cercano innanzitutto di massimizzare le loro probabilità di guadagno.
Dopo aver mostrato che il cervello della scimmia può codificare le
probabilità associate ad una ricompensa, un secondo momento
dell’esperimento era dedicato a verificare se la scimmia è capace di fare altrettanto con il valore della ricompensa. Gli autori, quindi, mantennero costanti i movimenti, gli stimoli e le probabilità di ricompensa
(che furono fissate al 50%) e variarono, da un blocco di compiti
all’altro, solo le quantità di ricompense (in alcuni blocchi, le quantità
di cibo ottenuto erano 0,2 ml guardando a sinistra e di 0,1 ml guardando a destra; in altri blocchi, le quantità erano invertite). I risultati
misero in luce come anche i valori del guadagno, quando gli elementi
sensori-motori erano mantenuti costanti, venissero codificati dai neuroni; in pratica, i neuroni erano più attivi quando la speranza di guadagno era elevata. Il fatto che, il cervello delle scimmie, fosse in grado, allo stesso tempo, di codificare tanto l’ampiezza delle ricompense
quanto la loro probabilità, dimostra due cose. In primo luogo, che il
concetto di speranza dell’utilità, sembra avere trovato un corollario
neurologico. In secondo luogo, che alcuni concetti la cui base si trova
nella teoria economica possano trovare spazio nel quadro d’analisi
delle neuroscienze. L’economia e le neuroscienze possono, quindi,
beneficiare l’una dell’altra, cosa che del resto è stata già sottolineata
da Knutson e Peterson [Knutson e Peterson, 2005].
1.2.1. Utilità e substrato cerebrale umano
Comprendere meglio i meccanismi grazie ai quali il cervello valuta e
confronta delle alternative e delle forme variabili di ricompense ci può
permettere di risalire alle determinazioni delle nostre scelte e dei nostri comportamenti. Nel cervello, ogni alternativa deve essere incorporata e confrontata in modo automatico o delimitata, a partire da dei ve-
32
Capitolo I
ri domini di informazione. Queste alternative devono dunque essere
trasformati dal cervello in unità comparabili. Per esempio,
l’attivazione della corteccia parietale posteriore è correlata
all’ampiezza del guadagno monetario [Paulus et al., 2001] tanto che la
sua anticipazione sarà positivamente correlata con l’attivazione della
regione dello striato ventrale [Knutson e Peterson, 2005]. Due osservazioni importanti sono state fatte da Knutson e Peterson. In primo
luogo, lo striato ventrale è attivato solo dall’anticipazione di un guadagno monetario; un guadagno, una perdita, o l’anticipazione di una
perdita monetario non hanno, apparentemente, effetto su questa regione.
Il quadro d’interpretazione di questi autori (che diverge parzialmente da quello di Platt e Glimcher) si inserisce nell’ottica della teoria
prospettiva di Kahneman e Tversky [Kahneman e Tversky, 1984], vale a dire che la prospettiva di guadagno e di perdita sono supposte non
essere trattate dagli stessi algoritmi e dagli stessi meccanismi neurali.
Una seconda osservazione di Knutson e Peterson concerne la regione
mediana della corteccia prefrontale. Secondo loro, questa regione registra il guadagno monetario, disattivandosi quando il guadagno monetario è nullo. Fatto importante è che questa non è attiva né per quel
che concerne le anticipazioni (perdita o guadagno), né le perdite. La
corteccia prefrontale non è, quindi, funzionale all’anticipazione di una
ricompensa inizialmente appresa. La corteccia prefrontale può tuttavia
dare dei feedback quando le condizioni ambientali cambiano. Brian
Kuntson ed i suoi colleghi [Knutson et al., 2001] fanno la seguente
omologia: in pratica paragonano lo striato ventrale ad un pedale, mentre la regione mediana della corteccia prefrontale ad un volante che
può orientare le scelte, verso la direzione appropriata, quando cambiano le condizioni. I risultati ottenuti, oltre a convalidare l’esistenza di
due diversi tipi di processi mentali della presa di decisione (i processi
controllati dalla corteccia prefrontale ed i processi automatici dello
striato), danno forza al concetto che Daniel Kahneman [Kahneman,
1994] ha suggerito per l’utilità; cioè che le differenti forme d’utilità
trovano, rispettivamente, la loro fonte da differenti regioni del cervello. Secondo Knutson e Peterson, lo striato ventrale è legato al concetto
d’utilità anticipato, mentre la regione mediana della corteccia prefrontale è legata al concetto di utilità istantaneo. I loro risultati sono, inol-
Scienze cognitive e neuroeconomia
33
tre, ugualmente compatibili con la distinzione fatta da Berridge e Robinson [Bernheim e Rangel, 2002] secondo i quali esistono due distinti processi neurali attivi per quello che si ama (liking) e quello che si
vuole o si desidera (wanting).
È possibile, inoltre, stabilire alcuni parallelismi tra il modello
dell’utilità e la dopamina (composto chimico che dà origine alla noradrenalina; la dopamina è uno dei neuromediatori e la sua ridotta presenza, in alcuni nuclei del cervello, è messa in relazione al morbo di
Parkinson). Numerose ricerche, infatti, hanno dimostrato il ruolo primordiale giocato dalla dopamina nella selezione degli obiettivi. Le
prime evidenze del ruolo giocato dalla dopamina, nella presa di decisione, associano quest’ultima alla sensazione di piacere [Olds e Milner, 1954]; gli studi, infatti, mostrano l’aumento della dopamina in
certe regioni del cervello dei topi, quando questi venivano coinvolti in
delle attività gratificanti. Da questo sembra evidente il legame tra la
dopamina ed il piacere edonistico. Riassumendo, possiamo asserire
che, per assicurarsi la sopravvivenza, tutte le specie devono potere
compiere delle funzioni vitali diverse come cibarsi, reagire alle aggressioni e riprodursi. I circuiti cerebrali della ricompensa permettono
di ottenere questi obiettivi. L’area ventrale (ATV), un gruppo di neuroni situati nel pieno centro del cervello, è particolarmente importante
nel funzionamento di questo circuito. Questa riceve degli imputs da
diverse altre aree del cervello che la informano del livello di soddisfazione dei bisogni fondamentali (o più specificatamente umani).
All’arrivo di un segnale che annuncia una ricompensa, si riscontra un
aumento dell’attività della ATV. Questa regione trasmette, quindi,
queste informazioni grazie ad un messaggio chimico particolare, la
dopamina appunto, che viene emessa dalla corteccia prefrontale e
dall’amigdala, attivando in questo modo sia l’attività motoria sia
l’attenzione selettiva.
Dunque, è questa secrezione di dopamina ad ottenere l’effetto di
rafforzare determinati comportamenti, permettendo la soddisfazione di
bisogni fondamentali. Di conseguenza, possiamo affermare che il circuito della ricompensa, così come quello della punizione, forniscono
la motivazione necessarie alla maggioranza dei nostri comportamenti.
La secrezione della dopamina può essere causata dall’ambiente associato alla ricompensa e non dalla ricompensa stessa. Per esempio, la
34
Capitolo I
semplice vista di un bicchiere di vodka può generare un impulsivo desiderio (non cosciente) di fumare una sigaretta ad una persona che, in
passato, aveva preso l’abitudine di consumare insieme queste due sostanze. La dopamina sarà allora responsabile di un’insieme di comportamenti, fra cui anche l’acquisto economico [Laibson, 2001; Bernheim
e Rangel, 2002], destinati ad attendere diverse forme di ricompensa.