NEUROBIOLOGIA DELLA MEMORIA
Sommario: 1. Introduzione – 2. Aspetti Psicologici della Memoria – 3. Anatomo–Fi-siologia della
Memoria – 4. Mediatori Chimici della Memoria – 5. Neuropeptidi e Pla-sticità – 6. L’Engramma – 7.
Sedi della Memoria – 8. Tipi di Memoria – 9. Apprendimen-to – 10. Alterazioni della Memoria – 11.
Amnesia e Stati Dissociativi – 12. La Memoria dell’anziano – 13. Demenza – 14. Conclusioni.
Bibliografia.
1. INTRODUZIONE
Ma dove avrò messo quei documenti? Dove sono finiti i miei occhiali? Il nome di un attore?
Dove abbiamo lasciato la macchina? La data di un appuntamento? Come ho potuto dimenticare
quell’impegno? Quante volte capita di porgersi queste domande? Cosa disturba il ricordo? Cosa
impedisce di utilizzare informazioni che pure si possiedono? C’è qualche dato che sarebbe utile
ricordare ma che non si riesce a richiamare alla memoria? Il compito che ci si propone è di dare qualche
risposta a queste ed altre domande.
La memoria può essere considerata come il meccanismo che permette di fissare, conservare e
rievocare esperienze ed informazioni acquisite dall’ambiente (interno ed esterno) e, nell’uomo, derivate
anche dal pensiero e dalle emozioni.
La memoria è la capacità di un organismo vivente di conservare tracce della propria esperienza
passata e di servirsene per relazionarsi al mondo e agli eventi futuri. La funzione in cui si esprime la
memoria è il ricordo, la cui diminuzione o scomparsa determina l’oblio. Come fenomeno normale che
descrive la fase discendente di ogni processo mnemonico, l’oblio va tenuto distinto dall’amnesia, che è
un fenomeno patologico.
La memoria è una funzione fondamentale della mente umana, senza il suo coinvolgimento,
infatti, non potrebbero operare con successo le funzioni psichiche superiori, come la percezione, il
riconoscimento, il linguaggio, la pianificazione, la soluzione di problemi o il prendere decisioni.
Le funzioni psichiche, che comprendono anche l’attenzione, la coscienza, il pensiero, la critica,
l’intelligenza, l’affettività, l’istintualità, la volontà, hanno sede fisica in strutture cerebrali ben definite, ed
operano in modo diffuso attraverso connessioni interneuronali. Le strutture mnesiche cerebrali si
formano durante lo sviluppo embrionale e la loro particolare espressione dipende dalla struttura del
DNA, mentre il tipo di sviluppo e l’esperienza determinano la quantità di ricordi. Così la memoria,
come ogni funzione psichica, ha due aspetti: uno innato, di base che dipende dal DNA dei genitori e un
altro, acquisito, dipendente dal tipo e quantità di stimoli e dall’ambiente; entrambi gli aspetti sono
importanti per avere nell’età adulta un solido impianto delle funzioni psichiche.
L’apprendimento e la memoria sono influenzati da altri processi psichici, così veglia e stato
emotivo condizionano la quantità e il tipo di informazioni che vengono memorizzate.
La memoria rappresenta il “copione” che descrive lo svolgersi dell’esistenza della persona, nella
sua continuità e nella costanza della sua identità.
La memoria, pertanto, nel suo aspetto sensoriale ed emotivo, dà un sottofondo unitario ed
integrante agli eventi del mondo interno.
Confrontarsi con il passato significa interrogarsi sul significato dell’esistenza nel suo continuum
temporale. Un flusso di figure e di episodi del passato si impone alla coscienza agganciandosi al
presente, per gettare un ponte tra ciò che si è stati, ciò che si è e ciò che si vorrebbe essere.
2. ASPETTI PSICOLOGICI DELLA MEMORIA
La natura della memoria e le modalità del suo funzionamento sono state affrontate in termini
diversi e talvolta contrastanti dalle varie scuole psicologiche in base ai rispettivi orientamenti teorici. Le
prime ricerche, a partire da Ebbinghaus, si basano sul modello associativo, che in seguito è stato
1
contestato dalla psicologia della forma secondo la quale la memoria non è spiegabile in termini di nessi
associativi ma, come per i processi percettivi, in termini di organizzazione dell’insieme memorizzato,
che tende sempre ad assumere la struttura più semplice, più economica e più regolare. A questa tesi si è
opposto il comportamentismo, per il quale la memoria è “un capitolo” dello studio dell’apprendimento
che è possibile spiegare in termini di condizionamento sul modello stimolo-risposta.
Il cognitivismo ha contestato la tesi comportamentista relativa all’unicità del processo di
memorizzazione, distinguendo una memoria a lungo, a breve e a brevissimo termine (o memoria
iconica). Quest’ultima non sarebbe sottoposta a fenomeni di interferenza semantica come accade alle
prime due forme di memoria. Inoltre la memoria a lungo termine non avrebbe alcun rapporto con
l’ambito percettivo, che invece caratterizza la memoria a breve termine e quella iconica. Il cognitivismo,
che ha fatto della memoria l’oggetto specifico del suo studio, ha inoltre elaborato modelli interpretativi
mutuati dalle categorie concettuali della teoria dell’informazione e della cibernetica. Infatti la
costruzione di memorie per calcolatori permette di elaborare modelli che si rivelano validi anche per il
funzionamento della mente umana e viceversa. Infine, nell’ambito della sociobiologia, R. Dawkins ha
introdotto il concetto di “meme”, che a suo avviso rappresenterebbe la controparte culturale del gene,
sarebbe cioè l’unità dell’ereditarietà culturale che avrebbe alcune proprietà dell’evoluzione biologica con
possibilità di trasmissione culturale da individuo a individuo. Dawkins distingue i memi, che sono le
unità di informazione conservate nella memoria, dai prodotti dei memi, che sono le loro manifestazioni
esterne percepibili.
È importante che il cervello non sia impegnato in altri compiti quando memorizza un evento.
La questione dell’interferenza, ossia di come le nuove informazioni apprese interagiscono con le
vecchie, è un punto fondamentale per la formulazione di teorie dell’apprendimento e del ricordo.
Se ad una scolaro si presentano definizioni senza che tra queste sia lasciato del tempo e sempre
in una forma diversa, egli non può immagazzinare tutto il materiale nello stesso momento, poiché il
processo biologico dell’immagazzinamento di informazioni (la memoria immediata) non lo consente. Il
docente, allora, non fa altro che distruggere gli impulsi già eccitati con informazioni nuove che li
inibiscono proprio per la loro somiglianza. Si parla in questo caso di interferenza: ciò che sarebbe stato
necessario per l’apprendimento e la ritenzione, cioè una risonanza interna dei contenuti mnesici già
esistenti, non può verificarsi.
Se si deve recuperare un ricordo, è più facile farlo se esso risveglia un significato, se si riferisce
ad un oggetto animato o inanimato, meno se ne è privo.
Certe cose si possono ritenere solo per qualche istante, altre invece il tempo appena necessario
per superare un esame, altre ancora rimangono in testa per tutta la vita. Ciò avviene perché entrano in
gioco emozioni intense, desideri personali, preferenze, e naturalmente i momenti di straordinaria
commozione, le situazioni importanti, oppure eventi spiacevoli, che colpiscono o impressionano
profondamente, per un motivo qualsiasi.
La fissazione e la rievocazione del vissuto sono strettamente associate al corretto
funzionamento dell’attività di coscienza. Non tutto ciò che si presenta in ogni istante nel campo di
coscienza viene stabilmente memorizzato: è immagazzinato solo ciò che ha una rilevante connotazione
emotiva.
Tanto più profondo è il livello di destrutturazione della coscienza, tanto minore è la fissazione
del vissuto.
Per quanto riguarda la particolare esperienza del ricordo del vissuto, si è notato che i sogni
possono essere ricordati, almeno per qualche tempo, se prodotti nel sonno REM (rapid eye moviments)
immediatamente precedente al risveglio. Anche le esperienze deliranti e allucinatorie dei disturbi della
coscienza tendono a essere più o meno rapidamente cancellate dalla memoria.
Ricordare non corrisponde a recuperare immagini statiche immagazzinate nella memoria, non
viene acquisito l’evento di per sé ma la potenzialità di attivare categorie di quella esperienza. Ricordare
vuol dire ricostruire dinamicamente un’esperienza in un nuovo contesto, attraverso il continuo
rimaneggiamento di mappe neuronali, attraverso un continuo lavoro di elaborazione e ricatalogazione in
termini di rilevanza biologica/adattiva, risignificazione.
2
Il ricordo non è, perciò, la semplice ripetizione di una esperienza passata ma la ricostruzione di
una fantasia, secondo procedure non dissimili da quelle della immaginazione. L’immaginazione - nel
senso di previsioni e programmi - si dispiega come una ininterrotta narrazione di potenzialità future che
originano da eventi passati. Il confronto tra scenari ricordati e scenari immaginati, tra esperienze vissute
e obiettivi prefigurati, tra le emozioni ed i sentimenti che ad essi si accompagnano, guiderà la scelta delle
decisioni future.
La memoria - come Giano bifronte - guarda al passato, ma anche al futuro: le prospettive
tendono ad avere una coerenza con le esperienze passate. Sorgente di tale continuità è la incessante
ricostruzione della propria identità, ottenuta modellando - anche in termini neuronali - il passato ed il
futuro come aspetti di una stessa scena, attraverso la capacità - solo umana - di raccontare a sé e ad altri
il passato ed il futuro. Nella narrazione vecchie storie vengono risignificate, pensieri e sentimenti del
passato vengono distanziati dal presente, fantasie sul futuro vengono attualizzate e rivalutate.
L’inconscio è essenzialmente memoria che non può essere ricordata. Gran parte delle
operazioni psichiche che vengono comprese sotto la definizione di “memoria”, avvengono al di fuori
del controllo della coscienza.
La selezione del materiale percettivo, l’immagazzinamento delle cosiddette “tracce mnesiche”, il
riaffiorare improvviso di ricordi estranei al contesto del momento, sono tutti processi estranei alla
volontà cosciente.
Secondo S. Freud, la traccia mnestica non è un’immagine della cosa, ma la sua trascrizione
nell’apparato neuronico dove gli engrammi sono depositati in diversi sistemi e in relazione con altre
tracce fra loro legate per simultaneità, causalità e successione cronologica. Questi sistemi sono alla base
delle catene associative, mentre il loro grado di accessibilità alla coscienza dipende dalla riattivazione
promossa dall’investimento, dal controinvestimento e dal disinvestimento.
Proprio il processo analitico, che possiamo vedere come un paradigma generale dello svolgersi
della persona e delle sue relazioni, corre lungo la memoria, così come l’abbiamo intesa, ed ha come fine
la percezione più ordinata e precisa degli oggetti interni e della loro reciproca posizione, funzione
strettamente legata alla memoria.
Freud distingue la rappresentazione e l’importo d’affetto, per cui tutti i ricordi in linea di
principio vengono trascritti, ma la loro rievocazione dipende dal modo in cui vengono investiti: «Nelle
funzioni psichiche bisogna distinguere qualcosa (importo d’affetto, somma di eccitazione) [...] che può
essere aumentato, diminuito, spostato, scaricato e che si stende sulle tracce mnesiche delle
rappresentazioni quasi come una carica elettrica sulla superficie dei corpi». La teoria della traccia
mnesica costringe Freud a stabilire un’incompatibilità tra coscienza e memoria: «Troviamo difficile
credere che l’eccitamento lasci anche nel sistema P-C [percezione-coscienza] tracce permanenti siffatte.
Se rimanessero sempre consce, ben presto limiterebbero la capacità del sistema di ricevere nuovi
eccitamenti, se invece diventassero inconsce, ci metterebbero di fronte al problema di spiegare
l’esistenza di processi inconsci in un sistema il cui funzionamento è per il resto caratterizzato dai
fenomeni propri della coscienza. Con la nostra ipotesi che situa il processo del diventare cosciente in un
particolare sistema non avremmo per cosi dire cambiato nulla né guadagnato nulla. Pur ammettendo
che questa considerazione non sia probante in modo assoluto, essa può tuttavia indurci a supporre che
il diventare cosciente e il lasciare dietro di sé una traccia mnestica siano processi tra loro incompatibili
all’interno di uno stesso sistema».
S. Freud parla di «amnesia infantile» a proposito dell’estrema difficoltà nel ricordare gli eventi
della prima infanzia. Le ipotesi non psicoanalitiche formulate in proposito sono: o che il bambino
esperisca il mondo in modo diverso dall’adulto per cui da adulto non riesce a rievocare le scene infantili,
oppure che lo scarso immagazzinamento dipenda dallo scarso sviluppo del linguaggio.
Per Freud, alla base dell’amnesia infantile c’è la rimozione.
C. G. Jung utilizza il concetto di engramma per spiegare la memoria inconscia: «L’inconscio,
considerato il campo storico della psiche, contiene in forma concentrata l’intera serie di engrammi che
da tempo incommensurabile hanno condizionato l’attuale struttura della psiche. Gli engrammi non
sono che tracce funzionali che indicano la frequenza e l’intensità massime secondo le quali
generalmente la psiche umana ha funzionato. Questi engrammi funzionali si presentano come motivi e
3
figure mitologiche che si riscontrano - talora identici, talora assai simili - in tutti i popoli e che si
possono agevolmente ravvisare anche nei materiali inconsci dell’uomo moderno».
L’oblio è la dimenticanza parziale o totale delle esperienze passate, descrivibile graficamente con
la curva dell’oblio, messa a punto per la prima volta nel 1885 da Ebbinghaus, che dimostra come la
diminuzione del ricordo sia rapida all’inizio e più lenta in seguito. L’intensità della caduta di detta curva
dipende ovviamente dalla qualità del materiale appreso (verbale o motorio, con senso o senza senso),
dal tipo di memoria misurata (reintegrazione, rievocazione, riconoscimento), dalle condizioni di
apprendimento con le conseguenti ripercussioni sull’immagazzinamento e sul consolidamento. Per
spiegare l’oblio esistono tre teorie interpretative:
La teoria della traccia mnestica che ipotizza, oltre a un decadimento spontaneo della traccia col
passare del tempo per effetto dei normali processi metabolici del cervel1o, una sua distorsione
sistematica responsabile più dei cambiamenti qualitativi che di quelli quantitativi del ricordo. La
psicologia della forma ha mostrato come una relativa omogeneità fra gli elementi percepiti o la
mancanza di una chiara e definita struttura crei condizioni sfavorevoli non solo per la fìssazione del
ricordo, ma anche un’inibizione retroattiva per gli altri, per cui nel ricordo le figure tendono a divenire
più simmetriche di quel che sono, oppure accentuano le loro irregolarità; se poi assomigliano a qualche
oggetto specifico, tale somiglianza viene accentuata con conseguenti distorsioni, come nel caso dei
racconti successivi di una stessa storia.
La teoria dell’interferenza o inibizione che ipotizza un’inibizione dovuta all’attività svolta
nell’intervallo di tempo che corre tra l’apprendimento e il ricordo. A questo proposito si distingue
un’interferenza o inibizione retroattiva quando ogni nuova acquisizione rende più difficile il ricordo del
materiale precedentemente acquisito, ed una proattiva quando rende più difficile l’apprendimento e il
ricordo di materiale nuovo. L’interferenza si registra non solo tra compiti differenti. ma anche tra
sequenze dello stesso compito con maggior interferenza retroattiva nei riguardi delle prime parti o
maggior interferenza proattiva nei riguardi delle ultime, mentre le parti centrali sono oggetto di
entrambe le interferenze, per cui sono le più difficili da ricordare. Una diminuzione dell’interferenza si
ha riducendo, e al limite eliminando, l’attività tra l’apprendimento e il ricordo; ciò spiega perché l’oblio,
durante il sonno, è assai inferiore all’oblio durante lo stato di veglia.
La teoria della rimozione, elaborata da Freud, che ha concepito l’oblio come un meccanismo di
difesa volto a proteggere la psiche da esperienze dolorose che possono riferirsi a desideri insoddisfatti, a
conflitti non risolti o a impulsi socialmente riprovevoli. Questo tipo di oblio non comporta la
scomparsa o il deterioramento delle tracce mnestiche, ma semplicemente il loro mancato recupero che
l’analisi si propone di riattivare. I meccanismi indicati da Freud quali responsabili di questo oblio sono
la condensazione e lo spostamento, non il tempo: «Se le tracce mnestiche sono rimosse, si può rilevare
che esse non subiscono modificazioni durante lunghissimi intervalli di tempo. L’inconscio è soprattutto
fuori del tempo. Il carattere più importante e più strano della fissazione psichica è che tutte le
impressioni si conservano. non solo nella medesima maniera in cui furono accolte, ma anche in tutte
quelle forme che hanno assunto durante gli sviluppi ulteriori. [...] Secondo la teoria, dunque, si potrebbe
ripristinare il ricordo di ogni stato precedente del contenuto mnemonico, anche se i suoi elementi
hanno da tempo sostituito tutte le relazioni primitive con altre più recenti. Partendo dall’indicazione
freudiana secondo cui l’oblio da rimozione è un meccanismo di difesa psichica, C. Musatti formula la
legge dell’ottimismo mnestico secondo cui «noi esercitiamo proprio un’azione selettiva sull’insieme dei
nostri ricordi, nel senso che ricordiamo più facilmente e durevolmente gli avvenimenti piacevoli che
non gli accadimenti spiacevoli del nostro passato. Ciò dimostra l’effettiva tendenza a dimenticare che si
sviluppa in presenza di elementi spiacevoli. Questa tendenza risponde appunto a un’intrinseca finalità
biologica: la difesa dal dolore», che agisce anche quando si è ossessionati da ricordi spiacevoli, perché
questi hanno sostituito altri ricordi sepolti ancora più spiacevoli.
Un’ulteriore spiegazione dell’oblio si potrebbe trovare nel fatto che, durante il processo di
formazione della rete di cellule cerebrali, certi percorsi si sono fissati meglio di altri: gli impulsi vi
passano più facilmente favorendo il collegamento delle associazioni corrispondenti. In qualunque
momento. sia avvenuta questa modificazione strutturale provocata da esperienze o pensieri
particolarmente incisivi, l’effetto è lo stesso: impulsi che magari dovrebbero passare per le stesse cellule,
4
ma attraverso altre ramificazioni, sono svantaggiati e non giungono a destinazione benché in teoria
esistano le relative connessioni e il ricordo sia immagazzinato.
Talvolta l’oblio è importante per la vita: protegge ed aiuta a mantenere l’equilibrio psichico.
Certamente queste informazioni che non si riesce a richiamare volontariamente alla coscienza
influiscono dall’inconscio sul comportamento quotidiano e perciò la psicoterapia si è proposta di
studiare con i suoi metodi (uno di questi è l’ipnosi) le cause del comportamento non controllato
coscientemente.
3. ANATOMO – FISIOLOGIA DELLA MEMORIA
Il processo mnesico è l’espressione non di un singolo sistema unitario, ma piuttosto di una
interazione di molteplici sistemi che modulano le varie fasi del processo stesso (registrazione,
immagazzinamento e rievocazione delle informazioni). I sistemi conivolti sono molteplici, in diverse
regioni corticali (il lobo temporale, la regione frontale sia centromediale sia dorsolaterale, nuclei
corticali), in nuclei della sostanza grigia sottocorticale (l’ippocampo, il talamo) ed nel cervello anteriore
basale.
Un’informazione in arrivo non viene semplicemente trattenuta o dimenticata, ma si sofferma in
tre stadi di diversa durata.
I sistemi, biologici o artificiali, di immagazzinamento dell’informazione necessitano di:
1. poter essere codificati (encoding);
2. poter essere immagazzinati (storage);
3. poter essere accessibili e poter essere recuperati (retrieval).
Tutti gli impulsi che pervengono all’individuo tramite le percezioni derivanti dagli organi di
senso, circolano immediatamente nel cervello sotto forma di oscillazioni elettriche e, dopo dieci o venti
secondi, si perdono. Se non c’è alcun interesse o se le informazioni si possono ricollegare ad
associazioni già presenti, queste percezioni passano dentro di noi come i rumori della strada o i suoni di
una lingua straniera.
La mancanza di interesse e l’impossibilità di stabilire delle associazioni, oppure la
contemporaneità di percezioni distraenti (ad es. il dolore), portano alla dispersione dell’informazione
elettrica originaria, senza che essa venga consolidata nella memoria.
Anche quando siamo di fronte ad avvenimenti con un’intensa coloritura emotiva, prima che il
ricordo venga immagazzinato in maniera definitiva, l’informazione deve passare attraverso la memoria
immediata e quella a breve termine.
Quando le informazioni vengono etichettate come «non interessanti per un ulteriore
immagazzinamento» e quindi respinte già a livello di memoria immediata, queste percezioni devono
essere dimenticate rapidamente: per permettere reazioni adeguate, infatti, non possono essere elaborate
fino in fondo nella corteccia cerebrale.
Mentre nei normali processi di pensiero gli impulsi in arrivo tramite la vista o l’udito vengono
elaborati nella corteccia cerebrale provocando reazioni coscienti, in questo caso invece le percezioni
vengono avviate direttamente ai nervi motori, e quindi trasformate immediatamente in movimenti
muscolari senza render necessario l’intervento del pensiero a livello delle cellule grigie del cervello.
La codificazione di materiale che avviene facendo riferimento a conoscenze pregresse, consente
una rievocazione migliore rispetto alla semplice ripetizione di sequenze verbali. Questo determina la
creazione di codici semantici più ricchi che permettono al materiale di essere codificato secondo un
numero di dimensioni maggiore rispetto ai codici fonologici o visivi.
Interi gruppi di sinapsi possono essere bloccati da esperienze di stress che disturbano le
sostanze trasmittenti.
Gli impulsi giunti al cervello entrano in una specie di circuito elettrico chiuso, che si eccita per
un tempo brevissimo. Per quanto breve sia la permanenza nella memoria, si possono salvare le
5
informazioni dall’estinzione richiamandole entro pochi secondi, così vengono associate a qualche
contenuto mnesico già immagazzinato oppure entrano in risonanza con ricordi già esistenti.
La memoria immediata può essere distrutta con una lieve stimolazione elettrica, cancellando
tutta l’informazione registrata negli ultimi diciotto secondi, cosicché il programma d’apprendimento
deve ricominciare da capo. Il secondo stadio, la memoria a breve termine (che dura all’incirca per venti
minuti), può ancora essere cancellata, al contrario di quella a lungo termine, soltanto con un forte choc
(basta pensare all’esempio dell’incidente).
Il passaggio dalla memoria immediata a quella a breve termine corrisponde ad un filtro ed ha
quindi una funzione assai importante: questo filtro ci protegge (almeno parzialmente) dal sovraccarico
di informazioni, facilitando così l’orientamento.
È possibile dimenticare anche avvenimenti che, per la loro importanza, per la loro attrazione
particolare o l’alto significato personale, sono già stati registrati coscientemente. Impressioni forti, che
senz’altro hanno superato la memoria immediata, cioè la fase puramente elettrica della corrente di ioni,
possono ancora cancellarsi interamente. Questo è possibile finché non sono passate nella memoria a
lunga termine.
I ricordi dell’esperienza si possono cancellare per mezzo di una lieve stimolazione cerebrale, ma
soltanto nei primi venti o trenta secondi: questa è una prova della natura elettrica, non materiale, della
memoria immediata. Altrimenti non si potrebbe bloccare con tanta facilità il passaggio
dell’informazione dalla memoria immediata a quella a breve termine.
Dopo alcuni minuti dall’apprendimento, quando il ricordo ha già superato la memoria
immediata, ci vuole almeno un elettrochoc per estinguerlo, perché si tratta di sopprimere il passaggio
dalla memoria a breve termine, già più solida, a quella a lungo termine. Alcune ore o giorni dopo il
processo dell’apprendimento, infine, non basta neanche l’elettrochoc per far dimenticare quanto è stato
registrato; ciò dimostra che il ricordo è già passato nella memoria a lungo termine, dove rimane
conservato per sempre.
Tali fenomeni si possono notare anche in incidenti stradali quando l’automobilista subisce uno
choc. In questi casi la constatazione dei fatti è un’impresa assai problematica. L’automobilista dovrebbe
spiegare agli agenti la dinamica dell’incidente: cerca di pensarci, ma gli è impossibile ricordare qualcosa.
Anche la domanda, da quale parte gli sia venuto addosso il pedone, non aiuta. Sa soltanto che direzione
ha preso a partire da casa e che stava arrivando sui viali di circonvallazione. Ogni volta che si sforza di
richiamare alla mente gli avvenimenti successivi, perde il filo; ancora molto prima dell’incidente, il
ricordo gli sfugge.
Uno stimolo sensibilizzante (nocivo) che attiva un gruppo di neuroni (facilitanti), alcuni dei
quali hanno come neurotrasmettitore la serotonina (5HT), può indurre i neuroni facilitanti a formare
sinapsi asso-assoniche con le terminazioni dei neuroni sensoriali.
Il substrato cellulare della sensibilizzazione a breve termine è una forma di plasticità sinaptica
detta facilitazione presinaptica, in cui la 5HT (liberata dallo stimolo nocivo) interagisce con recettori
sulla membrana del neurone sensoriale attivando una proteina G che a sua volta attiva l’enzima
adenilato-ciclasi determinando l’aumento della concentrazione di AMP ciclico (AMPc) nei neuroni
sensoriali. L’AMPc attiva la protein-chinasi A (PKA) che determina la fosforilazione di diversi substrati
proteici. Nel neurone sensoriale l’attivazione della PKA produce tre importanti conseguenze: in primo
luogo viene fosforilata una proteina integrale di un canale per il K+, ciò produce un aumento o una
diminuzione della sua attività; secondariamente viene mobilizzata una maggior quantità di
neurotrasmettitore; in terzo luogo si aumenta la quantità di vescicole sinaptiche disponibili. L’insieme di
queste tre azioni determina una maggior liberazione di neurotrasmettitore e di conseguenza un aumento
dell’efficacia sinaptica. La conseguenza comportamentale è la seguente: una stimolazione tattile data in
una determinata sequenza produce una risposta riflessa sempre più ampia e duratura in quanto le
sinapsi che i neuroni sensoriali formano con il versante motorio del riflesso diventano sempre più
efficaci. Il ricordo neuronale dell’esperienza dolorosa dura quanto l’aumento del riflesso di retrazione
evocato dalla stimolazione tattile, per il periodo in cui le sinapsi sensitivo-motorie rimangono facilitate.
La maggiore efficacia sinaptica rimane tale fino a quando i substrati proteici della PKA rimangono
fosforilati.
6
Alcune conclusioni possono essere tratte da questi risultati.
Eventi correlati con l’apprendimento possono essere identificati in distinti distretti cellulari e
strutture molecolari.
L’apprendimento a breve termine implica modificazioni nel circuito neuronale che sottende al
comportamento.
I cambiamenti avvengono a livello delle sinapsi.
La modificazione dell’efficacia sinaptica avviene attraverso l’attivazione di un secondo
messaggero. La cellula nervosa usa la cascata molecolare di secondi messaggeri intracellulari (come
quella dell’AMPc), anche come mezzo di immagazzinamento dell’informazione e modificazione di
rappresentazioni interne.
La memoria a breve termine ha come substrato molecolare una modificazione covalente di
proteine preesistenti per cui svanisce nell’arco di poche ore. La ragione di questo deterioramento è
duplice: innanzitutto queste modificazioni covalenti sono rapidamente reversibili; in secondo luogo se
anche la modificazione covalente fosse irreversibile la molecola proteica ha una vita breve, da qualche
minuto a qualche giorno.
4. MEDIATORI CHIMICI DELLA MEMORIA
I sistemi neurotrasmettitoriali coinvolti nei processi mnesici sono numerosi e comprendono la
noradrenalina, la dopamina, la serotonina, l’acetilcolina, il GABA e i neuropeptidi.
Per quanto riguarda il sistema noradrenergico, esso è costituito principalmente da corpi cellulari
che sono localizzati a livello del locus coeruleus, nel mesencefalo.
L’attività del sistema noradrenergico è correlata allo stato di vigilanza e quindi influenza in
modo indiretto i processi di apprendimento e di memorizzazione.
Il sistema dopaminergico, attraverso la via mesolimbico-corticale, proietta su regioni limbiche
come l’ippocampo dorsale e su aree della corteccia frontale e prefrontale. Esso svolge quindi un ruolo
importante nel controllo dei processi di apprendimento.
Sembra che le proiezioni colinergiche facilitino l’elaborazione corticale degli stimoli modulando
le interazioni tra sistema serotoninergico e noradrenergico.
La specificità delle vie colinergiche a partenza dal nucleo basale di Meynert, le sue proiezioni
all’ippocampo e alla corteccia cerebrale, lo specifico effetto amnesico dei farmaci anticolinergici e la
distruzione dei neuroni colinergici nella demenza tipo Alzheimer, sembrano rendere univoca la
relazione tra memoria e sistema colinergico.
I nuclei colinergici basali e le loro connessioni con le formazioni ippocampali assumono
importanza per il loro ruolo nelle funzioni cerebrali superiori.
Il ruolo del sistema colinergico nella regolazione dei processi mnesici è stato desunto dal
riscontro di un deficit a carico di questo sistema in patologie in cui le capacità mnesiche sono
selettivamente compromesse, come nella demenza di tipo Alzheimer.
I neuroni colinergici localizzati in particolari regioni del SNC, aventi un ruolo significativo nei
processi di apprendimento e di memorizzazione, suscettibili di danni irreversibili nella malattia di
Alzheimer, sono particolarmente sensibili alle neurotrofine. Infatti la somministrazione del NGF è in
grado di contrastare danni sperimentali in questi neuroni colinergici.
Di tutte le informazioni sensoriali che lo bombardano, il cervello deve scegliere quali ricordare.
Lo fa con l’aiuto di due classi di proteine cerebrali con funzioni opposte, raggiungendo un equilibrio fra
attivazione e inibizione dei processi biochimici della memoria. Un rappresentante di queste proteine, la
fosfatasi 1 (PP1) ha un ruolo nella cancellazione dei ricordi, infatti animali privi di questa proteina
imparano bene quanto gli altri ma dopo un intervallo superiore a cinque minuti il cervello, avendo la
possibilità di bloccare la PP1, può cementare i ricordi.
Il calo di memoria dovuto all’età è in parte dovuto all’aumento di PP1 rispetto alle proteine che
aiutano la memoria, infatti inibendo la PP1 si può invertire il declino mnesico associato
all’invecchiamento.
7
Inoltre altri componenti proteici cerebrali sembrano agire favorendo l’eliminazione dei ricordi,
spiegando così il peggioramento della memoria con l’età e le modalità attuate dal cervello per eliminare
le informazioni non essenziali.
Il ricordo non arriva alla memoria a lungo termine se è bloccata la sintesi di proteine. Bloccando
questa sintesi più tardi, cioè quando il ricordo è già conservato nella memoria a lungo termine, non si
riesce più ad estinguerlo.
Una micromemoria è saldamente registrata negli acidi nucleici delle cellule come le parole di una
enorme e nello stesso tempo minutissima biblioteca, della quale la nostra riproduzione ci fa vedere
soltanto una sola frase. Ogni singola cellula può richiamare, secondo il bisogno, questi programmi e
ordini depositati, cioè può ricordarsene: esattamente come facciamo noi in grande coi ricordi nel nostro
cervello.
Non si parte dall’intero contenuto di informazioni, ma da un codice molecolare, da frammenti
che compongono un tutto organico, come i singoli suoni e vibrazioni formano un brano musicale.
È recente la scoperta di un processo cerebrale che controlla la formazione dei ricordi
permanenti. Segnali scambiati fra i neuroni stimolano la produzione delle proteine necessarie
all’immagazzinamento a lungo termine dei ricordi.
I ricordi che perdurano a lungo vengono conservati nel cervello mediante il rafforzamento delle
connessioni, o sinapsi, fra i neuroni. Ciò era già noto da tempo ai ricercatori, perché i neuroni devono
attivare la sintesi di nuove proteine per poter immagazzinare la memoria a lungo termine e rafforzare le
sinapsi, ma il meccanismo con il quale essi svolgono questo compito era ancora sconosciuto. È
acquisizione recente che la via molecolare che consente ai neuroni di aumentare rapidamente la propria
produzione di nuove proteine durante la formazione dei ricordi a lungo termine e il rafforzamento
sinaptico è controllata da un enzima (il MAPK), che innesca l’immagazzinamento dei ricordi a lungo
termine stimolando la sintesi di proteine.
Secondo alcuni ricercatori anche la cipina, una proteina presente in tutto il corpo, nel cervello
regola la ramificazione delle cellule nervose.
La cipina interferisce con le modificazioni di forma dei neuroni, per lo più nell’ippocampo, la
struttura del cervello associata con la regolazione delle emozioni e dei ricordi.
La crescita dendritica aumenta quando si apprende qualcosa. Una riduzione di questa
ramificazione è associata a diversi disturbi neurologici.
I processi cerebrali responsabili dell’immagazzinamento e dell’elaborazione dei ricordi sono
simili a quelli dell’informazione genetica: la traccia è costituita da acidi ribonucleici, (RNA), su di essa si
raggruppano diverse molecole di aminoacidi nell’ordine prescritto. Queste si uniscono direttamente ad
opera dei cosiddetti «ribosomi», piccole strutture a forma di panini, atte ad annodare diversi amino
acidi. Passando per questi ribosomi si formano dagli amino acidi catene di proteine, mentre la matrice di
RNA si dissolve.
Questo processo, comune a tutte le cellule, nei neuroni, al contrario delle cellule normali,
comporta che determinati segnali percettivi stimolino a formazione di RNA, fissando materialmente il
ricordo di tali segnali.
Nelle cellule cerebrali, che non si dividono, non aumentano e non crescono più, partecipando
solo a pochi processi metabolici, queste proteine vengono conservate ed immagazzinate ed acquistano il
ruolo di un deposito inattivo di informazioni. Quando sono riattivate da certi segnali, possono stimolare
come molecole di riconoscimento le rispettive cellule cerebrali ad emettere anche loro dei segnali. In tal
modo le cellule cerebrali coinvolte possono essere richiamate da determinate percezioni: è questa la
base per tutti i processi di pensiero e di memoria.
Prima del decadimento della memoria immediata, la sua informazione viene assunta nel
magazzino della memoria a breve termine, il quale è legato alla formazione di una matrice di RNA, che
dura per circa 20 minuti, sciogliendosi poi di nuovo come l’originale nella stampa, che dopo l’uso viene
fuso. A questo punto quindi l’informazione deve essere già passata alla memoria a lungo termine
mediante la formazione di certe proteine. La stessa memoria a lungo termine è collegata quindi al
deposito stabile delle proteine formatesi sulla rispettiva matrice di RNA.
8
5. NEUROPEPTIDI E PLASTICITÀ
Il SNC si trova in uno stato di equilibrio dinamico, al mantenimento del quale partecipano i
fattori di crescita e di differenziazione mediante la fine regolazione dei circuiti sinaptici.
Il termine “neuropeptidi” (NRP) identifica i peptidi, composti formati da due o più aminoacidi,
contenenti uno o più “legami peptidici” –CONH, collocabili a livello intermedio tra aminoacidi e
peptoni nella sintesi delle proteine, per i quali è stata dimostrata presenza o funzione all’interno del
sistema nervoso.
I NRP vengono sintetizzati come una normale proteina a livello dei ribosomi, cioè nel
pericarion del neurone. Poi i NRP vengono immagazzinati in vacuoli fino al rilascio, che avviene grazie
ad un meccanismo di trasporto attivo Ca ++ dipendente
I peptidi, così come gli ormoni, dei quali condividono alcune proprietà, sono efficaci in
concentrazioni notevolmente inferiori a quelle dei neurotrasmettitori, nell’ordine di nanomoli, e talvolta
agiscono a distanza dal sito di secrezione.
Sembra che i NRP agiscano provocando modificazioni rapide sulle membrane neuronali e, per
questo, alcuni Autori hanno ipotizzato che possano determinare anche il rilascio di neurotrasmettitori
classici.
Peptidi come l’ormone adrenocorticotropo (ACTH), la vasopressina, l’ormone stimolante i
melanociti (MSH), intervengono in processi cognitivi quali l’apprendimento, la memoria, l’attenzione e
la prestazione in test sperimentali.
L’ACTH avrebbe un effetto stimolante sulla memoria a breve termine: a livello centrale l’ACTH
e suoi analoghi hanno mostrato di influenzare e migliorare le capacità di prestazione, attenzione,
vigilanza, apprendimento e memoria.
La vasopressina (VSP, nota da anni come ormone antidiuretico, dati i suoi effetti periferici sulla
regolazione dei fluidi e del bilancio elettrolitico) facilita il consolidamento dei ricordi, migliora il
richiamo dei dati memorizzati, la concentrazione e l’attenzione in pazienti affetti da sindromi psicoorganiche.
La vasopressina partecipa alla memoria dei comportamenti acquisiti, infatti ritarda l’estinzione
dei comportamenti appresi. L’ossitocina, invece la accelera, facilitando l’estinzione di comportamenti
appresi (azione amnestica).
La VSP avrebbe effetti facilitatori sulla memoria sociale, mentre l’ossitocina interferirebbe con il
riconoscimento dei propri simili,
L’MSH ha, invece, migliorato i test di memoria visiva nell’uomo, non quelli di ritenzione
verbale.
Situazioni di apprendimento che comportano uno stress determinino sempre la liberazione di
endorfine.
La prolattina (PRL) è coinvolta nei processi di apprendimento e memorizzazione, in quanto si è
osservato che bassi livelli liquorali di PRL nell’uomo sono correlati a deficit mnesici.
La Sostanza P (SP) nell’ipotalamo laterale stimola i processi di apprendimento e di memoria,
nell’amigdala li riduce.
Alcuni fattori trofici, chiamati di differenziazione neuronale, possono indirizzare un particolare
neurone ad assumere un certo profilo trasmettitoriale e neuropeptidergico e controllare il tipo di sinapsi
che questo instaura con le cellule cui si rapporta.
Le citochine neuropoietiche sono i fattori di differenziazione più studiati. Producono effetti sia
sul sistema nervoso sia su quello emopoietico e possono intervenire nei processi di riparazione
conseguenti a danno neuronale.
I fattori di crescita e di differenziazione nel SNC possono essere coinvolti, così, nelle risposte a
danneggiamenti del sistema nervoso, nell’interconnessione fra il sistema nervoso e il sistema
immunitario, nei cambiamenti dei ritmi biologici degli organismi (come avviene per gli ormoni
steroidei), nelle trasformazioni della plasticità sinaptica correlate alla memoria ed all’apprendimento,
nella trasmissione sinaptica.
9
I neuropeptidi oltre ad essere implicati in molte funzioni a livello tissutale, di regolazione di
funzioni neuroendocrine, possiedono anche proprietà di modulazione del comportamento. Ciò in
considerazione della co – localizzazione insieme a neurotrasmettitori classici nelle sinapsi e nelle vie
della neurotrasmissione.
Vie neurotrasmettitoriali classiche sono modulate dai neuropeptidi, poiché è dimostrata la colocalizzazione con amine biogene, acetilcolina e Acido gamma amino butirrico (GABA).
Lesioni del SNC o di nervi periferici danno luogo ad un aumento di produzione di molti dei
fattori di crescita e di differenziazione sinora considerati. L’incremento rilevato potrebbe causare in
modo direttamente proporzionale la comparsa e la crescita di nuovi assoni neuronali
concomitantemente all’incremento di neuropeptide.
Il compito dei NRP è quello di recuperare, mediante processi di plasticità neuronale e/o di
crescita di nuove fibre nervose, la funzionalità danneggiata o persa.
L’effetto delle neurotrofine si produce sia su neuroni siti nel SNC sia localizzati nel SN
periferico. Per poter ottenere una risposta dalle neurotrofine deve essere presente il recettore specifico
per una determinata neurotrofina sulla membrana del neurone.
I neuropeptidi coinvolti nei processi di memorizzazione sono principalmente l’ossitocina e la
vasopressina (VSP) ma lo sono anche l’ACTH, gli OE e la pancreostatina.
I processi di apprendimento e di memorizzazione si possono riassumere in quattro fasi:
acquisizione ed immagazzinamento della memoria a breve termine, che ha una durata massima
di un giorno;
consolidamento, che sembra coinvolgere strutture quali l’ippocampo e l’amigdala;
immagazzinamento in engrammi a lungo termine;
richiamo delle informazioni acquisite.
Neuropetidi ed amine possono influenzare tutti questi stadi, mentre il consolidamento sembra
verificarsi solo in presenza di sonno paradosso nelle 24 ore. Queste osservazioni hanno condotto
all’ipotesi che il consolidamento possa essere influenzato dagli alti livelli di VSP ed ACTH presenti
durante tale fase di sonno. Questo spiegherebbe come piccoli stress aumentino le capacità
mnemoniche.
L’ACTH ed i peptidi ad esso correlati si trovano nell’ipofisi, il sito principale di produzione, ma
è stato identificato un circuito neuronale contenente ACTH a partenza dal nucleo arcuato
dell’ipotalamo che proietta principalmente al setto, all’ippocampo, ad altri nuclei ipotalamici e
all’amigdala. ACTH e MSH sono anche presenti in varie aree nel midollo, nel ponte, nel mesencefalo,
nel corpo striato, nella corteccia e nel cervelletto.
L’ACTH sembra avere proprietà neurogeniche: variazioni delle concentrazioni cerebrali di
ACTH e dei peptidi ad esso correlati determinano evidenti alterazioni della funzione neuromuscolare e
del comportamento.
Questi fenomeni, che possono essere definiti di plasticità, di crescita e di espressione genica,
sono presenti anche nell’età adulta, anche se forse meno evidenti rispetto al periodo di sviluppo. Le
implicazioni legate alla presenza di questa plasticità sono:
il sistema è in uno stato di equilibrio legato al bilanciamento fra fasi di crescita e di non crescita,
induzione di geni e repressione di geni;
i fattori di crescita e di differenziazione sono indispensabili per mantenere l’attività del sistema
maturo;
di conseguenza questi fattori potrebbero essere utilizzati per interventi terapeutici.
È stato provato che fattori di differenziazione possono indirizzare l’espressione genica, tanto
che un neurone noradrenergico può essere convertito in un neurone di tipo colinergico.
Questo meccanismo mostra come sia presente “plasticità” sia nella crescita che nell’espressione
genica, più evidente nella fase di sviluppo ma presente anche nella fase adulta. Questa plasticità
comporta che il sistema neuronale si trovi in uno stato di equilibrio legato al bilanciamento fra fasi di
crescita e di stasi, di induzione e repressione di geni; che per mantenere l’attività del sistema maturo
siano indispensabili i fattori di crescita e di differenziazione, utilizzabili per interventi terapeutici.
10
I fattori di crescita e di differenziazione possono recuperare neuroni la cui attività si sta
spegnendo e modificare il bilanciamento delle attività svolte dai diversi sistemi trasmettitoriali e
neuropeptidergici.
Tra i fattori di crescita, denominati neurotrofine, il più conosciuto è il fattore di crescita
neuronale (NGF, nerve growth factor), capace di recuperare dalla morte alcuni gruppi di neuroni
embrionali, che muoiono durante le fasi di sviluppo.
I processi di derivazione cellulare, di migrazione cellulare, di crescita degli assoni e di plasticità
come pure i processi in base ai quali il neurone acquista delle specifiche caratteristiche che lo
contraddistinguono da altri neuroni, contribuiscono alla differenziazione cellulare. La differenziazione
cellulare permette ad una cellula di passare da un immaturo e non specifico fenotipo ad uno maturo e
specializzato.
Variazioni fisiologiche della stimolazione sensoria possono indurre notevoli cambiamenti
nell’espressione delle neurotrofine. L’esposizione alla luce dopo un periodo di buio può raddoppiare i
livelli di RNA messaggero per il BDNF nella corteccia visiva. L’aumento potrebbe spiegare il riscontro
della concomitante comparsa e crescita di nuovi assoni neuronali.
Fra i neuroni particolarmente sensibili alle neurotrofine ci sono quelli colinergici, localizzati in
particolari regioni del SNC, il cui ruolo nei processi di apprendimento e di memorizzazione e la loro
perdita nella malattia di Alzheimer sono stati ampiamente studiati. In animale da laboratorio si è stato
dimostrato che la somministrazione del NGF è in grado di contrastare i danni indotti da lesioni dei
neuroni colinergici.
“Potenziamento a lungo termine” (LTP) e “depressione a lungo termine” (LTD) sono implicati
in processi di plasticità neuronale in diverse zone del sistema nervoso.
Oltre alla durata, l’LTP ha tre caratteristiche che lo rendono indubbiamente candidato ad essere
un meccanismo mnemonico:
la cooperatività (non basta l’attività di una sola afferenza presinaptica per indurre l’LTP, è
necessaria l’attivazione di più fibre);
la specifi cità (l’LTP si genera solo a livello delle sinapsi formate dalle fibre afferenti attive e non
in altre eventuali afferenze);
l’associatività (l’induzione di LTP richiede una coincidenza temporale tra l’attività delle afferenze
presinaptiche e l’attività della cellula post-sinaptica).
l’LTP è presente nell’ippocampo, per l’induzione dell’LTP è necessaria la contemporanea attività
delle cellule presinaptica e post-sinaptica.
I fattori di crescita e di differenziazione sono in grado di agire nel tessuto nervoso adulto, come
pure di avere un ruolo funzionale nella risposta a danneggiamenti di questo tessuto.
Riassumendo si può affermare che i fattori di crescita e di differenziazione nel SNC possono
essere coinvolti in molti importanti processi, quali ad esempio:
le risposte a danneggiamenti del sistema nervoso come pure l’intercomunicazione fra il sistema
nervoso e il sistema immunitario;
i cambiamenti, come nel caso degli ormoni steroidei, che si hanno nei ritmi biologici degli
organismi;
i cambiamenti nella plasticità sinaptica coinvolti nella memoria e nell’apprendimento;
la trasmissione sinaptica.
6. L’ENGRAMMA
Non si può paragonare la memoria durevole a correnti permanenti di ioni ma, piuttosto, a una
sorta d’incisione (il cosiddetto «engramma»). Si dice infatti che una cosa «s’imprime nella mente»,
un’espressione che sembra rispecchiare bene il processo reale. L’immagazzinamento inizia con impulsi
elettrici cerebrali:è la memoria immediata che però in base alla sola esistenza di tali circuiti elettrici non
si mantiene in nessun caso più di qualche secondo.
Non esiste un centro neuronale della memoria. Le funzioni mnestiche sono correlate con le
formazioni nervose superiori e sono regolate dall’attività corticale le cui cellule conservano le tracce
11
mnestiche con ampia possibilità di vicariazione. Secondo alcuni la traccia mnestica o engramma è del
tutto ipotetica perché, come scrivono E. R. Hilgard e R. C Atkinson, «la traccia mnestica non è qualcosa
che noi conosciamo o comprendiamo o qualcosa che possiamo indicare nel cervello. Si riferisce a
qualsiasi rappresentazione, che persista nel nostro sistema nervoso di un’esperienza che può essere
ricordata». La memoria non è quindi localizzata in singole zone ma è piuttosto il risultato
dell’interazione dell’intera attività corticale. La persistenza dei ricordi lascia supporre che nella struttura
del cervello si verifichi una trasformazione duratura che prevede, secondo l’ipotesi biochimica, che nelle
cellule nervose la traccia mnestica si depositi sotto forma di particolari molecole di acido ribonucleico
(RNA) prodotto dall’attività neuronale. Questa ipotesi troverebbe conferma nella possibilità, già
sperimentata a livello di animali inferiori, di trasferire biologicamente la memoria da un individuo a un
altro. Ci sarebbe infatti un parallelismo fra trasmissione genetica della memoria e trasmissione della
memoria genetica, depositata in sequenze di DNA in grado di determinare tutte le caratteristiche di una
data specie e trasmetterle alla progenie.
L’Engramma corrisponde ad una traccia mestica e risale ad una teorizzazione proposta da R.
Semon nel 1908. Questa traccia è depositaria di un certo contenuto informativo ed è conservata nel
tessuto nervoso in forma di «neurogramma» grazie a stimoli che modificano il «substrato organico».
Quando il substrato è attivato da uno stimolo analogo è predisposto a rispondere sempre allo stesso
modo.
Semon ha ipotizzato che il complesso di tutti gli engrammi acquisiti costituisca la base della
memoria, dove tutto ciò che è stato pensato, vissuto o sperimentato si mantiene in condizioni di
disponibilità, pronto a riemergere e tornare alla memoria dietro un semplice richiamo. Si suppone che la
base neurofisiologica che permette il processo mnemonico consista in modificazioni sinaptiche sotto
forma di cambiamenti microstrutturali delle sinapsi delle cellule cerebrali. Gli engrammi sarebbero i
moduli codificati di tutte queste modificazioni sinaptiche. Quando questi moduli sono attivati portano
alla formazione di impulsi che sono copie dei moduli responsabili dell’esperienza originale, disponibili
per una decifrazione e quindi per una riutilizzazione ai fini della memoria. In questo quadro ipotetico, di
certo c’è oggi solo il ruolo importante attribuito all’acido ribonucleico (RNA) responsabile di una sintesi
specifica da cui dipende lo sviluppo sinaptico e la conseguente attivazione della memoria.
Il concetto di engramma è stato utilizzato dalla psicoanalisi e dalla psicologia analitica per
spiegare la memoria inconscia e i processi associativi.
I meccanismi di trasferimento dell’informazione al nucleo, necessari per il processo di
consolidamento, coinvolgono il sistema di trasduzione dell’AMPc ed in particolare le proteine di tipo
CREB. Il termine CREB (CyclicAMP Responsive Element Binding protein) si riferisce ad una famiglia
di fattori di trascrizione attivati dall’AMPc attraverso la PKA. In seguito ad un marcato e/o ripetuto
aumento di AMPc, la subunità catalitica della PKA trasloca nel nucleo dove fosforila la serina di CREB
permettendo che questa proteina si leghi ad una specifica regione di DNA detta CRE. Questa
interazione porta all’attivazione della trascrizione di geni cosiddetti precoci (immediate-early genes).
Alcune delle proteine sintetizzate a partire da questi geni sono fattori di trascrizione che, a loro volta,
inducono la sintesi di proteine che sono alla base delle modificazioni strutturali responsabili della
codificazione a livello sinaptico della memoria a lungo termine.
La ricerca del CREgramma si è sostituita alla ricerca dell’engramma che caratterizzava gli studi
sulla memoria degli anni ’50.
La “sede” del cosiddetto engramma è distribuita in maniera ampia ed equivalente in molti
distretti del cervello.
7. SEDI DELLA MEMORIA
Uno dei problemi più affascinanti della moderna biologia riguarda i meccanismi molecolari alla
base dell’acquisizione, dell’elaborazione e dell’immagazzinamento dell’informazione nel cervello
dell’uomo.
I processi di memorizzazione a lungo termine riconoscono a livello cerebrale aree e soprattutto
circuiti abbastanza ben definiti.
12
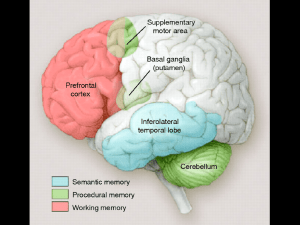
In particolare la memoria dichiarativa, archiviata nelle corteccie associative, poggia su un
insieme di strutture diencefalico - temporali appartenenti al sistema limbico: il giro del cingolo,
l’amigdala, l’ippocampo e le cortecce paraippocampali. All’interno di tale sistema la codificazione
dell’informazione avviene attraverso strette connessioni sinaptiche coinvolgenti le diverse strutture. Gli
stimoli esterni, percepiti a livello di cortecce sensitive primarie, vengono elaborati da aree corticali
associative multimodali ed integrate in mappe neuronali. Tali aree corticali proiettano ad altre strutture
corticali (giro paraippocampale e corteccia peririnale) che a loro volta attivano strutture dell’ippocampo,
in particolare la corteccia entorinale, realizzando un sistema di convergenza dell’informazione. Questa
procede quasi come in una sorta di imbuto dalla corteccia entorinale alle cellule del nucleo dentato e
culmina nell’area CA3 dell’ippocampo. Qui sono presenti neuroni dotati di ricorrenti collaterali,
strettamente connessi tra loro che realizzano - attraverso circuiti riverberanti - sinapsi (cioè punti di
contatto tra neuroni) cosiddetti forti, sede di una memoria autoassociativa. Così i diversi aspetti
dell’informazione primaria, raccolti separatamente da diverse aree corticali solo debolmente connesse
tra loro, sono “sintetizzati” a livello di un numero relativamente ridotto di neuroni che contengono
un’istantanea della “scena” iniziale colta in tempo reale. È quindi una memoria contestuale che dipende
da associazioni arbitrarie che si verificano nella stretta finestra temporale che caratterizza le
modificazioni sinaptiche, cioè circa un secondo.
L’area CA3 attiva l’area CA1 e questa, attraverso il subiculum, nuovamente la corteccia
entorinale con una proiezione questa volta divergente che compie a ritroso il percorso precedente, fino
alle sinapsi collocate nelle cortecce che per prime hanno avviato il processo. Questa stimolazione
all’indietro comincia circa 100 millisecondi dopo l’inizio e trova quindi le sinapsi, da cui il processo è
partito, ancora attive e quindi in quello stato di modificabilità sinaptica che dura circa 1 secondo. Lungo
tutto il percorso si verificheranno dunque delle modificazioni sinaptiche che determinano il formarsi di
sinapsi forti o, nei termini di Edelmenn, di selezionare un circuito secondo un processo di darwinismo
neuronale, cioè privilegiando un circuito e, quindi, un tipo di informazioni rispetto ad altre.
Un singolo neurone può appartenere contemporaneamente a differenti gruppi assemblati di
neuroni e, a seconda delle circostanze e dei segnali in arrivo, attivarsi in un gruppo o in un altro. La
possibilità di scarica neurale è incrementata dalla forza e dalla ripetitività della stimolazione in arrivo
cosicché si possono formare delle connessioni preferenziali, o facilitate.
La traccia mnesica è da considerarsi distribuita nel senso che non esiste un centro unico di
memoria, in quanto molti distretti del sistema nervoso partecipano all’immagazzinamento di una
determinata informazione; nello stesso tempo l’engramma può considerarsi localizzato, in quanto solo
determinate strutture sono implicate nella codificazione mnesica di un certo evento e ciascuna di esse
partecipa in maniera differente alla codificazione.
Si possono ipotizzare graduali reclutamenti ed una continua riorganizzazione di sistemi
neuronali per opera sia di una attività endogena che in seguito ad attività esterna tradotta dai sistemi
sensoriali, oppure grazie all’interazione di entrambi questi tipi di attività. Attraverso tale complessa serie
di eventi l’engramma matura nel tempo (probabilmente nella neocorteccia per quanto riguarda il
contenuto dichiarativo) svincolandosi da strutture sottostanti come l’ippocampo o altre regioni del lobo
temporale mediale. Questo processo spiegherebbe le forme graduali di amnesia retrograda osservate in
certe sindromi amnesiche dopo rimozione della parte mediale del lobo temporale.
Alcuni neurobiologi hanno identificato la regione del cervello chiamata cingolato anteriore come
la responsabile del recupero e della conservazione dei ricordi remoti.
La parte mediale del lobo temporale svolge una funzione chiave nel processo di
memorizzazione del contenuto dichiarativo. Il sistema limbico serve solo come serbatoio temporaneo
(parte mediale del lobo temporale) in quanto l’immagazzinamento definitivo ha luogo nella
neocorteccia. L’archiviazione e il recupero dell’informazione dichiarativa è ascrivibile all’interazione tra
siti neocorticali tra loro fisicamente separati ed il lobo temporale mediale.
Il consolidamento consisterebbe in un processo di graduale connessione funzionale delle
diverse aree che complessivamente contengono la rappresentazione dell’evento.
13
Danni al sistema limbico che includono la parte mediale del lobo temporale o strutture mediali
diencefaliche interferiscono con la funzione mnemonica dichiarativa senza intaccare apprezzabilmente il
processo di memoria procedurale.
I gangli della base e le connessioni cortico-striatali hanno un importante ruolo funzionale
nell’acquisizione e nell’immagazzinamento dei contenuti procedurali.
La corteccia cerebellare ed i nuclei cerebellari sono strutture implicate nell’acquisizione e
nell’immagazzinamento della memoria procedurale.
Le cortecce prefrontali esercitano una funzione di controllo nella codificazione e soprattutto nel
richiamo dei ricordi: consentono di mettere ordine, categorizzare, selezionare, ristabilire il giusto valore
nella memoria. D’altra parte il loro è il ruolo dei guardiani ed anche dei censori della memoria, possono
non guidare il richiamo fino in fondo o possono censurare alcuni ricordi parziali a favore di altri o
effettuare una verifica non accurata.
La memoria implica un cambiamento più o meno duraturo del rapporto sinaptico tra i neuroni,
sia attraverso una modificazione strutturale sia mediante modificazioni biochimiche.
Osservazioni sperimentali localizzano nell’ippocampo una delle sedi più importanti per i
processi di apprendimento e la memorizzazione.
Il sistema GABAergico ha un ruolo rilevante nella regolazione dei processi di apprendimento e
memorizzazione. Ciò è confermato dalla presenza di una grande quantità di recettori per il GABA
nell’amidgala, area coinvolta nei processi di consolidamento della memoria.
Il legame fra l’ippocampo e la memoria è noto agli scienziati sin dagli anni cinquanta, ma si sa
poco di come quest’associazione si manifesta nell’attività neurale.
L’ippocampo raccoglie informazioni semantiche a proposito delle parole, mentre il giro
paraippocampale richiama le informazioni sensoriali relative alle parole.
Diversi studiosi hanno identificato il modo con cui l’ippocampo contribuisce ad apprendere e a
ricordare i fatti e gli eventi che costituiscono la nostra memoria a lungo termine, o memoria
dichiarativa. Studiando l’attività dei neuroni dell’ippocampo, gli scienziati hanno capito come il cervello
segnala la formazione di nuove memorie associative, una forma di memoria dichiarativa. Ciò prova
l’elasticità dell’ippocampo in relazione all’apprendimento.
Il lobo temporale mediale è in grado di differenziare termini veri da altri falsi. Mentre
l’ippocampo raccoglie informazioni semantiche a proposito delle parole, il giro paraippocampale
richiama le informazioni sensoriali relative alle parole, come l’aspetto e la voce di chi le ha pronunciate.
8. TIPI DI MEMORIA
Nell’affrontare il tema della memoria è da tenere in considerazione come la formazione e
trasformazione del ricordo siano determinate dal fattore tempo e come i ricordi stessi assumano aspetti
peculiari in relazione alla sfera emotiva e all’attività della coscienza.
La reintegrazione è il ricordo più completo in grado di localizzare spazialmente e
temporalmente nel proprio passato l’evento ricordato.
Il riconoscimento è l’identificazione, in un’esperienza successiva, di uno stimolo
precedentemente esperito, come quando un oggetto, una persona o una situazione ci appaiono familiari
o «già visti».
Il riconoscimento facilita l’accesso a una traccia mnesica, ma non elimina del tutto la necessità
della rievocazione, inoltre il contesto ha una parte importante nel ricordo, in quanto materiali appresi in
un determinato ambiente sono meglio rievocati in quell’ambiente piuttosto che in un altro.
Anche rilevante è il modo in cui viene organizzato il materiale mnesico: è possibile conservare
una stabile capacità di recupero nel tempo se questo è ben codificato e classificato
Hebb ha intuito già nel 1949 che la memoria non è un sistema unitario, ma un insieme di
sottosistemi integrati. Sulla base di questa intuizione ha distinto una memoria a lungo da un’altra a breve
termine.
Il ruolo del tempo nel determinare le memorie è già evidente considerando che la
multifattorialità della memoria è individuata sulla base della durata della traccia mnesica stessa. Ciò
14
riporta alla distinzione di memoria a breve termine, di capacità ristretta, cui accedono ricordi fedeli, ma
di durata brevissima, inferiore ad un minuto, e di memoria a lungo termine, di capacità e durata
praticamente illimitate, che sottende a tutte le nostre conoscenze, innate ed acquisite.
Esistono, perciò, due meccanismi di immagazzinamento delle informazioni che, in entrata, si
collocano nella memoria a breve termine (MBT) dove sono soggette a una rapida dissoluzione.
restaurata dalla ripetizione che può essere di mantenimento per la sua conservazione nella memoria di
breve termine, o di integrazione nelle strutture di memoria a lungo termine (MLT).
Il materiale che sfugge alla ripetizione decade nell’oblio. La ripetizione non può aumentare la
capacità del sistema di memoria, per cui il mantenimento è possibile solo se il numero delle
informazioni in entrata è sufficientemente piccolo. Per questo si è soliti rappresentare il processo di
ripetizione come un anello che lascia la memoria a breve termine per poi ritornarvi, dove è evidente
che, se devono essere ripetute troppe informazioni, le ultime andranno perse prima che il processo di
ripetizione possa iniziare. La velocità di ripetizione ha un ritmo molto simile a quello del linguaggio
parlato.
Il riapprendimento è la ripetizione di un’esperienza acquisita un po’ di tempo dopo la prima
acquisizione.
La memoria immediata per il materiale verbale sembra basarsi su un tipo di codifica fonologico,
mentre la memoria a lungo termine sembra basata su codici semantici.
La memoria a breve termine ha una capacità di circa 5 elementi che, non sottoposti a
ripetizione, vengono subito perduti, come quando si comunica oralmente un numero telefonico. La
memoria a lungo termine è pensata invece come virtualmente illimitata, così che ogni informazione che
passa dalla MBT alla MLT, come ad esempio il proprio numero telefonico, vi troverà una collocazione
permanente. Nella MLT l’oblio non è determinato da rapido deterioramento come nella MBT, ma
dall’incompletezza dei richiami necessari alla sua identificazione, per cui risulta difficile «ripescare»
l’informazione adeguata, come nel caso delle parole «sulla punta della lingua».
Nella MBT l’oblio è determinato dal fatto che stimoli successivi hanno espulso l’informazione
che non ha potuto essere restaurata dalla ripetizione di mantenimento; nella MLT l’oblio può dipendere
o dal fatto che l’informazione non è mai stata trasmessa, o dal fatto che non sono disponibili richiami
sufficienti alla sua identificazione.
I meccanismi che favoriscono il passaggio dalla MBT alla MLT sono:
l’attenzione che, se in generale favorisce la memoria, talvolta la inibisce, come quando pensare a
ciò che è stato detto impedisce di fare attenzione a ciò che si sta dicendo;
l’elaborazione concettuale che associa le informazioni in entrata a conoscenze già acquisite,
migliorando il successivo recupero dell’informazione passata nella MLT;
le mnemotecniche che impongono un’organizzazione alle informazioni che non hanno rapporti
fra loro, utilizzando i metodi dei luoghi dove alcune collocazioni geografiche servono come
suggerimenti per ritrovare le informazioni, il metodo delle associazioni che collegano le informazioni
inserendole in una storia significativa, il metodo delle parole-chiave a cui riconnettere informazioni non
correlate, e altri espedienti che favoriscono il consolidamento, senza il quale non si ha il passaggio dalla
MBT alla MLT.
L’amnesia retrograda che si registra in soggetti che, dopo commozioni o lesioni cerebrali,
manifestano un’amnesia degli eventi immediatamente precedenti l’incidente, conferma la teoria secondo
cui l’attività neuronale deve disporre di un periodo di tempo, dopo una determinata esperienza, per il
consolidamento delle informazioni, affinché queste possano essere incamerate in maniera stabile e
definitiva nella MLT.
Una particolare memoria, quella di lavoro, si pone tra la memoria a breve e quella a lungo
termine, anche se sembra far parte della memoria a breve termine. Questo tipo di memoria trattiene “a
fuoco” le informazioni per un tempo molto breve, dell’ordine di secondi, per consentire che esse siano
analizzate, processate, confrontate da altri circuiti cerebrali: una sorta di taccuino della mente da
utilizzare per la formazione di concetti ed in ultima analisi per pensare.
La “memoria di lavoro” è, quindi, un deposito di informazioni a breve termine, consistente in
memoria che è attiva e rilevante solo per un breve periodo di tempo.
15
Pare che la capacità di divenire “intenzionali” sia legata al fatto che, istante per istante, si
compattano i singoli scenari di working memory: è possibile che la consapevolezza e, di conseguenza,
l’intenzionalità siano il prodotto della moltiplicazione working memory x tempo.
La memoria di lavoro viene interpretata come una riverberazione dello stimolo, mentre gli altri
tipi di memoria sarebbero il risultato della configurazione della matrice sinaptica, costituita dalle
caratteristiche dei collegamenti tra i neuroni.
Le procedure della memoria di lavoro hanno un ruolo di rilievo nella verifica della correttezza e
plausibilità della narrazione del ricordo.
Più recentemente è stato proposto un modello di memoria diverso da quello della memoria a
breve termine, basato sul concetto di ‘working memory’ strutturata in più componenti.
La struttura proposta si articola in almeno tre sottosistemi. Parte del sistema è costituita da un
centro di controllo dell’attenzione, il cosiddetto “central executive”, che forma un’interfaccia tra la
memoria a lungo termine e due o più sottosistemi. Uno di questi, lo “sketch pad”, o blocchetto per
appunti visuospaziale, è un sistema che può acquisire e manipolare il materiale mnesico di natura visiva
o spaziale. Questo sistema sembra essere associato a strutture dell’emisfero destro ed è specializzato nel
mantenimento dell’informazione visuospaziale, mentre l’informazione verbale è conservata utilizzando
il “loop” o circuito fonologico, che è un sistema che comprende un magazzino di informazioni
acustiche a breve termine, accoppiato ad un processo di ripetizione articolatoria.
Il centro esecutivo (central executive) è considerato il responsabile della selezione ed esecuzione
delle strategie e provvede a mantenere e spostare l’attenzione a seconda delle esigenze. Il centro svolge
anche la funzione di sistema attentivo di supervisione (SAS: supervisory attentional system), attivo
quando si deve far fronte ad un problema inaspettato, o superare“schemi” abituali inadeguati in una
situazione d’emergenza. Il SAS è anche responsabile delle attività di pianificazione e coordinamento.
La capacità di working memory è fondamentale nel definire l’intelligenza, ciò perché un gruppo
di compiti di working memory mostra punteggi altamente correlati con le più tradizionali misure
dell’intelligenza, basate su test di ragionamento.
Il centro esecutivo è anche responsabile della capacità di coordinare le informazioni provenienti
da fonti differenti.
Questo centro può essere interessato da un danno frontale, da cui deriva la cosiddetta sindrome
disesecutiva (dysexecutive syndrome), meglio nota come sindrome del lobo frontale.
Il bagaglio delle nostre conoscenze è comunque rappresentato da una memoria a lungo termine
in cui le esperienze possono essere codificate in modo consapevole o meno.
La formazione del ricordo è impedita se viene effettuato un elettroshock durante la seduta di
apprendimento, invece non c’è interferenza se l’elettroshock è indotto a distanza di ore
dall’apprendimento. Questo esperimento conferma l’esistenza di due forme temporali di memoria:
labile quella a breve termine, resistente quella a lungo termine.
Il ricordo può durare mesi, anzi una vita intera: non può basarsi su modificazioni evanescenti
ma deve essere sostenuto da modificazioni durature.
Nonostante la memoria a lungo termine sia una estensione di quella a breve termine, il
decadimento della prima ha un andamento temporale diverso dalla seconda e comporta che la memoria
non sia un processo unitario. Sono, perciò, state distinte forme temporalmente distinte di memoria:
quella “primaria” e quella “secondaria”.
La memoria primaria è molto più labile e molto più suscettibile alla inferenza retroattiva della
secondaria. Il contenuto mnemonico a breve termine è più facilmente cancellabile di quello a lungo
termine.
La trasformazione del ricordo a breve termine in quello a lungo termine necessita di un periodo
di consolidamento durante il quale l’informazione può essere facilmente perduta e distrutta da diversi
fattori disturbanti.
Una proprietà definitiva della memoria a lungo termine, che la distingue da quella a breve
termine, è la necessità della sintesi proteica in una preciso ambito temporale durante la fase
dell’apprendimento.
16
La memoria a breve termine appare, così, indipendente dalla sintesi proteica, mentre quella a
lungo termine si rivela dipendente dalla sintesi proteica.
Gli eventi responsabili della memoria consolidata avvengono nelle stesse cellule dove si
verificano quelli responsabili della memoria a breve termine attraverso un’estensione del sistema di
trasduzione dal citoplasma al nucleo.
Molte delle nostre azioni sono guidate da esperienze passate combinate con previsioni sul
futuro. Uno dei grandi obiettivi della biologia consiste nello scoprire come le cellule del cervello
possano creare una rappresentazione che si estende all’indietro e in avanti nel tempo.
La memoria esplicita o dichiarativa comprende la memoria dei fatti (o semantica) e degli eventi
(o episodica). La prima si riferisce a conoscenze generali, condivise da vasti gruppi di persone e
sostanzialmente decontestualizzate, corrisponde quindi alle nostre conoscenze “enciclopediche”, la
seconda è invece fortemente legata al contesto (chi, dove, come, quando), ha una forte componente di
soggettività e trova la sua espressione più tipica nella memoria autobiografica. È importante notare che
mentre la memoria semantica può essere espressa attraverso affermazioni, nomi, definizioni o brevi
frasi, la memoria episodica possiede intrinsecamente una componente sostanzialmente narrativa, spesso
con al centro il sé. Tale distinzione - presenza o meno di una componente narrativa - è secondo alcuni
la caratteristica principale che differenzia memoria episodica e memoria semantica.
I ricordi che vengono organizzati in base al tempo e al contesto sono chiamati memorie
episodiche (in precedenza rievocazione). Una memoria episodica è il ricordo di qualcosa senza alcun
riferimento circostanziato al proprio passato.
Lo schema di attivazione delle cellule suggerisce come l’ippocampo contribuisce alla formazione
delle memorie episodiche. Parcheggiando un’auto per recarsi al lavoro, i neuroni dell’ippocampo si
attivano secondo uno schema legato a quel luogo. Durante la permanenza sul luogo di lavoro si prende
spesso l’ascensore, ma non con l’intenzione di recarsi in parcheggio, pertanto non si attivano i ricordi
connessi al parcheggio. Quando si prende l’ascensore per tornare a casa, si attivano cellule che non
erano state coinvolte durante gli altri viaggi nell’ascensore, perché lo scopo è quello di raggiungere
l’auto.
Questa è una tipica memoria episodica con la quale verosimilmente il soggetto si incontrerà una
volta sola e con valenze soggettive: gli elementi visivi, uditivi, olfattivi, verbali registrati su aree corticali
diverse, ma simultaneamente, sono convogliati verso un unico gruppo neuronale in cui risultano
associati in un’unica situazione contestuale. Questo insieme di informazioni soggiornerà, in questa sorta
di buffer di memorie episodiche, per settimane o mesi, finché non verrà trasferito ad aree corticali con
alta densità cellulare ed eventualmente classificato ed inquadrato temporalmente. Tale trasferimento
sarà tanto più rapido quanto maggiore sarà il numero degli stimoli da memorizzare: più rapido in
ambienti ricchi di stimoli, meno in ambienti poveri, così da consentire la liberazione di circuiti che
verranno occupati da nuove informazioni.
Nell’ambito della memoria episodica, si distingue la memoria recente (che registra avvenimenti
verificatisi entro l’ultima ora) e la memoria remota (che registra fatti avvenuti più di un’ora fa).
La memoria episodica fa riferimento al sistema coinvolto nella rievocazione di particolari
esperienze o episodi. È influenzata dal livello di attenzione e organizzazione, riflette l’importanza di
questi processi per l’elaborazione di strutture di memoria che siano accessibili al recupero.
La memoria semantica riguarda la memoria del significato delle parole. A differenza della
memoria episodica, non ha carattere personale, ma è piuttosto comune a tutti coloro che parlano la
stessa lingua.
La memoria semantica sarebbe gradualmente influenzata dai processi educativi, iniziando con la
nostra conoscenza percettiva del mondo fisico intorno a noi, arrivando ad includere il linguaggio, la
nostra conoscenza della società fino a tutte le informazioni specialistiche dettagliate che acquisiamo
nell’ambito dei nostri rapporti individuali o professionali.
La memoria semantica immagazzina l’informazione che ha avuto origine in molte esperienze
separate che non sono rievocabili singolarmente. Mentre l’accesso alla memoria semantica può essere
preservato nella sindrome amnestica, l’evidenza clinica suggerisce che senza la memoria episodica, il
soggetto ha grosse difficoltà ad aggiungere nuove informazioni alla memoria semantica.
17
In sintesi si può affermare che la memoria semantica consiste nell’accumulare le informazioni
provenienti da numerosi episodi, riflette la nostra capacità di valutare globalmente l’insieme degli
episodi, enucleando e mettendo in risalto le caratteristiche che sono comuni a un gruppo di episodi
diversi, mentre la memoria episodica rappresenta la capacità di estrarre e recuperare un singolo evento
dall’insieme.
Il cervello dispone oltre che di memoria dichiarativa, di memorie implicite. Esse consentono,
per esempio, di ricordare come si va in bicicletta anche dopo anni – e si parla più precisamente di
memoria procedurale – e di fissare concetti astratti – in questo caso la memoria è detta semantica.
Anche chi ha l’ippocampo danneggiato non perde questi tipi di conoscenza, che risiedono in un’altra
regione, della corteccia cerebrale: il neopallio.
La memoria a lungo termine può essere divisa in due sottotipi: la memoria dichiarativa e la
memoria procedurale. La memoria dichiarativa o esplicita, si distingue in un tipo episodico ed in un tipo
semantico, è direttamente accessibile alla coscienza, può essere descritta “a parole” e concerne il vissuto
storico individuale (forma episodica) oppure la conoscenza del mondo in generale (forma semantica).
La memoria procedurale o implicita non è direttamente accessibile alla coscienza e non può
essere descritta “a parole” in termini di fatti, dati specifici ed eventi ben localizzati nel tempo e nello
spazio. È la memoria che si forma in modo automatico ed ampiamente inconscio tramite pratiche
motorie o percettive ripetute molte volte.
La memoria implicita e fa riferimento ad abilità motorie come danzare, praticare uno sport,
suonare uno strumento musicale o semplicemente guidare l’auto che - pur apprese coscientemente e
con esercizio ripetuto - vengono effettuate in modo automatico.
La memoria procedurale ricorda come si svolge un’azione sia fisica che mentale. La perdita della
memoria procedurale porta a difficoltà nello svolgimento delle attività quotidiane quali il vestirsi, il
lavarsi e cucinare. Essa include comportamenti che sono diventati automatici.
La memoria implicita è quella di cui si servono i classici riflessi condizionati, quella “emotiva”
per cui il soggetto senza consapevolezza attribuisca ad un evento scatenante un significato ansiogeno o
terrifico perché nella sua mente è connesso a situazioni o suoni o immagini in grado di scatenare
reazioni emotive quali ansia o paura. L’apprendimento implicito consiste in ciò che viene appreso
inconsapevolmente, attraverso messaggi cosiddetti subliminali, e che produce comportamenti
consequenziali.
La memoria non dichiarativa è il prodotto di un gruppo di sistemi di apprendimento che sono
in grado di accumulare informazioni, ma non di estrarre dall’insieme e identificare episodi specifici. È
un tipo di memoria specializzato nell’accumulare informazioni provenienti dalla realtà circostante, ma
non è in grado di individuare e tenere separati i singoli episodi.
Per recuperare ed organizzare un ricordo ci si può servire di
Priming, ovvero della capacità di identificare una parola in condizioni di rumore o di riprodurre
la parola quando viene loro proposta la radice o parte di questa.
Apprendimento procedurale, cioè acquisizione di abilità motorie (come guidare una macchina) o
cognitive (capacità di problem solving).
Condizionamento associativo, consistente nel comportarsi sulla base di un’esperienza appresa
senza riuscire a stabilire un collegamento con il ricordo dell’esperienza.
Condizionamento valutativo, ovvero dell’influenza dell’esperienza pregressa sulla valutazione
dell’importanza dello stimolo.
9. APPRENDIMENTO
L’apprendimento è un cambiamento permanente che si manifesta attraverso processi adattativi
del repertorio comportamentale dovuto all’esperienza.
L’apprendimento comporta una “variazioni del potenziale conoscitivo” dell’individuo. Può
essere definito anche come una conoscenza organizzata che si struttura sempre meglio: per imparare è
necessario avere già un bagaglio di conoscenze.
18
In anni recenti è stato provato che il cervello adulto è in grado di riorganizzare e ottimizzare la
sua struttura e le relative connessioni nervose.
È stato possibile ottimizzare il trasferimento di informazione tra neuroni comunicanti per
mezzo di brevi, intensi stimoli elettrici e misurare quantitativamente questo miglioramento, che si
riflette in una migliore risposte da parte del neurone. Questo test, detto Potenziamento a Lungo
Termine (LTP), è stato usato come unità di misura della capacità dei neuroni di adattarsi agli stimoli
ambientali, che è un equivalente dell’apprendimento.
I processi di apprendimento e di memorizzazione si possono riassumere in quattro fasi:
acquisizione ed immagazzinamento della memoria a breve termine, che ha una durata massima
di un giorno; in precedenza era definita come registrazione, ovvero l’impressione nel sistema nervoso
dell’evento percettivo, stadio in cui l’informazione entra nell’organismo rimanendo a disposizione un
tempo proporzionale al tempo di esposizione.
consolidamento, che sembra coinvolgere strutture quali l’ippocampo e l’amigdala; ipotizza
l’esistenza di un processo nervoso perseverativo che porta a un consolidamento della traccia mnesica.
Ciò significa che, finché questo processo non è compiuto, la traccia mnesica non è completamente
consolidata, per cui può essere cancellata in tutto o in parte da eventi o trattamenti che non sono più
efficaci quando il processo di consolidamento è concluso.
immagazzinamento (o ritenzione) in engrammi a lungo termine: consiste
nell’immagazzinamento dell’informazione sensoriale, detto anche registro sensoriale, che dà ai sistemi di
estrazione delle caratteristiche e ai sistemi di riconoscimento di configurazioni il tempo (compreso tra
0,25-2 secondi) di operare, sui segnali in arrivo, scelte precise in ordine agli input rilevanti da
discriminare da quelli inutili. Infatti l’enorme quantità di informazione presente in un’immagine
sensoriale non è generalmente importante per interpretare il significato, per cui si rende necessaria una
selezione ritentiva.
richiamo delle informazioni acquisite: il recupero, o ecforia, secondo R. Semon, è il richiamo alla
mente del materiale precedentemente immagazzinato nel registro sensoriale con la ritenzione e,
ipoteticamente consolidato, reso disponibile per l’effettivo ricordo. Il recupero dei ricordi non è un
processo semplice che segue la legge del tutto o nulla. È possibile dimenticare qualcosa, ma conservare
intorno ad essa informazioni che ne rendono possibile il recupero, come nel fenomeno delle parole
«sulla punta della lingua», dove i vocaboli che vengono in mente al loro posto hanno caratteristiche in
comune con la parola «dimenticata». Il recupero è inoltre facilitato dalle immagini mentali che
visualizzano l’azione, dagli schemi di organizzazione del materiale da memorizzare, dalla frequente
ripetizione che compie opera di consolidamento, e dal sovrapprendimento al di là della semplice
acquisizione del materiale.
Nella fase di apprendimento la regione temporale mediale potrebbe rapportarsi con i siti di
immagazzinamento della memoria localizzati altrove nel cervello.
Nell’apprendimento complesso l’informazione è immagazzinata anche nella neocorteccia
inferotemporale.
Il cervello ha un ruolo fondamentale nella fase di apprendimento dell’assuefazione a lungo
termine di un riflesso, mentre il cervelletto è implicato nella memoria motoria, nel condizionamento di
un riflesso e nelle modificazioni plastiche cui può andare incontro.
Sistemi neuronali, determinanti il comportamento e la percezione sensoriale, quando si
modificano, possono alterare la potenzialità alla reazione piuttosto che la reazione immediata allo
stimolo. La durata dell’apprendimento abbraccia un ambito temporale compreso tra pochi secondi ed
un arco di vita. La memoria è la ritenzione ed il recupero di tale rappresentazione interna.
L’apprendimento di memorie semantiche in genere si verifica come un processo di
decontestualizzazione e generalizzazione di memorie episodiche.
La memoria episodica consente di creare un legame associativo tra situazioni che si verificano
contemporaneamente nell’arco di tempo utile per la modificabilità sinaptica.
Perché la memorizzazione si verifichi sono necessari dei rinforzi esterni rappresentati dallo stato
emotivo e dal livello di attenzione.
19
Secondo il modello di Damasio, aree corticali diverse sono reciprocamente connesse con zone
cosiddette di convergenza che, a loro volta, inviano e ricevono impulsi da una zona di convergenza
localizzata nell’ippocampo. L’attivazione di un punto qualsiasi di questo circuito comporterà
l’accensione di tutto il sistema.
Una serie di eventi esterni può determinare reazioni emotive di tipo somatico (variazioni del
ritmo cardiaco o del respiro, pallore, rossore, sudorazione). Tali stati vengono registrati nel talamo, che
è connesso al nucleo dell’amigdala, a sua volta collegata con l’ipotalamo e quindi con la regolazione
endocrina e neurovegetativa. L’amigdala - considerata il centro della vita emotiva - è a sua volta
collegata bidirezionalmente all’ippocampo. Pertanto un determinato evento verrà memorizzato anche
con la sua coloritura emotiva, che dà all’evento un valore di base esclusivamente somatico aderente alle
esigenze di adattività e, in ultima analisi, di sopravvivenza. Nel ricordo riprodotto vi sarà anche il
significato in termini somatici, positivi o negativi, dell’evento, senza che ci sia sempre consapevolezza
del motivo di tale reazione emotiva.
La detta modificazione dello stato corporeo viene registrata come un cambiamento del sé.
Le vie neuronali considerate sono condizionate da meccanismi di punizione e di gratificazione
che possono intervenire nella codificazione della traccia mnesica e quindi nell’apprendimento. È questo
il motivo per cui luoghi emotivamente accattivanti facilitano la memorizzazione.
La risposta emotiva, infatti, non solo esprime il valore dell’evento, ma funziona anche come
propulsore di attenzione e memoria operativa. Un altro fattore che contribuisce alla registrazione
dell’evento è, infatti, lo stato di vigilanza e di attenzione, attivato da vie di proiezione dal setto mediale
all’ippocampo che sarebbero anche in grado di aumentare il tempo di modificabilità sinaptica
consentendo una memorizzazione più estesa e profonda.
La contestualizzazione associativa del ricordo - legata alla simultaneità della situazione - fa sì che
questo possa essere evocato nel suo insieme da qualsiasi particolare purché facente parte della scena
memorizzata
Questo può verificarsi perché la presentazione del frammento di ricordo attiva i neuroni della
area ippocampale che proietteranno all’indietro l’impulso fino alle aree corticali che per prime avevano
registrato l’evento, dove verrà ripristinata l’immagine primitiva (somma dell’evento stesso, del suo
valore biologico e del cambiamento dello stato del sé che ha determinato).
Saranno interessate solo queste aree corticali perché per queste era stato selezionato un circuito
preferenziale attraverso la formazione di sinapsi forti. D’altra parte la attivazione, spontanea o
intenzionale, di tale percorso a ritroso comporta ulteriori modificazioni di sinapsi che nel tempo
avranno comunque subito ulteriori modificazioni.
Il frammento di ricordo stimola il recupero automatico dell’evento, un atto riflesso che avviene
senza sequenziazione temporale, senza controllo di plausibilità o consistenza con altri elementi, senza
intelligenza, ma probabilmente con fantasia. Attraverso questo processo, definito di ecforia, uno
stimolo, un’immagine, un colore, un profumo, una musica, possono attivare una serie di ricordi a
catena, apparentemente scollegati tra loro.
Questo fenomeno è evidente in alcuni pazienti amnesici, dove la narrazione del ricordo può
assumere aspetti bizzarri o del tutto implausibili. Le libere associazioni fanno riferimento ad elementi
autobiografici quasi “semantizzati” (infermiere > feriti > PS > dimissione) per l’emergere evidente di
circuiti forti e possono assumere o meno un carattere di plausibilità.
Tali narrazioni appaiono quindi come storie inventate seguendo l’immaginazione, cioè
combinando tra loro particolari assunti da tutt’altra parte.
Elementi di nonsense, assurdità, bizzarria sono presenti in racconti fantastici dedicati
soprattutto ai bambini nei quali, per l’immaturità di determinate aree cerebrali, è frequente tale ecforia
riflessa e disordinata, per quanto non priva di significati.
La narrazione del ricordo può svilupparsi in modo ordinato, sequenziale temporalmente e
logicamente plausibile attraverso una serie di processazioni che vedono coinvolte essenzialmente le aree
prefrontali, in particolare quelle ventromediane, attraverso la memoria di lavoro. Questa opera iniziando
il recupero del ricordo in modo intenzionale, selezionando il metodo di recupero più adatto,
individuando frammenti strategici che per confronti successivi guidano il processo fino alla
20
individuazione del ricordo parziale associativo. Attraverso il richiamo di informazioni codificate nella
memoria a lungo termine e il confronto con l’obiettivo, i ricordi vengono organizzati temporalmente
(quando?) e nei contenuti (chi? come? dove? perché?). In questo senso la componente
episodico/narrativa della memoria si distingue da quella semantica/non narrativa.
L’individuazione delle procedure di richiamo che intervengono nella costruzione della
narrazione del ricordo è stata possibile analizzando le modalità con cui individui normali rievocano. Si
può procedere da un inizio generico fi no al recupero successivo, più o meno intenzionale, di dettagli
ulteriori, oppure ci si può accontentare di memorie incomplete ritenendole complete, o perché se ne è
perduta l’informazione o perché il loro valore emotivo non era forte, o ancora perché non sono
accessibili in quel momento. Inoltre la narrazione può essere falsata da memorie rese forti dall’abitudine
e, in un certo senso, di tipo semantico o dalla intensa componente emotiva che ne ha accompagnato la
codificazione. Infine possono intervenire fenomeni deduttivi di tipo inferenziale.
Il richiamo di eventi passati fornisce alla corteccia informazioni utili per edificare nuove
rappresentazioni nelle aree associative. Ogni richiamo quindi modifica le sinapsi e di conseguenza la
rappresentazione corticale del ricordo stesso.
Pertanto il ricordo è modificato dagli eventi sinaptici intrinseci alla rievocazione stessa e dagli
eventi - del mondo esterno e conseguentemente sinaptici - accaduti successivamente. Come già Freud
aveva intuito, le tracce mnesiche possono assumere un diverso significato a partire da eventi successivi.
Il recupero dell’informazione complessiva avviene per attivazione di un circuito forte,
selezionato dalla stimolazione retrograda di una rete già attiva. È possibile comunque che - data l’alta
possibilità di accesso alle informazioni contenute nell’ippocampo - il frammento di ricordo attivi anche
circuiti diversi, forse meno ripetuti, ma con maggior valore emotivo.
Il sonno ha almeno due diversi effetti sull’apprendimento: innanzitutto consolida i ricordi,
proteggendoli da interferenze successive o dal deterioramento, inoltre sembra essere in grado di
‘recuperare’ o di ripristinare i ricordi.
Estese regioni del cervello sono coinvolte nell’elaborazione dei ricordi durante una particolare
forma di sonno, detta a onda lenta.
Il sonno consente il consolidamento delle memorie da poco acquisite. In questo processo il
ruolo più importante è per il sonno a onda lenta, profondo e senza sogni, rispetto a quello REM (rapid
eye movement), associato con i sogni.
Il sonno ha, inoltre, un impatto importante nel migliorare le capacità delle persone di imparare
le lingue.
L’abilità degli studenti di ricordare parole straniere viene migliorata dal sonno, anche se gli
studenti sembravano aver dimenticato parte di quello che avevano appreso durante il giorno
precedente.
Molto probabilmente durante il sonno il proprio deposito originario di ricordi viene «sfruttato»
in una sintesi incontrollata (o dovuta a stimoli subconsci) di RNA nelle matrici genetiche. Questa
rassegna notturna dei registri genetici suscita naturalmente una folla di associazioni nella corteccia
cerebrale, cioè i sogni.
Siccome durante il sonno le informazioni non vengono fissate mediante una sintesi successiva
di proteine, le visioni dei sogni scompaiono con la dissoluzione degli acidi nucleici, cioè dopo venti o
trenta minuti. Così si ricordano soltanto gli ultimi minuti di un sogno, a meno che non ci si svegli
durante la notte, fissando così una parte delle ultime immagini sognate che possono allora essere
rievocate anche la mattina seguente.
L’immagazzinamento di tipo olografico rende possibile la creatività: dalle interrelazioni del gran
numero di immagini conservate, l’una sopra l’altra, tutte quasi eguali eppure un po’ diverse, risulta il
processo creativo che si esprime sempre di nuovo in particolari varianti. Creatività, intuizione ed idee
non hanno una determinata sede nel cervello, ma nascono dall’interazione che è propria
dell’immagazzinamento multiplo di percezioni esterne ed interne, o meglio, non sono altro che questa
interazione.
21
10. ALTERAZIONI DELLA MEMORIA
L’ansia da esame, il terrore d’una interrogazione improvvisa, o il black-out del candidato in un
quiz televisivo avviene quando si verificano eventi associati a paura o eccitazione. Questi sono dei
sentimenti collegati ad avvenimenti materiali. Percezioni insolute o associate al pericolo, oppure ricordi
spiacevoli provocano, tramite il diencefalo e i gangli del simpatico, una stimolazione diretta delle
ghiandole surrenali e di certe regioni cerebrali, le quali in pochi centesimi di secondo immettono in
circolo due ormoni: l’adrenalina e la noradrenalina, note come ormoni dello stress, con la funzione di
preparare il corpo all’attacco o alla fuga immediata, aumentando immediatamente la pressione
sanguigna e il metabolismo degli zuccheri e dei grassi.
Però ben dentro al cervello questi ormoni dello stress esercitano anche un’altra funzione:
influiscono sui collegamenti tra i neuroni.
Le sinapsi cioè si devono attivare. E proprio questo processo può essere disturbato e persino
impedito dagli ormoni dello stress.
La noradrenalina stessa, per esempio, è una sostanza trasmettitrice, soprattutto per le sinapsi
inibenti, e inoltre agisce come antagonista biochimico di un’altra importante sostanza trasmettitrice,
l’acetilcolina, di cui impedisce il rifornimento. Tutto questo è un blocco naturale causato dal
meccanismo dello stress, il quale, come vedremo ancora, deve in ogni caso essere interpretato alla luce
dell’istinto di conservazione, considerando che ogni «riflessione» ritarderebbe lo scatto che salva dal
nemico. La natura non poteva prevedere che la nostra società moderna collegasse proprio reazioni di
stress e di allarme all’apprendimento e al pensiero, cioè a processi in cui non hanno niente a che fare.
Nel pensiero, nell'apprendimento e nell’oblio, oltre a questi disturbi di breve durata ve ne sono
anche di cronici. Anche queste irregolarità, sia la lentezza nell’apprendimento che l’incapacità di
cambiare un comportamento appreso, possiamo metterlo in relazione con l’equilibrio ormonali delle
surreni, in particolare con un altro gruppo di ormoni di questo sistema: l’idrocortisone e il rispettivo
ormone stimolante ipofisario, il cosiddetto ormone andrenocorticotrpico (ACTH).
La mancanza o l’eccesso di ACTH disturbano l’apprendimento: contenuti nuovi
dell’apprendimento vengono acquisiti con maggior difficoltà ad alto livello di ACTH; questo ormone,
quindi, è necessario per una conservazione rapida e intensiva, ma un eccesso irrigidisce i contenuti
appresi, impedendo così l’acquisizione di nuove informazioni.
La memoria remota può essere inaccessibile in caso di problemi di rievocazione, mentre quella
più recente può essere definitivamente perduta quando la una traccia mnesica non si è consolidata o
non è stata adeguatamente immagazzinata.
Le alterazioni della memoria sono distinte in quantitative e qualitative.
Le alterazioni quantitative sono le ipermnesie, le ipomnesie e le amnesie.
L’ipermnesia è un aumento delle capacità mnestiche che si ha in due configurazioni:
a) L’ipermnesia permanente che comprende sia le capacità mnesiche globali che si rivelano al di
sopra della media, sia quelle circoscritte ad alcuni temi particolari come cifre, date, poesie. Tale
ipertrofia della memoria non corrisponde necessariamente ad una intelligenza superiore; infatti la
possiedono anche alcuni soggetti con deficienze mentali, i cosiddetti «idiots savants».
b) L’ipermnesia transitoria che consiste nella capacità di attingere a materiali solitamente
inaccessibili alla coscienza. Può verificarsi in casi di trauma o stress emotivi, durante stati febbrili intensi
o di elevata tossicità, in caso di lesioni cerebrali, crisi epilettiche o maniacali, nonché durante terapia
ipnotica o psicoanalitica.
Le ipermnesie sono costituite da un aumento quantitativo delle capacità mnemoniche e si
distinguono in permanenti e transitorie. Le ipermnesie permanenti, di natura non patologica, si
osservano in persone molto dotate sul piano intellettivo; talora sono limitate ad argomenti specifici.
Eccezionalmente possono essere presenti anche in soggetti oligofrenici.
Le ipermnesie transitorie sono per lo più connesse ad intensi stati emotivi, nell’ipomania,
oppure a condizioni di alterata coscienza, come stati crepuscolari (isterici ed epilettici) o ancora in
situazioni di catastrofe o di grande paura con rischio della vita.
L’Ipomnesia è un diffuso e progressivo indebolimento delle capacità mnesiche.
22
È spesso associata a fenomeni di impoverimento del substrato organico cerebrale, legato all’età,
a patologie psichiche e neurologiche, a sofferenza su base ipossica o tossica.
La memoria a breve termine si riduce spesso a causa dell’età e di malattie come la schizofrenia e
il morbo di Parkinson, consente agli individui di ricordare informazioni per brevi periodi, durante il
quale vengono valutate al fine di determinare un’azione.
La paramnesia è un’alterazione qualitativa della memoria dove i ricordi vengono deformati nel
loro contenuto, nel loro significato e nella loro localizzazione spazio-temporale, in base alla
psicopatologia in atto al momento dell’elaborazione. Oltre alla confabulazione, che nella varietà delle
sue manifestazioni presenta tratti paramnestici, le figure più significative di paramnesia sono:
La reminiscenza, che può essere definita una rievocazione senza riconoscimento, come nel caso
della criptomnesia, caratterizzata dalla riemersione di un ricordo scambiato per un’idea completamente
nuova.
La pseudoreminiscenza che è una rievocazione errata, immaginaria o riferentesi a un’esperienza
psichica realmente vissuta in passato ma ricordata con un contenuto, un significato o una collocazione
diversi da quelli reali (allomnesia), oppure attribuente caratteristiche di ricordo ad esperienze mai vissute
(pseudomnesia).
L’ecmnesia per cui il soggetto vive come se fossero attuali ricordi della sua infanzia. Il
fenomeno compare nella demenza e nell’alterazione dello stato di coscienza dovuta a lesioni cerebrali o
all’assunzione di sostanze allucinogene. Questa reviviscenza della memoria, che però sfugge al controllo
della coscienza, rende incerta la classificazione psicopatologica del fenomeno tra le forme di ipermnesia
e paramnesia. Esiste anche chi classifica il fenomeno tra i processi di depersonalizzazione.
L’esperienza di déjà vu in cui una situazione nuova viene percepita come «già vista». Tale
esperienza, detta anche falso riconoscimento, è spesso vissuta con la sensazione di sapere cosa accadrà
dopo. Fenomeni di déjà vu sono frequentemente descritti da soggetti epilettici e isterici per i quali
l’esperienza può durare ore o giorni, mentre negli individui normali permane al massimo qualche
secondo. La sensazione di familiarità del déjà vu può essere spiegata: come un’errata generalizzazione di
esperienze passate applicata a una situazione che risulta parzialmente somigliante; come la
perseverazione di uno stato emotivo precedentemente causato da una difficoltà di adattamento al
contesto presente; come il ricordo di fantasie inconsce riattivato dalla situazione attuale. Simili al déjà vu
sono il déjà entendu con la sensazione di aver già sentito ciò che si sta udendo al momento, il déjà fait
in cui il soggetto è convinto che quello che gli sta succedendo gli sia già accaduto in precedenza, il déjà
pensé come sensazione di aver già formulato in passato lo stesso pensiero.
L’esperienza di jamais vu, detta anche misconoscimento, che è un’esperienza in cui il soggetto
percepisce situazioni a lui ben note come nuove ed estranee. Può verificarsi nelle schizofrenie, in alcune
crisi epilettiche, in stati di tossicità o di particolare affaticamento. Rientrano in questo campo la
persuasione che individui o cose abbiano subito repentini cambiamenti o vere e proprie trasfigurazioni,
da uomo a donna, da giovane a vecchio, o abbiano cambiato radicalmente fisionomia.
11. AMNESIA E STATI DISSOCIATIVI
L’amnesia è una riduzione più meno grave della capacità di ricordare. Può avere una causa
organica o psicogena. Nel primo caso le amnesie differiscono a seconda delle strutture cerebrali
compromesse per cause traumatiche, tossiche, metaboliche, infiammatorie o degenerative. L’entità del
disturbo dipende dalla sede, dall’estensione, e dall’eventuale irreversibilità. Caratteristica del disturbo
amnestico è, come vuole la legge formulata da T. A. Ribot, di preservare, contrariamente al discorso
della memorizzazione, i dati acquisiti in un lontano passato, lasciando decadere per primi quelli di
recente acquisizione. Il disturbo può essere circoscritto a un determinato periodo di tempo e allora parla
di amnesia lacunare che, in base al tipo di lacuna, può essere semplice, sistematizzata o selettiva.
L’amnesia lacunare può essere inoltre transitoria o irreversibile. Se invece il disturbo è diffuso con
ricordi imprecisi sia per eventi recenti sia per eventi remoti che colpiscono la registrazione, la ritenzione
e il recupero, allora si assiste a una modificazione del comportamento e della personalità. L’amnesia può
23
essere retrograda se riguarda fatti precedenti l’insorgenza dell’evento amnesiogeno, anterograda se
riguarda i fatti successivi, retroanterograda se investe entrambi i periodi.
I soggetti con un Disturbo Amnestico presentano compromessa la loro capacità di apprendere
nuove informazioni, o sono incapaci di ricordare informazioni apprese precedentemente o eventi
passati.
Un danno cerebrale può compromettere la memoria producendo difficoltà nella acquisizione di
nuove informazioni e perdita delle informazioni già acquisite.
Le cause dell’amnesia possono essere molteplici e possono essere il risultato di un intervento
chirurgico sul lobo temporale, d’intossicazione cronica da alcool, traumi cranici, encefaliti, insufficiente
ossigenazione, tumori e disturbi vascolari.
Il disturbo amnestico persistente è dovuto ad abuso di sedativi (ipnotici o ansiolitici), o di
alcolici. All’abuso di alcol è correlata una tipica forma amnestica classicamente descritta come sindrome
di Korsakoff. In questa patologia, riscontrabile in alcolisti cronici, sono presenti difficoltà sia a
memorizzare nuovi eventi, sia amnesia per gli avvenimenti lontani nel tempo, precedenti all’instaurarsi
della malattia. In questa forma morbosa i fenomeni di confabulazione hanno un significato
compensatorio. Il soggetto elabora falsi ricordi per coprire la propria amnesia e rispondere in qualche
modo alle esigenze sociali della situazione in cui si trova.
Il deficit di memoria prodotto dalla somministrazione di medicinali antipsicotici può essere
interrotto attraverso la stimolazione con un farmaco D1-agonista, che agisce stimolando selettivamente
questi recettori. Il farmaco si è dimostrato efficace nel ripristinare la funzione di memoria a breve
termine nei primati sottoposti ai test sperimentali.
Nella sindrome di Korsakoff sono di frequente riscontro zone di atrofia corticale a carico dei
lobi frontali e lesioni del nucleo talamico.
I disturbi mnesici rappresentano il sintomo patognomonico in quelle sindromi caratterizzate da
un declino delle funzioni cognitive, come le demenze, dove processi degenerativi primari o secondari
del cervello rappresentano il fattore determinante per una diminuzione di quelle funzioni, come
l’attenzione e l’apprendimento, che sono essenziali per un buon funzionamento della memoria.
Le amnesie organiche, classificate in base alle strutture cerebrali compromesse, si distinguono
in:
amnesie assiali dove la lesione interessa il Sistema reticolare ascendente e il sistema libico,
indispensabili per la memorizzazione e l’utilizzazione delle informazioni. Si manifesta con amnesia di
fissazione caratterizzata dall’incapacità di apprendere dati nuovi per i quali, dopo pochi minuti, non
rimane alcun ricordo; evocazione mediocre del passato con disturbo della narrazione; adattamento
discreto al presente per quel tanto che ha a che fare con situazioni familiari da lungo tempo apprese;
amnesie corticali con distruzione parziale o totale dell’immagazzinamento mnestico
rappresentato dagli engrammi della corteccia dei due emisferi e drastica riduzione della possibilità di
formare nuove associazioni. Le lesioni che distruggono una parte dell’emisfero non determinano
un’amnesia di fissazione, ma turbe del linguaggio, dell’acquisizione ed esecuzione dei gesti,
dell’organizzazione del comportamento;
amnesie globali sono diffuse con gradi diversi e comportano non solo una riduzione delle
possibilità mnestiche, ma anche una progressiva alterazione e diminuzione del patrimonio cognitivo. Le
amnesie globali, oltre alla compromissione della memoria, investono tutte le capacità acquisite dal
soggetto, linguaggio e gestualità compresi.
Nell’amnesia le strutture lese contribuiscono all’immagazzinamento ma non intervengono
nell’acquisizione né trattengono in “archivio” l’informazione.
Le sindromi amnesiche sono correlate a processi patologici situati in strutture ascrivibili al
circuito limbico (la parte mediale del lobo temporale o le porzioni mediali del diencefalo).
La amnesia anterograda dipende dalla disfunzione di una delle due regioni del cervello che
corrispondono alla superficie mediale dei lobi temporali e all’area diencefalica mediana.
L’amnesia anterograda comporta una incapacità ad acquisire e registrare nuove informazioni,
quella retrograda potrebbe essere definita amnesia di rievocazione, per l’incapacità di riportare alla
coscienza engrammi fissati precedentemente.
24
La amnesia retrograda e la amnesia anterograda sarebbero il risultato di un danno alla regione
temporale mediale. Dopo un congruo periodo di tempo, le memorie diventano indipendenti dalla
regione temporale mediale e questa area del cervello non è più necessaria per la conservazione della
memoria o per il processo di rievocazione.
Inoltre l’amnesia può essere generalizzata o semplice, a secondo se interessa tutti i ricordi,
oppure se si riferisce a un determinato materiale mnemonico; si definisce sistematizzata, quando
vengono cancellati tutti i ricordi riguardanti determinati fatti o persone.
Lesioni bilaterali causano una amnesia globale, lesioni dell’emisfero sinistro causano deficit a
carico della memoria di materiale verbale, quelle dell’emisfero destro a carico della memoria di materiale
visuo-spaziale non verbale.
Le amnesie organiche transitorie dipendono da alterazioni organiche passeggere in cui i disturbi
della memoria, essendo associati a confusioni mentali e a compromissioni della vigilanza, rendono
difficile un’esatta valutazione dei processi mnestici. L’ictus amnestico si manifesta in modo improvviso
con un’amnesia di fissazione e rapido oblio di ciò che è stato detto o fatto. Dopo la crisi, che dura
diverse ore, il soggetto recupera improvvisamente o progressivamente le sue capacità mnestiche,
mantenendo un vuoto di memoria per il periodo critico;
Se il danno neuro-degenerativo è lieve, il riapprendimento delle informazioni memorizzate ne
migliora il riconoscimento. Oltre un certo livello di danno, quando l’apprendimento diventa deteriorato,
riapprendere informazioni precedentemente memorizzate può essere controproducente.
Le amnesie senili sono alterazioni fisiologiche che si manifestano con difficoltà ad evocare nomi
propri anche familiari, reperire oggetti che rientrano nella normale consuetudine, difficoltà di acquisire
nuovi dati se non rientrano nei temi del proprio passato interesse. Quasi annullate sono l’acquisizione di
nuove tecniche di pensiero e nuove strategie mentali. Il processo prende avvio in modo significativo a
partire dal quarto decennio con una progressione che risulta inferiore nei soggetti che, per il loro buon
livello intellettuale, sono in grado di acquisire nuove informazioni nel loro campo di interesse personale.
In soggetti con la classica sindrome amnestica è la memoria esplicita a essere elettivamente
compromessa mentre quella implicita risulta conservata.
Benché molti episodi di amnesia funzionale facciano seguito a un trauma cranico, Le amnesie
psicogene sono prodotte da disturbi affettivi in soggetti con strutture cerebrali indenni.
L’amnesia funzionale che compromette la memoria autobiografica è un segno patognomonico
di un gruppo di disturbi mentali noti come disturbi dissociativi.
L’amnesia dissociativa è contraddistinta dall’incapacità di ricordare importanti informazioni
riguardanti la propria storia personale, in particolare un’esperienza traumatica.
Esiste un meccanismo che permette di sopprimere consciamente alcuni ricordi. Freud suggerì
l’esistenza di questo meccanismo un secolo fa.
Vecchi ricordi e memorie terrorizzanti possono far sì che si dimentichino più facilmente le
informazioni appena apprese. I medici hanno notato che quando si subisce un’esperienza che suscita
paura, la si ricorda per almeno un quarto della normale durata della vita. Inoltre, se a primati viene fatta
ricordare ripetutamente una situazione angosciante, essi mostrano comportamenti simili a quelli degli
esseri umani affetti da Disturbo Post Traumatico da Stress: evitano tutto ciò che è associato con
l’esperienza originale, e le loro memorie emotive a lungo termine interferiscono con la capacità di
ricordare informazioni appena apprese.
Freud ha individuato la causa del disturbo amnestico di origine psichica nella rimozione di
particolari eventi di natura affettiva o conflittuale a scopo difensivo. La soppressione della
rappresentazione cosciente dell’evento o del conflitto non comporta la soppressione dell’affetto ad essa
legato che si sposta su un’altra rappresentazione in modo che gli elementi rimossi non siano accessibili a
tutto ciò che potrebbe attivarli. Questa inattivazione non è però assoluta, come dimostra il trattamento
analitico in grado di far emergere il rimosso. Rifiutando l’interpretazione basata sull’immaturità
funzionale, Freud scorge nell’amnesia infantile una rimozione riguardante la sessualità che si estende alla
quasi totalità degli eventi dell’infanzia e che si risolve col declino del complesso edipico e con l’inizio
dell’età della latenza. Nell’amnesia infantile Freud vede inoltre la condizione delle amnesie successive e,
in particolare, quelle dell’amnesia isterica che è un sintomo nevrotico per cui si dimentica quello che
25
non si «vuole» ricordare perché incompatibile con il concetto che uno ha di sé, e riguarda un periodo di
tempo circoscritto o un episodio particolare.
Possono essere distinte tre forme di amnesia psicogena:
sistematizzata, che riguarda solo avvenimenti specifici;
localizzata, che riguarda un periodo di ore o di settimane;
generalizzata, che si riferisce a tutta la vita del soggetto.
In quest’ultimo caso, l’amnesia si sovrappone ovviamente alla fuga, che può essere di tre tipi:
l’amnesia con cambiamento di identità;
l’amnesia con perdita di identità;
La maggior parte delle perdite psicogene di memoria autobiografica sono associate con qualche
alterazione dell’identità.
L’amnesia dissociativa (su base psicogena) comporta l’incapacità di ricordare importanti notizie
personali; è di solito di origine traumatica o stressogena, e risulta troppo estesa per essere spiegata con
una normale tendenza a dimenticare. Il disturbo comporta una menomazione reversibile della memoria,
in cui i ricordi personali non possono essere recuperati in forma verbale.
Solitamente l’Amnesia Dissociativa si presenta come una lacuna, o una serie di lacune, riportate
retrospettivamente, nella rievocazione di momenti della storia della vita di un individuo. Queste lacune
sono di solito legate a eventi traumatici o estremamente stressanti.
La caratteristica fondamentale dell’amnesia dissociativa è la perdita della memoria episodica,
ossia della capacità del soggetto di rievocare particolari avvenimenti che appartengono alla sua
esperienza personale; la fuga dissociativa comprende l’amnesia, ma comporta anche un cambiamento
dell’identità.
La fuga dissociativa è caratterizzata dalla confusione sulla propria identità, o dalla perdita
dell’identità, o dall’assunzione di un’identità nuova. Questo significa arrivare a dimenticare chi si è, non
solo quello che si è fatto.
Il disturbo dissociativo dell’identità assomiglia alla fuga, ma in questo caso il passaggio da
un’identità all’altra e fra i corrispondenti sistemi di ricordi autobiografici è ciclico.
Ciascuna di queste identità (qualche volta indicate come alter ego) ha il proprio magazzino di
ricordi autobiografici, che si riferiscono ad eventi ed esperienze che si sono verificati quando quella
identità era dominante. Infine, gli alter ego sono separati da un’amnesia simmetrica o asimmetrica, che
impedisce a una personalità di conoscere le azioni, le esperienze o perfino l’esistenza dell’altra.
L’amnesia funzionale è presente anche in altre forme di disturbi dissociativi.
Un particolare tipo di amnesia dissociativa è nota come “sindrome di Ganser”, tipica di soggetti
reclusi. Nel disturbo sono spesso presenti risposte inesatte e approssimative alle domande, disfunzioni
sessuali, menomazione nelle attività, nelle relazioni interpersonali, auto-mutilazione o impulsi aggressivi,
impulsi e gesti suicidiari.
L’amnesia acuta può risolversi spontaneamente allorché il soggetto viene allontanato dalle
situazioni traumatiche a cui l’amnesia era collegata; alcuni individui con amnesia cronica possono
prendere gradualmente a rievocare i ricordi dissociati; altri soggetti possono sviluppare una forma
cronica di amnesia. I soggetti che hanno avuto un episodio di Amnesia Dissociativa possono inoltre
risultare predisposti a sviluppare amnesia in circostanze traumatiche successive.
Le immagini oniriche sono ricavate in gran parte dalle esperienze recenti, pertanto persone
colpite da amnesia in modo permanente, in cui si rilevano danni nella regione cerebrale chiamata
ippocampo, pur riuscendo a trattenere informazioni per qualche minuto, subito dopo le dimenticano e
non sono sostanzialmente in grado di aggiungere quanto appreso recentemente alla propria memoria
dichiarativa o a quella episodica. Sono, infatti, compromesse sia la cosiddetta memoria dichiarativa - che
permette di dichiarare una conoscenza, per esempio ricordare un nome – sia la memoria episodica –
che consente di fissare le esperienze vissute.
I disturbi qualitativi della memoria comprendono le allomnesie, le pseudomnesie e le
criptoamnesie.
Le allomnesie sono illusioni della memoria, cioè distorsioni o deformazioni di engrammi
precedentemente registrati. L’incompletezza e l’ambiguità rievocative sono generalmente in rapporto al
26
tono affettivo ed a particolari contenuti di pensiero. Sono riscontrabili nei disturbi dell’umore con
rievocazione del passato in chiave pessimistica o nei disturbi psicotici ma possono osservarsi anche in
condizioni normali.
Le Pseudomnesie sono “allucinazioni della memoria”: comprendono i falsi riconoscimenti, cioè
riconoscere come ricordo una situazione mai sperimentata (déjà vu o del déjà vécu considerati come
alterazioni dell’affettività associata con la memoria) ed i falsi ricordi, produzioni compensatorie di fatti
immaginari e fantastici in soggetti con gravi lacune mnemoniche (confabulazione, falsificazione della
memoria in rapporto con un disturbo amnestico di origine organica, distinguibile in Confabulazione di
imbarazzo, in cui il soggetto cerca di mascherare il suo vuoto di memoria di cui il soggetto si rende
conto, e Confabulazione fantastica, in cui il soggetto che ha un vuoto di memoria descrive esperienze
avventurose e fantastiche). La confabulazione fantastica per deterioramento organico da abuso di alcool
e nel disturbo amnestico, su base organica ma non indotto da abuso di sostanze psicoattive.
Caratteristica della confabulazione è la notevole suggestionabilità, che dipende
dall’obnubilamento della coscienza, dall’indebolimento delle capacità di giudizio e dall’intervento della
fantasia: è una condizione molto somigliante ai sogni ad occhi aperti. Una tipo di confabulazione
fantastica con contenuti persecutori si verifica in stato di coscienza lucida, quando il paziente si
convince che altre persone vogliano danneggiarlo, derubarlo o di defraudarlo, e produce falsi ricordi di
natura fantastica per giustificare tale convinzione.
La criptoamnesia, un disturbo psicogeno, consiste nel vissuto di non riconoscere il ricordo
come tale, perché questo perde il suo carattere mnesico specifico e viene esposto come fatto o idea
attuale.
12. LA MEMORIA DELL’ANZIANO
Non è infrequente che, invecchiando, capiti di ricordare chiaramente avvenimenti che non sono
mai accaduti, o che si sono svolti in modo diverso rispetto a quanto è rimasto impresso nella memoria.
Gli anziani hanno una maggiore propensione a ricordare eventi mai successi.
Il soggetto anziano manifesta un fisiologico deficit della capacità di memorizzare fatti recenti,
mentre è solitamente meno compromessa la memoria a lungo termine (fenomeno noto come legge di
Ribot).
L’anziano normale presenterebbe le seguenti funzioni cerebrali deficitarie:
velocità di apprendimento, di elaborazione di nuove informazioni, dei tempi di reazione a
stimoli complessi;
memoria ed apprendimento;
funzioni sensoriali (in particolare la pallestesia);
forza muscolare;
postura;
deambulazione.
La funzione mnesica si caratterizza per una riduzione delle capacità di fissazione e, in misura
minore, di rievocazione; il linguaggio risulta alterato per una perdita dell’efficacia espressiva con
tendenza a ridurre soprattutto l’uso di aggettivi; si riduce la capacità di orientamento temporo-spaziale e
la capacità psicomotoria di reazione, a livello intellettivo si riduce la capacità di risolvere quesiti
complessi e di apprendere in tempo limitato nuove informazioni; a livello comportamentale si osserva
tendenza alla depressione, labilità emotiva e comportamento stereotipato.
L’anziano ha l’impressione di essere espulso e di vivere al di fuori della vita sociale e culturale,
maturando sentimenti di isolamento e di inutilità. La sua memoria ha valore soltanto come fenomeno
soggettivo, senza alcuna legittimazione sociale, e non è più la memoria dispensatrice di norme e di
regole per il gruppo di appartenenza.
L’anziano, isolato e quasi fuori del tempo, in preda ad un sentimento di inadeguatezza e di
inutilità, spesso indulge alla nostalgia ed alla apologia del tempo passato.
Il vecchio diventa depositario della memoria. Ma non subisce passivamente l’impronta del
tempo, del passato, lo ricostruisce invece in un rapporto dinamico e creativo che rappresenta il
27
momento distintivo del “fare memoria”. Nella rievocazione del vecchio gli avvenimenti trascendono la
dimensione di un riferimento rigorosamente storico, di eventi temporalmente dati e cronologicamente
riguardati, per entrare piuttosto nella forma del racconto storico.
Gli uomini anziani con elevati livelli di testosterone nel sangue hanno una migliore memoria
visiva e verbale e possono svolgere meglio compiti spaziali. Potrebbe esserci una modulazione
ormonale delle capacità cognitive durante l’invecchiamento.
Nei soggetti anziani,è noto, il deterioramento mentale può essere secondario a disordini
metabolici, malattie cerebrovascolari, farmaci, intossicazioni, lesioni intracraniche occupanti spazio,
come, anche, ad infezioni croniche. Il decadimento cognitivo, associato a cause trattabili, rappresenta
circa il 10-20% di tutte le cause note di demenza.
Il processo che si instaura è una correlazione tra perdita di terminali presinaptici, degenerazione
neuronale e diminuzione della performance cognitiva e comportamentale.
Avere livelli più alti di testosterone libero è associato con una riduzione del rischio di certi tipi di
perdita di memoria. A differenza della forma legata, il testosterone libero può circolare nel cervello e
influenzare i neuroni. Il testosterone potrebbe, però, aumentare il rischio di tumori alla prostata
sconsigliando, così, l’utilizzo terapeutico. Inoltre stimola la produzione di un numero eccessivo di
globuli rossi, addensando il sangue e contribuendo al rischio di ictus.
Molti anziani sviluppano una riduzione di tolleranza al glucosio, che impedisce al corpo di
muovere il glucosio dal sangue alle cellule che ne fanno uso. Test cognitivi, radiologici e di laboratorio
indicano che soggetti con livelli di zucchero particolarmente elevati hanno peggiori prestazioni nel test
di memoria. Inoltre, il loro ippocampo è più piccolo.
13. DEMENZA
Caratteristica comune nei soggetti più anziani risulta, com’è noto, la diminuzione o le alterazioni
della memoria, e quelle delle funzioni cognitive, spesso limitate ad un grado modesto che consente di
mantenere un buon adattamento alla vita privata ed a quella sociale. In tale caso il deterioramento
funzionale viene in genere considerato come un’espressione dell’invecchiamento cerebrale fisiologico. I
problemi iniziano quando la modesta menomazione iniziale progredisce fino ad un vero stato
demenziale. Il modo di prevenire tale pericolosa eventualità,ed il momento più opportuno per
intervenire, sono ancora sconosciuti e rappresentano un’importante materia di studio
nell’interpretazione del processo di senescenza. Le alterazioni della cognizione, che si associano
all’invecchiamento, coinvolgono funzioni diverse (memoria, rapidità dei processi mentali, attenzione,
linguaggio ed attività visive e spaziali ) e passano da un semplice stato di riduzione mnemonica,
attraverso un deterioramento cognitivo di grado moderato, allo stato demenziale conclamato.
Un aumento della glicemia è risultato associato a disturbi dell’umore (per lo più tristezza ed
ansia), della memoria e della capacità di attenzione.
Possibili cause di demenza trattabile comprendono l’ipotiroidismo, l’intossicazione da farmaci, i
tumori, l’idrocefalo normoteso, l’ematoma subdurale, la depressione.
La demenza è un disturbo acquisito e con base organica delle funzioni intellettive che sono state
in precedenza acquisite: memoria (a breve e lungo termine) e almeno una tra pensiero astratto, capacità
critica, linguaggio, orientamento spazio temporale, con conservazione dello stato di coscienza vigile.
Attacchi epilettici possono provocare danni cerebrali e perdita di cellule nervose, in particolare,
nelle aree dell’ippocampo che presiedono ai meccanismi della memoria, fattori ambientali possano avere
effetti benefici sul recupero delle capacità cognitive.
Anche in un cervello danneggiato, l’arricchimento ambientale migliora il recupero o la
compensazione, ma non è distinguibile l’effetto sul danno istologico indotto dagli attacchi epilettici,
nemmeno a livello di alterazione nella perdita di cellule o di sinapsi.
Queste evidenze suggeriscono che le alterazioni ambientali abbiano un significativo peso
biologico sulle funzioni cerebrali: modificare il contesto di vita di pazienti con danni cerebrali potrebbe
diventare un’efficace strategia di intervento.
28
Nel complesso è affetto da demenza oggi circa il 5% della popolazione over 65 anni, ma
addirittura il 30% degli over 85.
Con l’invecchiamento, spesso i livelli di zucchero nel sangue salgono lentamente. In casi
estremi, ciò può essere causa di diabete. Ma livelli di zucchero anche solo moderatamente alti possono
provocare problemi di memoria e restringere l’ippocampo, la parte del cervello fondamentale per
l’immagazzinamento di nuove informazioni.
Clinicamente si distinguono demenze corticali e sottocorticali:
demenze corticali: (rappresentate soprattutto dalla malattia di Alzheimer) con estesa atrofia
corticale, precoci alterazioni della memoria e successivamente perdita del pensiero astratto, agnosie,
afasia, aprassia.
demenze sottocorticali: più precoce rallentamento dei processi cognitivi, meno perdita della
memoria, ma alterazioni della personalità con apatia e rallentamento motorio.
Una distinzione può basarsi su un piano eziopatogenetico:
Demenze primarie tra cui malattia di Alzheimer, malattia di Pick, afasia progressiva primaria e
altre forme più rare.
Demenze associate a degenerazione neuronale primitiva:malattia di Parkinson, corea di
Huntington, altre forme di sindromi extrapiramidali
Demenza vascolare
Demenze da prioni
Demenza da disturbi endocrino-metabolici (patologie tiroidee, epatiche, da insufficienza renale)
Demenza carenziale (sindrome di Korsakoff-Wernicke, pellagra, deficit di B12 e folati)
Demenza da encefalopatie tossiche
Demenza da malattie organiche di varia natura(tumori, traumi, sclerosi multipla)
Demenza da infezioni (AIDS, meningiti, malattia di Whipple)
Si ritiene che poco più della metà dei dementi sia affetto dalla malattia di Alzheimer, il 15% circa
da demenza su base vascolare (demenza vascolare), il 15% da forme miste ed il restante 15% da forme
di varia natura, tossica, traumatica, tumorale, infettiva, da idrocefalo normoteso.
LA DEMENZA DI ALZHEIMER
La malattia di Alzheimer colpisce la memoria e le funzioni mentali (ad es. del pensare, del
parlare, ecc.), ma può anche causare altri problemi, quali confusione, cambiamenti di umore e
disorientamento spazio-temporale.
I primi sintomi sono rappresentati dalla difficoltà a ricordare e la perdita delle capacità
intellettive, possono essere così lievi da passare inosservati, sia all’interessato che ai familiari e agli amici.
Ma, col progredire della malattia, i sintomi diventano sempre più percettibili, e cominciano a interferire
col lavoro di routine e con le attività sociali.
È triste constatare che la nostra memoria “va a pezzi”, ma non è detto che per tutti sia così. I
problemi cognitivi associati all’Alzheimer potrebbero essere meglio concettualizzati riferendosi a un
deterioramento dell’attenzione, anziché a una perdita della memoria in senso tradizionale. Recenti prove
neurologiche legano la malattia di Alzheimer a disturbi fisiologici nei lobi frontali del cervello, dove
avviene l’elaborazione di gran parte dell’informazione, incluso il coordinamento delle funzioni relative
all’attenzione
I pazienti di Alzheimer allo stadio iniziale, di solito, non sono in grado di immagazzinare nuovi
ricordi a lungo termine. La briostatina migliora questa capacità stimolando la produzione di proteine
essenziali per la memoria a lungo termine (aumenta la sintesi delle proteine che consolidano i ricordi a
lungo termine).
Infatti, l’incapacità dei pazienti di Alzheimer di ricordare eventi di pochi giorni prima potrebbe
essere legata alla carenza delle proteine che rafforzano i punti di contatto, o sinapsi, fra i neuroni del
cervello.
Molti ricercatori sono convinti che, ben lungi da avere una singola causa, la malattia di
Alzheimer sia solitamente causata dalla combinazione di più fattori di rischio. I nuovi studi, in effetti,
29
dimostrano che la distruzione delle proteine della memoria può richiedere l’interazione di differenti
molecole che promuovono la malattia. Alcuni ricercatori hanno scoperto che le proteine possono essere
spazzate via non soltanto da elevati livelli di beta-amiloidi, ma anche da bassi livelli di beta-amiloidi in
combinazione con alti livelli di attività di Fyn.
Beta-amiloidi e Fyn, così, possono collaborare per dare origine ai cambiamenti cerebrali
caratteristici dell’Alzheimer.
In un numero estremamente limitato di famiglie, la malattia di Alzheimer si presenta col
carattere di malattia genetica dominante.
È stato scoperto un legame tra malattia di Alzheimer e cromosoma 21. Poiché la sindrome di
Down è causata da un’anomalia di questo cromosoma, i soggetti affetti da sindrome di Down hanno
maggiori probabilità di sviluppare il malattia di Alzheimer.
Una volta raggiunti i 40 anni di età, quasi tutti i pazienti con la sindrome di Down esibiscono le
caratteristiche formazioni cerebrali del morbo di Alzheimer, anche se non tutti sviluppano la demenza.
Gli adulti con la sindrome di Down presentano nella regione ippocampale del cervello una
concentrazione di mioinositolo significativamente più alta del normale. Questo incremento è associato
con una ridotta capacità cognitiva. Se fosse possibile ridurre terapeuticamente la concentrazione di
mioinositolo potrebbe curarsi questa condizione.
Una persona che ha subito un violento colpo alla testa possa essere a rischio di sviluppare la
malattia di Alzheimer. Il rischio è maggiore se al momento del colpo la persona ha più di cinquant'anni
e ha perso conoscenza subito dopo il colpo.
I pazienti che soffrono del morbo di Alzheimer rimangono capaci di una specifica forma di
memoria usata per l’apprendimento meccanico delle cose, anche se i loro ricordi di cose e persone
vengono cancellati. La scoperta suggerisce nuove strategie in grado di migliorare l’addestramento e i
programmi riabilitativi per conservare le funzioni cognitivi dei pazienti di Alzheimer e anche degli
anziani sani.
Sembrerebbe che nei malati di Alzheimer un certo numero di sistemi cerebrali sia più intatto di
quanto si pensasse. Fornendo ai pazienti gli stimoli e le istruzioni corrette, si potrebbero aiutarli ad
usare in modo ottimale questi sistemi, migliorando così le loro funzioni.
Nonostante la malattia, determinati processi mnesici che in teoria dipenderebbero dalle regioni
cerebrali danneggiate, rimangono fondamentalmente intatti.
In futuro, in base a studi recenti, potrebbe essere possibile invertire gli effetti della perdita di
memoria disattivando un transgene che causa i sintomi dell’Alzheimer, persino nei pazienti che hanno
perso tessuto cerebrale o neurale.
Il rischio di contrarre la malattia a un certo punto della vita può riguardare chiunque. Tuttavia
un gene che si trova nel cromosoma 19, ed è responsabile della produzione di una proteina chiamata
apolipoproteina E (ApoE) può influenzare questo rischio.
Alcune evidenze segnalano il ruolo cruciale svolto dal sistema colinergico nella demenza di
Alzheimer tramite sia i recettori nicotinici che muscarinici. Il sottotipo m1 postsinaptico, localizzato
prevalentemente a livello della corteccia, è indubbiamente implicato nei processi mnemonici.
L’esperienza clinica del trattamento della demenza di Alzheimer con agonisti ha dato risultati
contrastanti, poiché solo un numero limitato di pazienti risponde alla terapia migliorando le funzioni
cognitive e comportamentali. Il ruolo eccitatorio dei recettori m1 ed m2, combinato con la loro
localizzazione a livello della corteccia e dell’ippocampo, fa di questi siti importanti bersagli terapeutici
per il trattamento farmacologico dei deficit cognitivi. Un recente studio ha evidenziato una riduzione
dei sottotipi m1 e m2 ed un aumento del m4 nella demenza di Alzheimer.
Una ricerca ha documentato uno dei fenomeni molecolari più precoci che sono responsabili del
neurodegenerazione cerebrale, cioè della formazione di una sostanza responsabile delle prime perdite di
memoria legate alla malattia di Alzheimer. Si tratta dell’ammasso nel cervello di peptidi beta amiloidi,
sostanze che si accumulano intorno alle cellule e danneggiano la memoria prima della comparsa delle
placche, normalmente associate al disturbo. Tali assembramenti danneggiano la memoria
indipendentemente dalle placche o dalla morte di neuroni e, in specifiche circostanze, possono
innescare una serie di eventi che portano all’Alzheimer. Per questo persone che rischiano di ammalarsi
30
del morbo di Alzheimer iniziano ad avere disturbi della memoria anche molti anni prima dell’inizio dei
sintomi clinici più gravi.
Gli studiosi hanno ipotizzato che causa del calo di memoria fosse una sostanza presente nel
cervello e che questa fosse presente prima della morte cellulare. Per verificare questa ipotesi hanno
utilizzato topi il cui profilo genico era stato manipolato affinché gli animali sviluppassero una perdita di
memoria simile a quella osservata nelle persone prima di sviluppare il morbo di Alzheimer. Utilizzando
quindi i topi con i primi segni di disturbo ma senza placche e senza morte cellulare nel cervello hanno
scoperto una forma della proteina beta - amiloide distinta da quella che forma le placche. La prova
dell’influenza della sostanza è arrivata quando questa è stata estratta dai topi malati, purificata e iniettata
in animali sani, che a loro volta hanno mostrato i primi sintomi di demenza.
14. CONCLUSIONI
La memoria è un processo complesso, che permette di utilizzare l’esperienza per accumulare
conoscenza da utilizzare in termini maturativi sia sul piano psichico che cognitivo.
L’influenza dei fattori psicologici sulla memoria è ormai riconosciuta ed acquisita. Il ricordo del
vissuto è la chiave d’accesso all’inconscio nell’esperienza psicoanalitica.
La memoria non è consultabile in forma completamente automatica, ma è in gran parte
inconsapelvomente associata alle percezioni interne od esterne.
Anche l’aspetto cognitivo è di grande importanza nel permettere alla memoria di svolgere la sua
funzione: il materiale da memorizzare deve essere trasformato in impulsi elettrici capaci di modificare il
trasferimento di neurotrasmettori cerebrali in particolari circuiti neuronali. La traccia mnesica se è
ritenuta rilevante comporta l’attivazione di agenti di trasferimento su un substrato organico protidico e
modificazioni strutturali di connessioni interneuronali.
Il processo mnesico può essere alterato per motivi di ordine psichico e fisico.
Dr. Giuseppe Giunta
Psichiatra
BIBLIOGRAFIA (IN ITALIANO )
Baddeley A., La memoria. Laterza, Bari, 1984.
Bartleit F. C., La memoria. Angeli, Milano, 1974.
Bergson H., Materia e memoria. in Opere 1889 - 1896, Mondadori, Milano, 1986.
Bracco L., Ricerca: I processi della memoria. L’Alzheimer, Firenze, 3, 1998, www.italz.it
Cornoldi C., Apprendimento e memoria nell’uomo. Utet Libreria, Torino, 1986.
Dawkjns R, Il gene egoista. Zanichelli, Bologna, 1979.
Ebbinghaus H., La memoria. Zanichelli, Bologna, 1975.
Freud S., Al di là del principio di piacere. In Opere, Boringhieri, Torino, vol. IX, 1977.
Freud S., La rimozione. In Metapsicologia. In Opere, Boringhieri, Torino, vol. VIII, 1976.
Freud S., Le neuropsicosi da difesa. In Opere, Boringhieri, Torino, vol. II, 1968.
Freud S., Le origini della psicoanalisi. Lettere a Wilhelm Fliess 1887-1902. Boringhieri, Torino, 1968.
Freud S., Progetto di una psicologia. In Opere, Boringhieri, Torino, 1968, vol. II.
Freud S., Psicopatologia della vita quotidiana. In Opere, Boringhieri, Torino, vol. IV, 1970.
Freud S., Studi sull’isteria (in collaborazione con Josef Breuer). In Opere, Boringhieri, Torino, vol. I, 1967.
Gala C., Chiesa S., Depressione e invecchiamento. In: Gala C. (Ed.): Psicogeriatria. Masson, Milano, 1990.
Galimberti U, Dizionario di Psicologia. Utet, Torino, 1994.
Giberti F., Psicoterapia e senilità. Minerva psicogeriatrica, 1, 1986.
Goldberg E., Il paradosso della saggezza, Ponte Alle Grazie, Milano, 2005.
Hebb D.O., L’organizzazione del comportamento. Franco Angeli, Milano 1975.
Hilgard E. R., Atkinson R. C., Atklnson R. L, Psicologia. Giunti - Barbera, Firenze, 1984.
Jung C. G., Tipi psicologici. In Opere, Boringhieri, Torino, vol. VI, 1969.
31
Lurija A. R., Neuropsicologia della memoria. Armando, Roma, 1981.
Lurija A. R., Viaggio nella mente di un uomo che non dimenticava nulla. Armando, Roma, 1979.
Musatti C., Trattato di psicoanalisi. Boringhieri, Torino, 1949.
Ravizza L., memoria. In G. Cassano (a cura di) Trattato Italiano di Psichiatria, Masson, Milano, 1993.
Ravizza L., Torta R. (Eds.), La Memoria, Le Memorie. Dalla memoria cellulare alla memoria sociale. Cortina
Ed., Torino, 1989.
Robustelli F., La memoria. in Nuove questioni di psicologia, La Scuola, Brescia, 1972.
Roncato S., Apprendimento e memoria. il Mulino, Bologna, 1982.
Sigala S., Cinelli A. et Al., Sistemi dopaminergici e memoria: aspetti neurochimici e farmacologici. In: Ravizza L.,
Torta R. (Eds.): La memoria, le memorie. Dalla memoria cellulare alla memoria sociale. Cortina, Torino,
1992.
Spinnler H. B., Sterzi A., Vallar G., Le amnesie. Angeli, Milano, 1977.
Tanzi E., I fatti e le induzioni nell’odierna istologia del sistema nervoso. Rivista Sperimentale di Freniatria, 19,
1983.
Vester F., Il pensiero, l’apprendimento e la memoria. ed. Giunti, Firenze, 1987.
Viale R. (a cura di), Mente umana, mente artificiale. Feltrinelli, Milano, 1989.
Zazzetta S., Un ambiente che cura. www.dica33.it
32