Dipartimento di Psicologia
Facoltà di Medicina e Psicologia
Corso di laurea Magistrale in Neuroscienze Cognitive e
Riabilitazione Psicologica
Tesi di Laurea:
RAPPRESENTAZIONE CORPOREA: UNO STUDIO
fMRI IN SOGGETTI CON ALTERAZIONI
PERIFERICHE
Relatore:
Prof.ssa Cecilia Guariglia
Correlatore:
Prof. Gaspare Galati
Laureanda:
Propato Filomena
Matr. 1232336
Anno Accademico 2014/2015
INTRODUZIONE……………………………………………………………………………………….1
CAPITOLO I: La Rappresentazione Mentale Del Corpo. ................................................................ 3
1.1 Origine del concetto scientifico della rappresentazione corporea. ................................................. 3
1.2 Tassonomia diadica: schema corporeo e immagine corporea. ....................................................... 4
1.3 Tassonomia triadica della rappresentazione corporea.. ................................................................. 8
1.4 Plasticità Cerebrale e rappresentazione corporea......................................................................... 13
CAPITOLO II: Elaborazione Sensorimotoria In Pazienti Con Deafferentazione Periferica. ..... 16
2.1 Aree cerebrali selettive per il corpo. ............................................................................................ 16
2.2 Meccanismi Top-Down e Bottom-Up della rappresentazione corporea. ..................................... 19
2.3 Simulazione mentale di azioni motorie in pazienti amputati. ...................................................... 21
2.4 Cambiamenti strutturali in seguito a cambiamenti periferici. ...................................................... 24
CAPITOLO III: Rappresentazione Corporea: Uno Studio fMRI In Soggetti Con Alterazioni
Periferiche. ........................................................................................................................................ 28
3.1 Introduzione ................................................................................................................................. 28
3.2 Metodologia ................................................................................................................................. 31
Soggetti ........................................................................................................................................ 31
Stimoli .......................................................................................................................................... 31
Test ............................................................................................................................................... 33
Procedura...................................................................................................................................... 34
Acquisizione delle immagini ........................................................................................................ 34
Preprocessamento delle immagini ................................................................................................ 35
3.3 Analisi dei test carta e matita. ...................................................................................................... 35
3.4 Analisi dei dati comportamentali. ................................................................................................ 35
3.5 Analisi dei dati fMRI ................................................................................................................... 36
3.6 Risultati ........................................................................................................................................ 37
T-Test nei compiti carta e matita ................................................................................................. 37
Anova: Accuratezza ..................................................................................................................... 37
Anova: Tempi di risposta. ............................................................................................................ 39
Aree attivate durante il compito di rotazione mentale. ................................................................ 40
Voxel Based Morphometry .......................................................................................................... 42
3.7 Discussione. ................................................................................................................................. 44
CONCLUSIONI .................................................................................................................................... 49
BIBLIOGRAFIA ................................................................................................................................... 51
RINGRAZIAMENTI…………………………………………………………………………………...60
Introduzione
L’immagine corporea è “il quadro mentale che ci facciamo del nostro corpo, vale a dire il modo in
cui il nostro corpo appare a noi stessi” (Shilder, 1935). Nel sistema nervoso centrale esistono
diverse rappresentazioni del corpo utili per la percezione della posizione degli arti nello spazio, la
localizzazione di stimoli cutanei, la programmazione di movimenti e per la coscienza del proprio
corpo. La percezione ed il controllo del proprio corpo richiede però l'integrazione di informazioni
provenienti da più canali sensoriali e relative a più parti del corpo. A tal fine è necessario che
informazioni somatosensoriali, visive, propriocettive, acustiche e vestibolari siano integrate in
rappresentazioni più complesse, di natura multisensoriale (Tessari et al., 2010). Quindi, mediante
informazioni top-down e bottom-up viene a crearsi una rappresentazione del corpo astratta. Ma cosa
succederebbe alla nostra rappresentazione se avvenisse un cambiamento improvviso conseguente
alla perdita di informazioni derivanti dalla periferia del nostro corpo? Le aree deputate
all’elaborazione dello schema corporeo sono sovrapponibili tra pazienti amputati e soggetti sani? La
perdita di informazioni sensorimotorie comporta una modifica della struttura cerebrale? L’obiettivo
del presente studio è quello di chiarire le modificazioni a livello funzionale e strutturale in pazienti
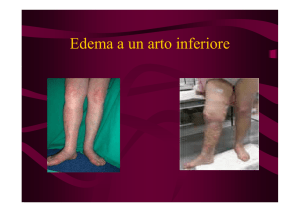
con amputazione di arto inferiore.
Nel primo capitolo verrà presentata una descrizione dell’origine del concetto di
rappresentazione corporea seguita da un dibattito sul binomio “schema corporeo - immagine
corporea” e sulla difficoltà nel sopraggiungere ad un’unica definizione di questi due concetti
utilizzati da diversi autori in modo intercambiabile (Macdonald Critchley, 1979). Successivamente
verrà esposto il concetto di tassonomia triadica della rappresentazione del corpo sottolineando
l’indipendenza funzionale di ogni rappresentazione corporea; in seguito saranno descritti casi in cui
mediante cambiamenti fisici e/o conflitti multisensoriali, il nostro cervello va incontro a processi di
riorganizzazione e riprogrammazione.
Nel secondo capitolo verranno discusse le aree cerebrali che codificano in modo selettivo il
nostro corpo e i relativi studi; inoltre verrà discussa l’influenza della perdita di informazioni topdown e bottom-up sulla mappa visuo-spaziale del corpo (mediante il sub test di rievocazione frontecorpo di Daurat-Hmeljiak et al. 1978) in pazienti con danno cerebrale unilaterale e pazienti con
amputazione di un arto (Palermo et al., 2014). In seguito saranno presentate diverse prove
sperimentali per indagare l’efficacia della simulazione mentale dell’azione in soggetti amputati. In
1
particolare sarà presentato uno studio di rotazione mentale di mani (Nico et al., 2004) in cui i
soggetti con amputazione si mostreranno più lenti e meno accurati rispetto al gruppo di controllo.
Un altro studio condotto da Curtze et al. (2010) su pazienti amputati e un paziente con chirurgia
plastica rotazionale, mostrerà un’assenza di effetto di lateralità rispetto ai controlli e una riduzione
dei tempi di risposta; successivamente saranno presentate le varie ipotesi su questi risultati. Infine,
in questo capitolo, verranno discussi i cambiamenti strutturali in seguito a deafferentazione
periferica mediante un’analisi VBM in soggetti amputati con dolore da arto fantasma lieve e medioalto (Preißler et al., 2013). I risultati dello studio mostreranno l’influenza di questi due fattori
sull’aumento e la riduzione di materia grigia nelle strutture della corteccia cerebrale.
L’ultimo capitolo verterà sulla nostra ricerca e verranno introdotte le ipotesi, il metodo,
materiali, i processi relativi all’acquisizione e al processamento delle immagini fMRI. Verranno poi
esposte le analisi condotte mediante neuroimaging funzionale e analisi Voxel Based Morphometry
(VBM). La prima utilizzata per indagare i pattern di attivazione di pazienti con amputazione di arto
inferiore durante un compito di rotazione mentale di parti corporee e non; la seconda per indagare
se la perdita di informazioni sensorimotorie potesse portare ad una modificazione strutturale della
corteccia. Saranno infine presentati e discussi i risultati della presente ricerca.
2
CAPITOLO I
La rappresentazione mentale del corpo.
1.1 Origine del concetto scientifico della rappresentazione corporea.
L’interesse scientifico per le rappresentazioni corporee risale al XVI secolo, quando un chirurgo
francese, Ambroise Paré, migliorando le tecniche di amputazione, evitò che i pazienti andassero
incontro ad un destino infausto. Egli notò in soldati provenienti dai campi di battaglia europei, in
seguito all’amputazione di un arto, l’insorgere di manifestazioni anomale riguardanti la parte del
corpo mancante; diede una prima descrizione medica dei sintomi, consistente in sensazioni tattili,
spesso anche dolorose, provenienti dall’arto asportato.
Dall’inizio di questo secolo il corpo come rappresentazione mentale incomincia ad avere
una rilevanza sempre maggiore, diventando oggetto di studio di varie discipline. Fino alla fine del
XIX secolo la consapevolezza corporea è stata concepita come un insieme di sensazioni corporee
interne (Frederique de Vignemont, 2010). Bonnier, nella sua pubblicazione L’Aschématie (1905), fu
il primo a introdurre il termine “schema” con riferimento all’organizzazione spaziale delle
sensazioni del corpo. Il neurologo definì l’aschematia come un disturbo della rappresentazione
topografica del corpo, basata principalmente su sistemi di informazione spaziale, di forma,
posizione del corpo nello spazio e di posizionamento delle parti del corpo, come ad esempio
l’orientamento e la postura. Su questa linea di pensiero, molti neurologi del tempo accettarono
l’esistenza di rappresentazioni mentali del corpo, spesso chiamate “schema” o “immagine”
corporea.
Tuttavia, lo studio di questa raffigurazione mentale nel corso degli anni non si è presentata
priva di difficoltà, a causa della mancanza di un’unica definizione del concetto. Vi è stata una
confusione diffusa sulla natura e le proprietà delle nozioni di schema e immagine del corpo
(Gallagher, 1986). Una situazione inevitabile data la varietà di modi che abbiamo di rapportarci al
nostro corpo, non si tratta solo di percezione, ma anche di come riusciamo a costruire una
rappresentazione durevole, che si realizza grazie alle esperienze passate e presenti attraverso il tatto,
3
la vista, la propriocezione, il comportamento motorio, la comprensione semantica e anche per
mezzo dell’influenza derivante dalle esperienze affettive, dalle esigenze biologiche e dai molteplici
disturbi della consapevolezza corporea (De Vignemount, 2010).
1.2 Tassonomia diadica: Schema corporeo e Immagine corporea.
Lo studio di come il cervello rappresenta il proprio corpo è stata oggetto di interesse già dal
secolo scorso, ma nonostante i vari riferimenti alle due rappresentazioni di schema e immagine
corporea, gli autori non sono riusciti a giungere ad un consenso univoco.
Il binomio costituito da schema corporeo e immagine corporea, non ha dunque subito nel
corso degli anni chiarificazioni per giungere ad una netta distinzione tra le due definizioni tanto che,
spesso, assumono lo stesso significato nei lavori di autori diversi. Due neurologi britannici, Head e
Holmes (1911-1912), sono stati i primi a descrivere pazienti con deficit relativi alla
rappresentazione, alla localizzazione e alle sensazioni del corpo. Da queste osservazioni hanno
ipotizzato che tali deficit fossero dovuti ad un malfunzionamento della rappresentazione posturale e
hanno tentato di affrontare l’importante problema di come il cervello sia in grado di elaborare il
proprio corpo attraverso esperienze tattili e propriocettive, proponendo uno schema corporeo per la
valorizzazione della postura o del movimento passivo e un altro schema corporeo che rende
possibile la localizzazione delle sensazioni tattili applicate alla superficie corporea. In questi termini
la rappresentazione mentale del corpo viene concepita come un modello plastico di se stessi in cui
vengono valutate tutte le successive modifiche della postura, del movimento e della stimolazione
tattile (Giovanni Berlucchi, Salvatore M. Aglioti, 2009).
Macdonald Critchley (1979) volge una critica ai due autori a causa del loro “linguaggio
poco chiaro” che ha fatto sì che i termini di immagine, schema corporeo e immagine di sé, fossero
usati in letteratura in modo intercambiabile creando una confusione maggiore.
Schilder (1923-1935), in accordo con Head, equipara il modello posturale con la sensazione
cosciente della posizione corporea, ovvero, equipara lo schema corporeo, come definito da Head,
con l'immagine o rappresentazione del proprio corpo che formiamo nella nostra mente (Schilder,
1935, p.11). Merleau-Ponty (1945-1962) infine, utilizza il termine immagine corporea per designare
un’esperienza e un funzionamento dinamico del corpo che agisce nel suo ambiente. Questa
rappresentazione include una consapevolezza marginale del corpo e funziona come un sistema
4
dinamico che appartiene ad una sfera di abitudine piuttosto che ad una scelta consapevole (MerleauPonty,1945-1962, pp.141/142).
Nasce la necessità di coniare un termine onnicomprensivo capace di integrare sia le
componenti concettuali che le componenti percettive della rappresentazione mentale del corpo.
Un primo tentativo di chiarimento fu realizzato da Shaun Gallagher (1986), che distinse la
consapevolezza conscia del proprio corpo e la sua rappresentazione inconscia. Gallagher considera
la prima come base dell’immagine corporea, la cui consapevolezza è limitata solo a certe situazioni,
come ad esempio la fatica, l’eccitamento sessuale, situazioni stressanti o cambiamenti fisici come
avviene negli atleti. Inoltre, a volte, il corpo viene concepito come un pensiero, un oggetto, un peso
o un fastidio (Plügge, 1967). Con il termine pre-intenzionalità, Gallagher(1986) intende il corpo
normalmente impegnato nel mondo, allorché la consapevolezza è occupata in compiti o processi di
pensiero, ovvero, quando il corpo non è esplicitamente un oggetto della coscienza. Non si è
consapevoli del proprio corpo fino a quando non vi è un riflesso volontario o un riflesso forzato
causato dal dolore, dal disagio, dal piacere, dalla fatica. Inoltre, quando l’attenzione conscia è
diretta al nostro corpo, avviene di solito una discriminazione o isolamento della funzionalità fisica,
definito dalla circostanza. In tale consapevolezza il corpo diventa disintegrato, coscientemente
articolato in parti, sebbene la caratteristica fisica o parte corporea isolata continui a funzionare solo
in relazione al resto del corpo che è rappresentato in maniera non consapevole. Nell'immagine
cosciente del corpo non si ha una consapevolezza olistica, ma una consapevolezza delle parti che
possiamo controllare perché le percepiamo come nostre, mentre le altre parti del corpo, come gli
organi interni che non possono essere controllati, non riusciamo ad averne coscienza. Questa
consapevolezza del proprio corpo viene definita dall’autore come “immagine corporea”, un
concetto complesso che include in sé tre caratteristiche principali: “percettiva”, il corpo come viene
percepito dalla mia coscienza immediata; “cognitiva”, come costrutto concettuale e di
comprensione intellettuale del proprio corpo; “emotiva”, una caratteristica che concerne i sentimenti
e l’atteggiamento emotivo. Al contrario lo “schema corporeo” è una rappresentazione non
consapevole e operativa, è il modo in cui il corpo sperimenta il suo ambiente. Lo schema rispecchia
e determina la postura, la posizione generale che occupa il corpo nell’ambiente. La postura non
equivale alla posizione di un oggetto in uno spazio oggettivo ma è un’esperienza spaziale del corpo,
uno spazio vissuto che non è mai pienamente rappresentato nella coscienza. Questa non
consapevolezza riflette due aspetti: il primo, interessa la mancanza di consapevolezza della maggior
parte delle reazioni che avvengono ad un livello neurologico inferiore all'interno del nostro corpo
5
(Mason, 1961). Il secondo aspetto riguarda la non consapevolezza del nostro corpo applicata non
solo al nostro vissuto fisiologico ma anche al modo in cui il nostro corpo vive il suo ambiente.
L’affaticamento degli occhi ne è un esempio; la consapevolezza è diretta intenzionalmente
all’ambiente piuttosto che al nostro corpo. Quando durante una lettura gli occhi si stancano, “il
lettore non percepisce la sua fatica, ma rileva che la luce è troppo debole o che il libro è davvero
noioso o incomprensibile...I pazienti non percepiscono immediatamente che le funzioni corporee
sono disturbate, ma si lamentano circa il fatto che il lavoro non riesce, che l'ambiente è irritante e
faticoso” (Buytendijk, 1967/1974, p.62). Non siamo consapevoli della nostra postura e di come il
nostro sistema visivo scorre le pagine, avvertiamo la stanchezza attraverso la percezione di
cambiamenti nell’ambiente. Compaiono la fatica e il mal di testa e, gradualmente, l’assenza di
consapevolezza intenzionale dei nostri occhi cambia, perché ora la nostra attenzione è rivolta ad
essi ed entrano nel dominio della consapevolezza corporea. Il dolore diventa l’inquietudine
principale e causa del turbamento della lettura. Gallagher (1986) ha allontanato la confusione che
gravitava intorno al concetto di rappresentazione corporea, definendo in modo chiaro la distinzione
dicotomica tra schema e immagine corporea.
A sostegno di questa teoria, alcuni autori hanno cercato di fornire una base neuronale
attraverso lo studio delle doppie dissociazioni in pazienti con lesioni neurologiche. Paillard (1999)
ha associato la dicotomia schema-immagine corporea con due dicotomie anatomo-funzionali, quella
del cosa-dove di Ungerleider and Mishkin (1982) e quella di visione-per-percezione e visione-perazione di Milner e Goodale (1995). L’autore identifica l’immagine del corpo come oggetto della
coscienza e lo schema corporeo come oggetto non consapevole, anche se a volte quest’ultimo può
influenzare la percezione spaziale, la percezione degli oggetti e l’azione intenzionale. La tesi di
Paillard (1999) assume che una modalità sensorimotoria di elaborazione delle informazioni spaziali
coesiste con una modalità di rappresentazione, ed entrambe si organizzano e generano la propria
mappatura dello spazio (Paillard, 1987). La modalità sensorimotoria include principalmente quella
parte del mondo fisico con cui l'organismo è in relazione in virtù del suo apparato sensorimotorio di
base. Gli strumenti sensorimotori locali mantengono un rapporto diretto con il mondo fisico e
contribuiscono così al continuo aggiornamento della mappa del corpo in relazione con lo spazio
extra-corporeo in cui si trovano gli oggetti e a cui sono dirette le azioni. La modalità di
rappresentazione derivante dall’attività neurale, che esplora e consulta le rappresentazioni interne
dell'ambiente fisico, vengono immagazzinate nei sistemi di memoria. Essi comprendono
rappresentazioni mentali locali, relazioni spaziali di percorsi relativi a punti di riferimento, le
6
posizioni relative tra gli oggetti e la posizione del corpo stesso in relazione al suo ambiente. La
domanda che nasce spontanea è se le due modalità di elaborazione operino in parallelo, ciascuna
utilizzando i propri circuiti neurali e generando la propria mappatura dello spazio in due diversi
sistemi di riferimento, rispettivamente centrati sul corpo e sull’ambiente. Studi su pazienti con
lesioni neurologiche hanno tentato di rispondere a questo quesito.
Paillard (1999) applicò degli stimoli tattili su due pazienti con due differenti lesioni; il
primo, con infarto parietale sinistro, ebbe una anestesia sensoriale completa della mano destra;
l’esaminatore, dopo aver bendato il paziente, applicò uno stimolo tattile sulla mano non sensibile
del soggetto, il quale riuscì a toccare il punto esatto che era stato stimolato pur continuando a negare
qualsiasi sensazione corrispondente. Il secondo paziente, colpito da una neuropatia periferica con
assenza di propriocezione nei quattro arti, al contrario, riusciva ad identificare consapevolmente il
punto toccato dall’esaminatore ma non poteva indicarlo se non aiutato dalla vista. Questi risultati
hanno portato Paillard a concludere che nel primo paziente il deficit fosse correlato ad un danno a
livello dell’immagine corporea, quindi al sistema di percezione conscia e del cosa, mentre nel
secondo caso il deficit nella performance poteva essere ricondotto ad un danno inerente lo schema
corporeo, una disfunzione del sistema di azione inconscia e del dove.
Con How the Body Shapes the Mind, Shaun Gallagher (2005) fornisce un quadro generale
dell’importanza del corpo nei processi cognitivi. Gallagher riassume significativi risultati
sperimentali provenienti da un'ampia varietà di domini: neuropsicologia (deafferentazione, arto
fantasma e schizofrenia), neuroscienze (neuroni specchio), psicologia dello sviluppo (imitazione nel
neonato) e psicologia sociale (gesti comunicativi). Dall’insieme dei suoi studi Gallagher vuole
fornire una descrizione dettagliata della consapevolezza corporea e analizzare l’influenza del corpo
sull’autocoscienza, sulla percezione, sul linguaggio e sulla cognizione sociale. L’autore indica con
immagine corporea un sistema di percezioni, atteggiamenti e credenze relative al proprio corpo, in
contrasto con lo schema corporeo, un sistema di capacità sensorimotorie che operano senza
consapevolezza o necessitano di un monitoraggio percettivo (Gallagher, 2005). Egli giustifica
l’esistenza della dicotomia immagine/schema corporeo attraverso risultati ottenuti da studi di
doppia dissociazione in pazienti neurologici. Ha analizzato soggetti con neglect personale, il cui
deficit si esprime mediante un’incapacità ad orientare l’attenzione verso il lato controlaterale alla
lesione che si manifesta, di solito, più frequentemente a sinistra. Egli ipotizza che questo potrebbe
essere causato da un'interruzione della percezione dell’immagine corporea. Al contrario, in pazienti
con deafferentazione, che non ricevono informazioni tattili o propriocettive sotto il collo, il deficit si
7
manifesta con un’incapacità motoria se non osservano con attenzione ciò che stanno facendo. In
questo caso è il loro schema corporeo ad essere gravemente compromesso.
Queste dissociazioni possono essere riassunte affermando che lo schema corporeo è
generalmente dedicato all'azione inconscia e l'immagine del corpo è generalmente dedicato alla
percezione e all'azione cosciente (Berlucchi e Aglioti, 2010). La comprensione di come il cervello
rappresenta il proprio corpo è un argomento di studio complesso e di difficile concettualizzazione,
ma l’identificazione di aree della corteccia associate alla rappresentazione corporea, che
argomenterò nel secondo capitolo, hanno portato dei contributi empirici e teorici importanti.
1.3 Tassonomia triadica della rappresentazione corporea.
I dati riportati dalla letteratura su casi singoli e piccoli gruppi di studio hanno suggerito una
distinzione tra rappresentazioni del corpo che va oltre la tassonomia diadica, proponendo una tripla
rappresentazione. Il concetto di schema corporeo rimane invariato, conservando la caratteristica di
rappresentazione sensorimotoria del corpo che guida le azioni mediante informazioni efferenti ed
afferenti; il concetto originario di immagine corporea viene respinto e trasformato in due ulteriori
rappresentazioni: la descrizione strutturale del corpo (o mappa visuo-spaziale del corpo) e la
semantica del corpo. A livello visuo-spaziale si parla di una mappa topologica che origina
principalmente dall’ input visivo che definisce i confini, la vicinanza e la posizione delle parti del
corpo (Buxbaum e Coslett, 2001; Sirigu, Grafman, Bressler, e Sunderland, 1991). Questa
rappresentazione si basa principalmente sulla visione e sulla percezione somatica. A livello
semantico, l'immagine del corpo è in primo luogo concettuale e linguistico. Descrive lo scopo
funzionale delle parti del corpo, ne definisce il nome e il rapporto categorico, ad esempio, polsi e
caviglie sono entrambe articolazioni (De Vignemont, 2010). Diverse evidenze in letteratura
supportano la distinzione tra queste tre differenti rappresentazioni.
Molti ricercatori hanno riportato dati fisiologici che suggeriscono che l’efficacia di
un’azione probabilmente dipende dalla codifica della posizione tra le parti del corpo, come la
relativa posizione di ogni dito rispetto all'altro (Gallese, Fadiga, Fogassi, e Rizzolatti, 1996), nonché
le posizioni relative degli occhi, della testa e del tronco (Snyder, Grieve, Brotchie, e Andersen,
1998). Alcuni studi sul controllo motorio hanno suggerito che sia le informazioni sensoriali che
quelle di copia efferente (segnale motorio che informa il sistema sensoriale dell’azione che è in fase
di programmazione o di attuazione), possono essere integrati per consentire la correzione on-line di
errori motori così da generare una stima più accurata della posizione del corpo (Desmurget &
8
Grafton, 2000; Wolpert & Ghahramani, 2000;. Desmurget et al, 1999; Wolpert, Ghahramani, e
Jordan, 1995). Inoltre, Buxbaum, Giovannetti, e Libon (2000) hanno esaminato un paziente con
aprassia sostenendo che il suo deficit potesse dipendere da danni dello schema corporeo. Gli autori
suggeriscono che il deficit nel paziente aprassico con alterazione della pantomima, del
riconoscimento e dell’imitazione non dipende principalmente dall’integrità della rappresentazione
del gesto ma da un deficit della codifica dinamica delle posizioni intrinseche delle parti del corpo.
Anche Lackner (1988) ha dimostrato l’importanza tra queste relazioni, infatti, inducendo una
stimolazione vibratoria del muscolo del bicipite mantenendo il braccio fermo, si percepisce non
solo un prolungamento del braccio, ma anche la distorsione di altre parti del corpo con cui la mano
del braccio stimolato è in contatto. Questi studi suggeriscono la presenza di una rappresentazione
on-line delle relative posizioni tra le parti del corpo che implica quindi un richiamo dello schema
corporeo.
Per quanto riguarda la rappresentazione della struttura, ovvero, la mappa topologica del
corpo, diversi dati ne mostrano la validità. Pick (1922) ha descritto pazienti incapaci di indicare il
nome di parti del corpo su stessi e sugli altri (autotopoagnosia). E’ possibile ipotizzare che alla base
ci sia un danno della rappresentazione strutturale del corpo, che diversamente dallo schema
corporeo, riceve principalmente input visivi (Buxbaum & Coslett, 2001; Sirigu et al., 1991).
Buxbaum e Coslett (2001) hanno osservato un paziente autotopagnosico con danno diffuso
dell’emisfero sinistro che risultava deficitario rispetto al gruppo di controllo in un compito in cui
veniva chiesto di indicare verbalmente le parti del corpo o identificarle visivamente su se stessi o su
gli altri. Tuttavia questo paziente era in grado di eseguire perfettamente compiti in cui gli veniva
chiesto di indicare parti del corpo di animali e parti di oggetti inanimati. Questi risultati
suggeriscono un’incapacità di accedere in modo selettivo alla descrizione strutturale delle parti del
corpo umano, mantenendo una rappresentazione intatta dello schema corporeo dimostrata
dall’assenza di una differenza significativa rispetto ad un soggetto di controllo normale durante la
preparazione di un’azione motoria che permettesse di afferrare un oggetto.
L’ultima rappresentazione è quella della mappa semantica e lessicale del corpo deputata
all’elaborazione delle relazioni tra le parti, alle funzioni e ai nomi delle parti del corpo. Sirigu et al.
(1991) hanno riportato un caso di un paziente con autotopoagnosia e atrofia cerebrale diffusa che
aveva una “conservazione” selettiva dell'immagine del corpo. La prestazione del paziente non era
deficitaria quando doveva indicare le funzioni delle parti del corpo, ma risultava deficitario quando
doveva indicare verbalmente le relazione spaziali tra le parti del corpo (ad esempio, '' è il polso
9
accanto al braccio? ''). Anche Buxbaum e Coslett (2001) hanno osservata nel loro paziente
autotopagnosico con danno diffuso dell’emisfero sinistro, un esecuzione non deficitaria durante n
compito in cui doveva indicare l’associazione tra parti del proprio corpo e abbigliamento (ad
esempio con la presentazione di una foto che raffigurava una scarpa veniva chiesto di indicare la
parte del suo corpo con il quale era più strettamente associato), suggerendo che la sua conoscenza
semantica di parti del corpo era intatta. Inoltre Coslett et al. (2002) hanno osservato un paziente con
conservazione selettiva di informazioni semantiche di parti del corpo nonostante presentasse un
deficit nella comprensione delle parole da altre categorie (Coslett et al., 2002). Quando il paziente
doveva indicare e nominare le immagini raffiguranti parti del corpo e oggetti non corporei, egli
identificava correttamente tutte le parti del corpo e solo la metà degli stimoli di altre categorie.
Risultati differenti emergono da uno studio condotto da Suzuki, Yamadori, e Fujii (1997) che hanno
riportato un paziente con afasia di Broca ed infarto dell’emisfero sinistro con inclusione
dell’opercolo frontale. Il soggetto presentava danni a livello della rappresentazione semantica di
parti del corpo ma un’intatta comprensione semantica di parole inerenti categorie non corporee e
una conservata capacità di identificazione visiva di parti del proprio corpo.
I singoli casi descritti con deficit nelle diverse tre rappresentazioni corporee non mostrano prove
concrete in merito alla validità psicologica e alle basi anatomiche. Schwoebel e Coslett (2005)
hanno effettuato uno studio atto ad indagare la prevalenza e i substrati anatomici di queste tre
rappresentazioni del corpo. Gli autori hanno esaminato la performance di 70 pazienti con ictus
unilaterale e 18 controlli sani di pari età in una serie di compiti sviluppati per valutare lo schema
corporeo, la descrizione strutturale del corpo e l'immagine del corpo semantico/lessicale. I compiti
relativi alla valutazione dello schema corporeo sono:
“Compito di immaginazione/azione della mano”: quattro movimenti con difficoltà variabile,
che va dal tocco ripetitivo di pollice e indice, all’estensione indipendente dell’indice e del
mignolo. I soggetti sono stati testati in entrambe le condizioni sia con la mano ipsilesionale
che con quella controlesionale. Quando l’esaminatore dava verbalmente il via i pazienti
avevano il compito di immaginare di eseguire il movimento specifico della prima
condizione per 5 volte, in modo rapido e accurato e dichiarare verbalmente la fine
dell’attività mentale; nella seconda condizione, è stato chiesto ai pazienti di eseguire lo
stesso movimento per 5 volte. Ogni movimento immaginato ed eseguito è stato testato due
volte e sono stati registrati i tempi.
10
“Compito di lateralità della mano”: compito spesso utilizzato in letteratura (Parsons, Fox et
al., 1995;. Parsons, 1987, 1994; Tomasino, Rumiati, e Umiltà, 2003; Johnson, 2000; Coslett
1998). Sono state presentate ai soggetti delle immagini con stimoli di mani con la
prospettiva del palmo o del dorso in quattro orientamenti diversi: le dita che puntano verso
l’esterno, dita rivolte verso il soggetto, le dita che puntano a sinistra e le dita che puntano a
destra. I soggetti avevano il compito di dare un giudizio di lateralità muovendo la mano
destra o sinistra. I pazienti emiplegici rispondevano indicando alla mano controlesionale.
Per ogni soggetto è stata calcolata l’accuratezza complessiva per la mano ipsilesionale e
controlesionale.
Compiti per la valutazione della descrizione strutturale del corpo:
“Localizzazione di parti del corpo isolate”: sono state presentate 24 immagini di parti del
corpo ed è stato chiesto ai pazienti di indicare la stessa parte sul proprio corpo.
“Localizzazione di stimoli tattili”: l’esaminatore dopo aver applicato degli stimoli tattili con
un pennello sul corpo del soggetto che si trovava seduto con gli occhi chiusi, ha chiesto di
localizzare su un manichino il punto esatto in cui aveva percepito lo stimolo sul proprio
corpo.
“Abbinamento di parti del corpo mediante localizzazione”: una parte del corpo target è stata
presentata visivamente ed è stato chiesto ai pazienti di indicare 1 delle 3 parti del corpo
raffigurate che era più vicino sulla superficie del corpo alla parte del corpo target.
Compiti di valutazione dell'immagine corporea semantico/lessicale:
“Abbinamento di parti del corpo rispetto alla funzione”: sono stati utilizzati gli stimoli del
precedente compito di localizzazione. I pazienti avevano il compito di selezionare 1 delle 3
parti del corpo della foto più strettamente correlata in termini di funzione alla parte del
corpo target. Le immagini comprendevano anche una parte sulla superficie del corpo in
prossimità alla parte del corpo raffigurato e una parte del corpo estranea. Ad esempio, se
l'obiettivo era un gomito le opzioni di risposta includevano un ginocchio (la scelta giusta),
un avambraccio, e il naso.
“Abbinamento di parte del corpo con abbigliamento e oggetti”: un capo di abbigliamento è
stato presentato con 4 parti del corpo. Ai soggetti è stato chiesto di indicare l'immagine della
parte del corpo con la quale il capo di abbigliamento o un accessorio era più strettamente
associato. Le foto includevano anche immagini concettualmente correlate, non collegate e
parti del corpo contigue.
11
In linea con i dati dei singoli casi e piccoli gruppi in letteratura, i risultati dello studio di questo
gruppo molto ampio hanno confermato con forza l’indipendenza delle tre rappresentazioni del
corpo. Lo schema corporeo, la descrizione strutturale del corpo e l'immagine del corpo sono
rappresentazioni dissociabili. Sette soggetti hanno mostrato un deficit sul compito di
immaginazione/azione della mano con esecuzione nella norma su tutti gli altri compiti di
rappresentazione del corpo. Allo stesso modo, 6 soggetti hanno eseguito in modo anomalo il solo
compito di giudizio di lateralità della mano e due soggetti in modo deficitario solo il compito di
descrizione strutturale del corpo. Infine, tre soggetti hanno avuto una performance carente nel
compito di immagine corporea. Inoltre mediante studi di imaging Schwoebel e Coslett (2005)
hanno indagato le basi neurali delle rappresentazioni del corpo. I risultati suggeriscono che le
rappresentazioni della descrizione della struttura del corpo e l’immagine del corpo sono lateralizzate
nell'emisfero sinistro. In particolar modo il danno al lobo temporale sinistro è più costantemente
associato con prestazioni ridotte su tali compiti (Schwoebel e Coslett, 2005). Nonostante queste
evidenze, i risultati riportati inerenti al deficit di performance associato a lesioni del lobo temporale
non consente la discriminazione della differenza tra le regioni temporali associate alla
rappresentazioni strutturale del corpo e all’immagine corporea. L’ analisi dello schema corporeo ha
evidenziato una doppia dissociazione tra le prestazioni in compiti di immaginazione/azione di una
mano e compiti di lateralità della mano associato ad un deficit bilaterale in entrambi i casi. In
letteratura ci sono dati discordanti sull'attivazione bilaterale o unilaterale rispetto a questi compiti.
La spiegazione potrebbe dipendere dalle differenti richieste del compito o dalla localizzazione della
lesione (Schwoebel e Coslett, 2005). Tuttavia, nonostante i dubbi sulla lateralizzazione emisferica, i
risultati dei due compiti mostrano un’attivazione della Corteccia Frontale dorso-laterale e/o lobo
parietale ma la base anatomica per la dissociazione non è stata chiarita.
Complessivamente questi risultati mostrano una tripla dissociazione tra schema corporeo,
immagine strutturale del corpo e immagine del corpo semantico/lessicale, suggerendo che le
rappresentazioni sono funzionalmente dissociabili. Anche l’associazione delle basi anatomiche
correlano con i processi e le regioni cerebrali che mediano il riconoscimento e la rappresentazione
delle parti del corpo nello spazio (Goodale e Milner, 1992; Mishkin, Ungerleider, e Macko, 1983).
Alla luce di questi dati non sorprende che la descrizione strutturale e l'immagine del corpo siano
compromesse da lesioni temporali e che le alterazioni dello schema corporeo siano associate con
lesioni che coinvolgono la Corteccia Frontale dorso-laterale e / o lobi parietali.
12
1.4 Plasticità cerebrale e rappresentazione corporea.
Ognuno di noi possiede rappresentazioni multiple del proprio corpo a livello cerebrale. La
neuropsicologia suggerisce che la simulazione mentale di qualsiasi movimento è guidato dal nostro
schema corporeo (de Lange et al. 2005; Parsons 1994; Schwoebel et al. 2001), inteso come una
mappa topologica del nostro corpo in generale (Corradi-Dell’Acqua et al. 2009; Schwoebel and
Coslett 2005; Buxbaum and Coslett 2001; Sirigu et al. 1991; Pick 1922). Quando una parte
anatomica subisce una modificazione, come nel caso dell’amputazione di un arto, la
rappresentazione mentale del nostro corpo deve essere sottoposto ad un “rimappaggio”.
Ramachandran e Rogers-Ramachandran (1996) individuano la presenza di tre fattori che
giocano un ruolo fondamentale in pazienti con amputazione che avvertono la presenza di un arto
fantasma:
a) Se a causa di una lesione del nervo periferico, prima dell’amputazione, l’arto è
paralizzato, dopo l’asportazione chirurgica il paziente percepisce la presenza
dell’arto fantasma con la stessa sensazione di paralisi precedente. Questo fenomeno è
stato nominato “paralisi appresa” (Ramachandran, 1994);
b) I pazienti, dopo aver subito l’amputazione, riferiscono di poter generare
volontariamente dei movimenti con l’arto fantasma, ma con il passare del tempo
questa capacità svanisce in quasi tutti i soggetti.
c) Come succede nella maggior parte dei casi, gli amputati provano sensazioni dolorose
a livello del moncone e hanno difficoltà nell’avviare il movimento poiché un tale
tentativo potrebbe amplificare il dolore.
A volte i pazienti percepiscono movimenti involontari dell’arto fantasma, probabilmente
perché la parte frontale del cervello continua ad inviare messaggi ai muscoli dell’arto nonostante
quest’ultimo non ci sia più. Questo avviene perché il cervello non è a conoscenza del fatto che la
mano ad esempio, è stata amputata, probabilmente questi comandi di movimento sono
contemporaneamente monitorati dai lobi parietali che si occupano dell’immagine corporea
(Ramachandran, 1996).
L’autore crea uno strumento, la “virtual reality box”, al fine di aiutare i pazienti a risolvere il
problema della paralisi dell’arto fantasma che induce sensazioni dolorifiche. La virtual reality box è
composta da una scatola nella cui superficie interna è posizionato uno specchio, mediante il quale, il
paziente, attraverso l’immagine riflessa dell’arto sano, riesce a ricreare quello mancante su cui è
13
localizzato il dolore. Il soggetto dopo aver inserito entrambi gli arti (arto sano e moncone)
all’interno della scatola ha il compito di svolgere alcuni esercizi. Vedendo l’immagine dell’arto
sano riflessa allo specchio, il paziente è in grado di compiere il movimento oltre che con l’arto sano,
anche con quello amputato. In assenza di feedback tattili e propriocettivi, viene attivata una
strategia diversa a livello cerebrale, con cui, attraverso la vista e l’osservazione del movimento
riflesso che attiva i neuroni specchio, il paziente riceve un feedback per il controllo del movimento.
Avviene una riprogrammazione a livello corticale dello schema corporeo, il soggetto perviene ad
un’illusione propriocettiva dell’arto amputato e questo controllo permette la diminuzione del
dolore.
La capacità di riorganizzazione e riprogrammazione a livello della corteccia è un processo che
affascina tuttora la neuropsicologia. Molti studi hanno dimostrato che questa plasticità può, nella
maggior parte dei casi, portare ad un adattamento in risposta ad una lesione cerebrale. Il cervello
riesce ad attuare delle modifiche in brevissimo tempo e può anche indurci ad allucinare una diversa
percezione del nostro corpo. La rappresentazione cerebrale del corpo origina dall’integrazione delle
informazioni sensoriali, quali vista, tatto e propriocezione. Mettendo in conflitto queste diverse
modalità sensoriali è stato dimostrato come esse possano influenzare la rappresentazione mentale
del nostro corpo.
Lo studio condotto da Lackner (1988) ha evidenziato come un conflitto tra tatto e
propriocezione distorca la rappresentazione della superficie corporea. Questa distorsione, creata
sperimentalmente, è stata denominata “illusione di pinocchio” (Lackner, 1988). I fusi muscolari,
sono dei recettori propriocettivi localizzati nei muscoli che forniscono informazioni sulla variazione
di lunghezza di questi ultimi. Oltre che fisiologicamente, i fusi possono essere attivati
sperimentalmente mediante la stimolazione del tendine con uno stimolo vibratorio. Se la
stimolazione viene applicata sul bicipite, nonostante questo rimanga fermo, si ha la sensazione che
il braccio si sia steso. Se la stimolazione viene effettuata mentre il soggetto tocca la punta del suo
naso con le dita dello stesso braccio si origina una condizione paradossale, ossia, la percezione che
il braccio si allunghi, la mano si allontani dal viso e il naso si allunghi fino a 30 cm (Medina, 2010).
Ciò dimostra che l’informazione tattile si integra con quella vestibolare, andando ad
offuscare l’informazione visiva, da cui origina l’illusione di pinocchio (Lackner, 1988).
L’esperimento spiega il conflitto sensomotorio: visto che il corpo e la testa sono stazionari,
l’illusione colpisce solo il naso in quanto è a contatto con il braccio stimolato. Le parti del corpo
vengono rappresentate nel loro rapporto reciproco e questa rappresentazione è il risultato delle
14
numerose informazioni sensoriali che provengono dal corpo e sono integrate in un tutto funzionale
(De Vignemont, 2005).
Botvinick e Cohen nel 1998 hanno proposto uno studio diverso per indagare se una mano
artificiale può entrar a far parte del nostro schema corporeo innescando un conflitto multisensoriale
tra sistema propriocettivo e sistema visivo. Sono stati testati 10 soggetti e ognuno di loro doveva
sedersi davanti ad una scrivania sulla quale, in corrispondenza del braccio sinistro, si trovava una
mano di gomma di dimensioni reali e molto simile a quella vera, mentre la mano vera veniva
nascosta dietro ad un pannello. Gli sperimentatori con due pennelli stimolavano in maniera sincrona
sia la mano reale che quella di gomma mentre il soggetto doveva tenere lo sguardo fisso su
quest’ultima. Dieci minuti dopo la stimolazione fu chiesto ai partecipanti di compilare un
questionario volto ad indagare l’esperienza diretta dei soggetti. Il risultato evidenziò che durante la
stimolazione i soggetti avevano percepito la sensazione tattile del pennello effettuata sulla mano
artificiale. Gli autori hanno ipotizzato che questa illusione potesse dipendere dal tentativo del
cervello di risolvere il conflitto multisensoriale attraverso l’incorporazione della mano di gomma
nel proprio schema corporeo. Botvinick e Cohen (1988) hanno riproposto l’esperimento precedente
dividendo i soggetti in due gruppi (sperimentale e controllo) e somministrando il compito
chiedendo ai soggetti di chiudere gli occhi. Nei soggetti del gruppo sperimentale la stimolazione
venne effettuata in modo sincrono mentre nel gruppo di controllo in modo asincrono. Dopo aver
indotto l’illusione i soggetti del gruppo sperimentale, nell’ effettuare una ricerca intermanuale per
indicare la posizione della loro vera mano, commettevano un errore di posizionamento verso la
mano finta. Questo risultato suggerisce che il sistema visivo ha portato ad una ricalibrazione del
braccio (Botvinick e Cohen, 1988).
La risposta fisiologica a questo tipo di illusione è stata una scoperta importante in ambito dei
disturbi che riguardano la consapevolezza corporea, come ad esempio nel caso della sensazione
dolorifica dell’arto fantasma percepita dai pazienti con amputazione di un arto. Una riabilitazione
mediante queste procedure infatti porta ad una riorganizzazione cerebrale, facilitando il
riadattamento del paziente.
15
CAPITOLO 2
Elaborazione sensorimotoria in pazienti con deafferentazione
periferica.
2.1 Aree cerebrali selettive per il corpo.
Ogni individuo attraverso la visione e l’interpretazione delle azioni altrui acquisisce, anche
per imitazione, nuove competenze; questo è un processo che interessa tutto l’arco della vita
(Meltzoff e Moore, 1977). Con l’esperienza gli esseri umani sono capaci di distinguere un’azione di
natura “biologica” da una compiuta da meccanismi artificiali, anche se gli stimoli disponibili sono
limitati (Johansson, 1973). Questa abilità nell’essere umano è dovuta a meccanismi cerebrali
specializzati nell’elaborazione del corpo e dell’azione. Studi di risonanza magnetica funzionale
hanno evidenziato che i volti e i corpi vengono elaborati in regioni corticali specializzate
denominate: occipital face area (OFA), fusiform face area (FFA), extrastriate body area (EBA) e
fusiforme body area (FBA).
Nello specifico, per quanto riguarda EBA ed FBA, Downing et al. (2001) utilizzando un
paradigma di risonanza magnetica funzionale (fMRI) hanno dimostrato che i soggetti, alla visione
di immagini raffiguranti il corpo umano o parti di esso, producevano un’attivazione a livello della
corteccia occipito-temporale molto più forte rispetto a quando erano di fronte ad immagini di
oggetti, o parti di oggetti, inanimati. Quest’area, denominata EBA, è stata trovata specifica per la
codifica del corpo e parti del corpo, senza volto. Infatti, risulta una diminuzione dell’attivazione di
EBA con stimoli che rappresentano un viso rispetto a quanto si mostra solo il corpo (Morris et al.
2006). E’ stata individuata un’ulteriore area selettiva per i corpi senza la codifica del volto, questa
zona è stata denominata FBA, situata nel giro fusiforme medio (Taylor et al., 2007). La differenza
tra queste due aree risiede nella diversa analisi che effettuano sul corpo. Infatti, sembrerebbe che
EBA risponda alla codifica di piccole parti del corpo, come le singole dita di una mano (Taylor et
al., 2007), mentre FBA elabora il corpo nella sua globalità, codificando le parti più grandi, come il
busto e gli arti (Taylor et al., 2007). Le prime dimostrazioni sulla specializzazione di queste aree
sono state fornite da Urgesi et al. (2004) attraverso l’utilizzo della Stimolazione Magnetica
16
Transcranica ripetitiva. Interrompendo transitoriamente la funzionalità di EBA si è osservata una
significativa diminuzione dei tempi di risposta durante lo svolgimento di un compito visivo
strutturato con stimoli raffiguranti parti del corpo. Il dato è stato rinforzato dall’assenza di effetti
significativi sui tempi di risposta utilizzando lo stesso compito visivo ma con stimoli facciali e non
corporei. “La rTMS è stata applicata tra i 150-250 ms dopo la presentazione del trial, suggerendo
un coinvolgimento precoce di EBA nell’elaborazione delle informazioni visive” (Berlucci e Aglioti,
2009). Inoltre altri studi hanno rilevato che l’attivazione di EBA è maggiore per le rappresentazioni
allocentriche del corpo rispetto a quelle egocentriche (Chan et al 2004; Saxe et al 2006).
Uno studio di fMRI di Chan e colleghi (2004) sembra indicare che l’attività di questa area sia
influenzata in modo specifico dalla prospettiva, egocentrica o allocentrica, in cui sono presentate le
immagini visive del corpo, senza però rispecchiare una differenza nella discriminazione delle parti
corporee come appartenenti a sé stessi o a un altro (Chan, Peelen e Downing, 2004). Questo è stato
confermato anche da studi funzionali che hanno riportato un’attivazione di EBA rispetto alla
codifica di parti del corpo da diversi punti di vista: EBA di destra, anche se di poco, si attiva
maggiormente in risposta ad una visione allocentrica più che egocentrica del corpo (Chan et al
2004) e delle sue parti (Saxe et al. 2005) rispetto a EBA di sinistra. Saxe et al. (2005) mediante uno
studio fMRI hanno indagato l’attivazione di EBA in un gruppo di 10 soggetti sani sottoponendoli ad
un compito di riconoscimento dello stimolo. L’esperimento comprendeva quattro condizioni di
riconoscimento: (I) foto in scala di grigi di corpi interi escluso la testa, (II) parti del corpo, (III)
oggetti interi e (IV) parti di oggetti. Gli stimoli per il riconoscimento della prospettiva del corpo
comprendevano 20 fotografie raffiguranti parti del corpo da una prospettiva allocentrica e 20 da una
prospettiva egocentrica, i soggetti avevano il compito di premere un pulsante se lo stesso stimolo
appariva due volte in successione immediata. Le immagini sono state presentate su uno schermo per
16 secondi in blocchi di 20 immagini per la stessa condizione. I risultati dell’analisi fMRI hanno
mostrato un’attivazione maggiore di EBA per le immagini dei corpi umani. In particolare,
l’attivazione di EBA di destra era maggiore durante il riconoscimento di parti del corpo elaborati da
una prospettiva allocentrica (Saxe et al. 2005). Questi risultati confermano che EBA di destra ed
EBA di sinistra sono coinvolte rispettivamente nel riconoscimento del corpo da una prospettiva
allocentrica ed egocentrica (Turk et al, 2002;.. Brady et al, 2004). La rappresentazione mentale del
nostro corpo, quindi, deriva da un’integrazione multisensoriale che coinvolge non solo le
informazioni sensorimotorie ma anche il nostro sistema visivo (Corradi-Dell’Acqua, 2008).
17
Uno studio condotto da Van Elk et al. (2013) ha verificato come la diversa elaborazione che
viene effettuata dal nostro cervello per gli arti superiori e inferiori, modula l’integrazione
multisensoriale dello spazio peripersonale attraverso la congruenza e la postura dell’arto. Sono stati
utilizzati stimoli cross-modali in cui i partecipanti avevano il compito di discriminare gli stimoli
tattili applicati sulle proprie mani o piedi reali cercando di ignorare la distrazione derivante dalla
visione di stimoli applicati su arti di gomma. Ponendo la parte del corpo di gomma in una posizione
anatomicamnete congruente a quella reale è stato osservato un effetto di congruenza cross-modale
(CCM per stimoli visuo-tattili applicati alle mani e ai piedi. Questo mostra che in una condizione di
arti non incrociati la rappresentazione peripersonale di mani e piedi sono simili e questo effetto è
rafforzato quando le parti del corpo in gomma sono collocate in una posizione anatomicamente
congruente. Tuttavia si verifica una situazione diversa quando dobbiamo elaborare separatamente
gli stimoli di mani e piedi ponendo le parti del corpo reali e di gomma in una posizione incrociata.
Per le mani l’effetto cross-modale era regolato dalla posizione incrociata, mentre per i piedi non
c’era un effetto cross-modale dipendente dalla posizione. Questo riflette il fatto che l’informazione
visiva delle mani è più facile da elaborare in modo propriocettivo rispetto ai piedi, dovuto
probabilmente a differenti rappresentazioni funzionali di queste parti. Infatti non si evidenziano
effetti cross-modali per i piedi né se i piedi reali né se quelli di gomma erano posizionati in una
posizione congruente o meno. Probabilmente l’effetto è legato all’esperienza visiva, infatti abbiamo
più esperienza delle nostre mani che dei nostri piedi, di conseguenza le informazioni visive della
mano di gomma saranno integrate più facilmente a livello percettivo e un’incongruenza porta ad un
ridotto CCM (Pavani et al, 2000;. Zopf, Savage, e Williams, 2010). L’assenza di CCM per i piedi
reali e di gomma nelle diverse posizioni indica che l’elaborazione degli arti superiori e inferiori
avviene mediante meccanismi diversi (Van Elk et al., 2013). Queste differenze sono state
evidenziate anche in studi di rotazione mentale che hanno riportato una maggior facilità nel compito
immaginativo per le mani rispetto ai piedi (Gentilucci, Daprati, e Gangitano, 1998; Ionta, Fourkas,
Fiorio, e Aglioti, 2007; Parsons, 1987). Un altro fattore rilevante potrebbe essere che i piedi sono
più distali rispetto alle mani e le informazioni visive potrebbero essere ridotte. Questo studio
dimostra che l’esperienza delle nostre azioni ha una forte influenza sulla rappresentazione visuomotoria e propriocettiva del nostro corpo (Calvo-Merino, Grezes, Glaser, Passingham, e Haggard,
2006; Gazzola et al., 2007). “Di conseguenza ci si aspetterebbe che lo spazio peripersonale
verrebbe rimappato più facilmente per le parti del corpo con cui abbiamo un’esperienza più
profonda” (Van Elk et al., 2013).
18
2.2 Meccanismi Top-Down e Bottom-up della rappresentazione
corporea.
Le informazioni top-down e bottom-up interagiscono tra di loro dando forma alla rappresentazione
del proprio corpo. In letteratura sono state descritte diverse tassonomie per spiegare il modo in cui
viene ripartita la consapevolezza corporea a livello cognitivo. E’ stata proposta una tassonomia
diadica che distingue tra “schema corporeo (una rappresentazione sensomotoria basata su
informazioni afferenti ed efferenti) e immagine del corpo (una descrizione astratta del corpo basata
principalmente su l’esterocezione visiva)” (Dijkerman & de Haan, 2007; Gallagher 2005; Paillard,
1999 Rossetti, Rode, e Boisson, 1995). Altri autori hanno proposto una tassonomia triadica
(Schwoebel & Coslett 2005; Sirigu, Grafman, Bressler, e Sunderland, 1991) che suddivide il
concetto di immagine del corpo in una mappa visuo-spaziale del corpo (la postura e la posizione del
corpo in relazione all’ambiente) e una rappresentazione semantica del corpo (la rappresentazione
linguistica e concettuale del corpo), mentre il concetto di schema corporeo rimane invariato.
Uno studio condotto da Palermo et al. (2014) ha indagato l’influenza della perdita di
informazioni top-down e bottom-up sulla mappa visuo-spaziale del corpo somministrando il sub
test di rievocazione fronte-corpo (Daurat-Hmeljiak et al., 1978) sia ai pazienti con danno periferico
(amputati di arto inferiore) che ai pazienti con danno centrale (danno cerebrale unilaterale). I
risultati hanno riportato una prestazione peggiore degli amputati e dei cerebrolesi rispetto ai
controlli. In particolare è emersa una maggior compromissione durante la localizzazione della
gamba (destra e sinistra) rispetto alle altre parti del corpo. Quindi la rappresentazione del corpo può
essere alterata in entrambi i gruppi. Ciò dimostra, secondo gli autori che tale rappresentazione non è
innata e che non è strettamente legata alla percezione di altri corpi. Studi di neuroimaging hanno
evidenziato che le relazioni visuo-spaziali del corpo sono regolate da un’attivazione bilaterale delle
aree del cervello, principalmente nelle regioni parietali e temporali. Per questo motivo Palermo et
al. (2014) hanno ipotizzato una compromissione della rappresentazione sia in soggetti con danno
dell’emisfero destro che quelli con danno dell’emisfero sinistro. Infatti anche l’analisi effettuata sui
casi singoli (gruppo con due pazienti con amputazione dell’arto sinistro e gruppo con un paziente
con danno cerebrale sinistro) hanno mostrato una prestazione peggiore nel compito FBE rispetto ai
controlli. Complessivamente i risultati dello studio hanno mostrato che la prestazione di entrambi i
gruppi di pazienti con danno destro era compromessa durante il compito con difficoltà nella
codifica della localizzazione delle le parti del corpo. Quindi la perdita di informazioni dell’arto
amputato inferiore sia destro che sinistro incide significativamente sulla capacità di rappresentare la
19
posizione e le relazioni tra le parti del corpo così come avviene anche nei pazienti cerebrolesi destri
e sinistri che non ricevono informazioni top-down. Tuttavia gli studi effettuati sullo schema
corporeo mediante compiti di rotazione mentale (Parsons, 1987) in soggetti sani hanno mostrato un
effetto di dominanza in cui il piede destro è riconosciuto più velocemente rispetto al piede sinistro.
Anche i risultati ottenuti da Nico (2004) hanno rafforzato i dati di Parsons (1987); i pazienti con
amputazione di arto superiore dominante eseguendo un compito di giudizi di lateralità si sono
mostrati più lenti e meno accurati rispetto a quelli di arto non dominante. Su queste evidenze è
possibile ipotizzare che la mappa visuo-spaziale in pazienti con danno destro è più compromessa
rispetto ai pazienti con danno sinistro (Palermo et al., 2014). E’ stata inoltre evidenziata una
compromissione del compito che si estendeva anche alle altre parti del corpo oltre che all’arto
colpito e sono state fatte due ipotesi (Palermo et al., 2014):
a) si è verificato un effetto di ampliamento della rappresentazione di altre parti del corpo
b) il deficit era dovuto alla forte necessità di confrontare la posizione di una parte del corpo
in relazione alle altre.
I risultati sui pazienti cerebrolesi si discostano da altri studi, poiché Palermo et al. (2014)
hanno riscontrato un deficit della rappresentazione visuo-spaziale in pazienti con danno unilaterale
destro ma anche nel caso singolo con danno cerebrale sinistro, mentre studi precedenti hanno
riscontrato questo disturbo solo in pazienti con lesioni dell’emisfero sinistro (Schwoebel & Coslett,
2005). Gli autori in base a questi dati suggeriscono che la mappa visuo-spaziale è elaborata da
entrambi gli emisferi e la diversa attivazione dipende dal tipo di strategia (verbale o non) utilizzata
dai soggetti durante l’esecuzione del compito. In base ai dati ottenuti dalla letteratura si è a
conoscenza della specializzazione dell’emisfero destro per l’analisi di parti globali e dell’emisfero
sinistro per la codifica di caratteristiche locali (Sergent, 1982). Di conseguenza Palermo et al.
(2014) hanno suggerito un’implicazione dell’emisfero destro nella codifica topologica dell’intero
corpo e le relazioni spaziali di tutte le parti del corpo, mentre l’emisfero sinistro è deputato
all’elaborazione delle caratteristiche topologiche di singole parti del corpo. Nello studio inoltre è
stata riscontrata una correlazione tra FBE e il punteggio al test per la valutazione del neglect visuospaziale. La performance era caratterizzata dall’utilizzo di una prospettiva egocentrica sottolineando
che la costruzione della mappa visuo-spaziale non origina dall’esperienza visiva di corpi di altri
soggetti ma dipende dalle esperienze che sperimentiamo direttamente con il nostro corpo. Lo studio
ha messo in evidenza l’importanza di una mappa visuo-spaziale per la consapevolezza del nostro
20
corpo e ha inoltre sottolineato il contributo di entrambi gli emisferi affinché la nostra mappa
topologica sia efficiente (Palermo et al., 2014).
2.3 Simulazione mentale di azioni motorie in pazienti amputati.
La letteratura riporta molte dimostrazioni della somiglianza tra attività cerebrale che simula un
movimento mentalmente e attività cerebrale che compie realmente un’azione. Questa analogia si
riscontra ad esempio in soggetti con compromissione delle abilità motorie. Infatti la disabilità
motoria si riflette anche nell’immaginazione motoria del movimento, ad esempio in pazienti con
emiparesi, in seguito a lesione unilaterale della corteccia motoria, si riscontra una lentezza anche
durante la simulazione mentale del movimento che coinvolge l’arto emiplegico (Sirigu et al., 1995).
La somiglianza è dimostrata anche in compiti che richiamano implicitamente l’esecuzione mentale
di un movimento, come nel caso della valutazione della lateralità di una mano. Parsons (1987) ha
somministrato un compito di giudizio di lateralità di una mano o di un piede ad un gruppo di
soggetti. I partecipanti compivano una trasformazione mentale prima di dare una risposta legata al
movimento reale che doveva essere compiuto dalla mano. Di conseguenza è emerso che i tempi di
risposta aumentavano in relazione all’angolo di rotazione dello stimolo presentato (Pearson, 1987)
sottolineando l’analogia tra queste due attività cerebrali di immaginazione e azione.
Nico et al. (2004) hanno condotto uno studio su pazienti con amputazione dell’arto superiore
per verificare se e in che modo, la mancanza di una parte fisica del proprio corpo potesse
influenzare un compito di immaginazione mentale di un’azione rispetto all’arto amputato e se
l’utilizzo di una protesi potesse migliorare o peggiorare l’esecuzione del compito. Sono state fatte
diverse ipotesi circa l’esecuzione del compito immaginativo. La prima sostiene che se
precedentemente era presente la rappresentazione completa del corpo, l’azione mentale dell’arto
mancante doveva essere implicita e in alterata, permettendo ugualmente la simulazione. La seconda,
come dimostrato in letteratura da Dominey et al (1995) e Sirigu et al (1996) su soggetti
parkinsoniani ed emiplegici, la simulazione del movimento dipende dallo stato attuale del corpo. Di
conseguenza una deafferentazione periferica dovrebbe comportare una compromissione della
simulazione mentale rispetto all’arto amputato. Il compito proposto da Nico et al. (2004) richiedeva
ai soggetti di dare, velocemente e in modo accurato, un giudizio verbale sulla lateralità di una mano
presentata sullo schermo di un computer. Il set di stimoli era costituito da figure di mani destra e
sinistra, visti da 4 diversi punti di vista (due posizioni frontali: dorso e palmo, e due laterali: lato del
pollice e lato del mignolo laterali. Inoltre gli stimoli avevano orientamenti diversi: sei orientamenti
corrispondevano a posture facilmente raggiungibili (mano destra, da 30 ° a 240 ° in senso
21
antiorario, mano sinistra, da 330 ° a 120 ° in senso orario). Le altre restanti sei posizioni invece,
richiedevano movimenti innaturali e difficili da raggiungere (mano destra: da 60 ° a 210 ° in senso
orario; a sinistra: da 300 ° a 150 ° in senso antiorario). I risultati riportano che i pazienti con
amputazione dell’arto superiore nonostante trovassero maggior difficoltà nel compito rispetto ai
soggetti sani (dati i maggior tempi di risposta), riescono a dare giudizi di manualità. Questo
dimostra che l’attivazione del sistema motorio avviene anche senza la controparte fisica, infatti, i
pazienti che avevano perso la sensazione di arto fantasma già da tempo, hanno riportato che durante
l’esecuzione del compito questa sensazione è riemersa. Questo è in linea con l’ipotesi di
Ramachandran (1996) che afferma che un falso feed-back visivo può indurre l’illusione di
movimento dell’arto fantasma. L’aumento dei tempi di reazione può essere spiegato dalla diversa
strategia utilizzata dai pazienti dato che le informazioni dalla periferia non sono più disponibili
(Nico et al., 2004). Per l’esecuzione del compito evidentemente hanno utilizzato una strategia
visuo-spaziale e non cinestetica-motoria scegliendo di elaborare la rotazione della mano trattandola
come se fosse un oggetto. Gli orientamenti più facili invece è possibile che siano stati trattati con
una strategia di corrispondenza visiva e i tempi di risposta più lunghi derivano probabilmente dalle
modificazioni sulla rappresentazione mentale del corpo avvenuta nei pazienti amputati. “Infatti, la
perdita di un arto può aumentare la difficoltà di riconoscere la mano mancante perché la
familiarità visiva dell'arto corrispondente non è più disponibile” (Nico et al., 2004). Nel loro
insieme questi risultati dimostrano un’attivazione inalterata del sistema motorio. Un altro dato
significativo si riscontra in pazienti amputati di arto dominante il cui compito è risultato più difficile
rispetto al gruppo di amputati di arto sinistro. Ciò può dipendere da una fase preliminare in cui i
soggetti selezionano in modo automatico la rappresentazione della loro mano dominante e non
avendo un riscontro della controparte fisica, attivano o una strategia di immagini visive o traslano
l’azione motoria dell’arto presente. Questo porterebbe a tempi di risposta più lunghi e ad errori di
giudizio di lateralità. Infine è stato riportato un effetto negativo per i soggetti provvisti di protesi
meno che per due soggetti dotati di protesi mioelettrica che permetteva il movimento del polso e un
pollice opponibile. I risultati fanno ipotizzare che soggetti con protesi estetica avevano solo un feedback visivo che evidenziava l’inefficacia delle innervazioni motorie per l’arto con conseguente
decadimento nelle prove di simulazione motoria. Mentre i pazienti con protesi che permettono un
movimento artificiale della mano, probabilmente integravano le informazioni nello schema
corporeo riaggiornando il sistema motorio è migliorando le prestazioni di simulazione mentale.
22
Curtze et al. (2010) hanno condotto uno studio simile su pazienti amputati di arto inferiore e
su un soggetto con giroplastica di una gamba. L’autore ha voluto valutare gli effetti di un arto
amputato e di un arto trattato con giroplastica per verificare se sussistono differenti effetti di
rappresentazione del corpo a livello della corteccia di questi pazienti. E’ stato utilizzato un set di
stimoli con immagini riprese dai disegni originali di Parsons (1987), di piedi naturali o protesici
destri e sinistri organizzati in diversi orientamenti. Questi ultimi erano rappresentati in sei
orientamenti sia da una prospettiva ventrale che dorsale, per un totale di 72 diversi stimoli. Il
compito del soggetto era quello di valutare la lateralità dello stimolo premendo un apposito pulsante
con i due indici in modo rapido ma accurato ed evitando la vista dei piedi i quali dovevano essere
posti sotto un tavolo in modo da non avere un confronto visivo diretto. Inoltre era stato chiesto ai
soggetti di non muovere i piedi. I risultati mostrano che entrambi i tipi di pazienti erano in grado di
svolgere il compito ruotando anche l’arto colpito e non è stata riportata nessuna differenza
significativa per l’accuratezza e i tempi di reazione. Un risultato interessante riguarda la mancanza
di differenza nell’esecuzione del compito tra pazienti con amputazione dell’arto dominante e non
dominante. Questo dato è in contrasto con i risultati riportati da Nico (2004) sulla differenza
riscontrata tra soggetti con amputazione di arto dominante e quelli di arto non dominante.
Il paziente con giroplastica ha mantenuto la capacità di rotazione mentale nonostante fosse trascorsi
21 annidopo l’operazione. Dal momento che questo soggetto non aveva subito un’amputazione ma
una rotazione di 180° dell’arto, poteva ricevere input sensoriali dalla periferia. Questo ha fatto
ipotizzare che durante il compito il soggetto avrebbe avuto un comportamento diverso per quanto
riguarda la risposta all’orientamento rispetto ai pazienti amputati. La mancanza di differenze nei
risultati delle due comparazioni, è per alcuni spiegata dalla presenza di una rappresentazione innata
del corpo prototipico cervello (Berlucci e Aglioti, 1997).
La prova di questa rappresentazione innata viene da bambini con agenesia degli arti che riportando
sensazioni di arto fantasma confermano questa ipotesi (Melzack et al. 1997). Come suggerito da
Corradi-Dell’Acqua et al. (2008) esistono due rappresentazioni corporee nell’essere umano: la
prima elabora le informazioni del corpo in relazione allo spazio ed è denominata schema corporeo;
la seconda descrive la struttura del corpo comparandolo ad un corpo prototipico. La comprensione
della lateralità nasce presto nella vita degli individui ed è la prova che i soggetti sani utilizzano la
rappresentazione del proprio corpo per rispondere in modo corretto. Al contrario, i pazienti dello
studio di Curtze (2010) mostrando di trattare l’immagine del piede come un oggetto, utilizzando
una rappresentazione astratta del corpo, una trasformazione mentale visiva per far fronte al compito.
23
Questi studi suggeriscono che l’amputazione di un arto comporta cambiamenti nella
rappresentazione dello schema corporeo che costringe i pazienti ad utilizzare una diversa strategia
per l’esecuzione di compiti di immagine mentale.
2.4 Cambiamenti strutturali in seguito a cambiamenti periferici.
L’amputazione di un arto comporta modificazioni a carico del sistema nervoso centrale e
spesso la comparsa di sensazioni a carico dell’arto mancante (arto fantasma). Quest’ultime si
manifestano differentemente negli individui ed è definito come dolore da arto fantasma. Si presenta
con sensazioni di bruciore, crampi (Giummarra et al. 2007), che devono essere differenziati dalle
sensazioni dolorose inerenti al moncone. Infatti i soggetti esperiscono la presenza dell’arto fantasma
per periodi piuttosto lunghi dopo l’amputazione. Alcuni studi hanno dimostrato che il dolore
cronico da arto fantasma coincide con sostanziali cambiamenti strutturali in corteccia (May 2011).
Diversi autori nelle analisi strutturali delle aree cerebrali riportano casi di diverse modificazioni con
riduzione o aumento della materia grigia in diversi pazienti con amputazione degli arti. Alcuni
autori hanno individuato riduzioni dello spessore cerebrale in diverse regioni quali: corteccia
insulare, corteccia cingolata e corteccia frontale (Treede et al. 1999). E’ stato riportato un singolo
caso in cui la perdita di arti superiori in un lasso di tempo di 21 settimane, ha portato ad una
graduale riduzione di materia grigia nelle aree quali: corteccia somatosensoriale primaria (S1),
corteccia motoria primaria (M1), corteccia parietale superiore e area motoria supplementare, (Gaser
et al . 2004). La riorganizzazione della S1 sembrerebbe svolgere un ruolo principale nel
mantenimento dell’arto fantasma (Flor et al 1995, 2006;. Birbaumer et al 1997;. Preißler et al 2011).
Draganski et al. (2006) hanno invece trovato che i soggetti con amputazione di arto superiore o
inferiore, mostravano lo stesso tipo di riduzione dello spessore corticale a livello del talamo
controlaterale al lato dell’amputazione. Preißler et al. (2013) hanno effettuato uno studio più
specifico comparando le strutture cerebrali di pazienti amputati con dolore di arto fantasma lieve
(SPLP) e soggetti con dolore di arto fantasma medio-alto (HPLP) e infine è stato comparato il
cervello di entrambi i gruppi con uno di controllo di soggetti sani. L’arto amputato e la sensazione
di dolore dell’arto fantasma, sono entrambi associati ad una riorganizzazione strutturale del sistema
nervoso centrale. L’alterazione causata dall’amputazione di un arto è supportato da modifiche
corticali globali in cui si evidenzia un ridotto volume di materia grigia nei pazienti con amputazione
dell’arto superiore destro, rispetto al gruppo dei soggetti sani, nella corteccia motoria di sinistra
(M1) che rappresenta l’arto amputato. Inoltre i pazienti mostrano un aumento della materia grigia
24
nelle regioni della via visiva ventrale e dorsale. Dall’analisi VBM (voxel based morphometry) i
pazienti hanno mostrato una significativa riduzione totale della materia grigia. Ciò è probabilmente
dovuto ad una mancanza di input sensoriali che ha portato ad una riorganizzazione strutturale nel
cervello adulto (Pascual- Leone et al 1993, 2005;. Elbert et al 1994;. Flor et al 1995;. Weiss et al.
2004; Draganski et al 2006). Le alterazioni verificatesi nel giro precentrale hanno mostrato una
riduzione strutturale a livello della corteccia motoria primaria di sinistra. Questa zona sembrerebbe
coincidere con la regione motoria della mano rimossa (Preißler et al., 2013). Nonostante la
moltitudine di aree strutturalmente modificate trovate in studi precedenti, la riduzione di materia
grigia nella corteccia motoria di sinistra coincide con risultati ottenuti da altri autori. Dettmers et al.
(1999) ad esempio, hanno avuto prove mediante la valutazione di bambini con amputazione
dell’arto nei primi anni di vita che hanno mostrato alterazioni a livello della M1. Questi risultati
indicano che con il passare del tempo, la parte cerebrale che codificava l’azione della parte
amputata ha gradualmente perso la sua funzionalità portando ad una riduzione di materia grigia in
quell’area. Oltre ad una riduzione di materia grigia l’analisi VBM ha mostrato alterazioni
anatomiche della via visiva con aumento di diverse parti del lobo parietale superiore e del lobo
temporale dei pazienti amputati, precisamente nel lobo temporale, nel lobo temporale centrale e
nella corteccia fusiforme. Queste aree comprendono il circuito visuo-motorio che si attiva mediante
due vie (Mishkin, Ungerleider e Macko, 1983; Rossetti et al 2003;. Cardoso-Leite e Gorea 2010):
la prima è quella dorsale che comprende l’area occipitale, temporale posteriore e le regioni parietali
responsabili dell’orientamento spaziale degli oggetti e nel reperire informazioni circa il movimento.
La seconda via invece è coinvolta nella riconoscimento dell’oggetto e comprende il lobo occipitale
e temporale (Ellison e Cowey 2006, 2007, 2009; Cavina- Pratesi et al 2007;. Rice et al 2007;..
Cohen et al 2009). I risultati dell’analisi VBM hanno rilevato un’alterazione di entrambe le vie
visive con aumento di queste zone corticali. L’aumento di materia grigia è in corrispondenza delle
regioni che ricevono input visivi (Cardoso- Leite e Gorea 2010). E’ probabile che avvenga una
compensazione attraverso un utilizzo maggiore della vista. E’ stato osservato che i soggetti con
protesi mioelettrica facevano un continuo ricorso al controllo visivo dell’azione della mano
artificiale. Questa necessità di costante utilizzo della vista potrebbe portare ad un fenomeno di
plasticità cerebrale che spiegherebbe l’aumento della materia grigia. La riorganizzazione strutturale
causata da dolore dell’arto fantasma, invece, implica un’alterazione anatomica della via visiva. In
pazienti con dolore di arto fantasma cronico medio-alto è stata riscontrata una riduzione di materia
grigia nella via visiva ventrale, contrariamente a quanto emerso in pazienti con dolore lieve.
25
Preißler et al. (2013) hanno ipotizzato che questa differenza sia probabilmente dovuto ad una
riduzione di movimento del moncone o della protesi a causa del forte dolore percepito. Questo si
traduce in una mancata elaborazione visiva che non porta ad un riadattamento cognitivo come
succede nei pazienti con SPLP. Quindi se lo spostamento dell’attenzione dalla modalità
somatosensoriale a quella visiva fallisce, i soggetti con HPLP non riuscendo a trovare un riscontro
né dalla periferia, né a livello cognitivo avranno di conseguenza un aumento del dolore dell’arto
fantasma (Melzack 1990, 2005). Questa ipotesi è in linea con i dati riportati da Ramachandran et al
(1996) e Ramachandran e Rogers-Ramachandran (2006) attraverso una tecnica riabilitativa che
utilizzava una visual-box per ridurre il dolore dell’arto fantasma attraverso il feed-back visivo.
Questo sottolinea l’importanza del processo visivo nella riorganizzazione corticale dopo aver subito
la perdita di una parte del proprio corpo. Infine è stata rilevata anche un’alterazione strutturale del
circuito deputato all’elaborazione del dolore. L’analisi VBM ha mostrato una riduzione di materia
grigia nella corteccia orbito-frontale. Quest’area è coinvolta nell’elaborazione delle informazioni
nocicettive, dolorifiche e inoltre è importante per l’elaborazione dei premi e delle punizioni.
L’attività di questa regione media il ''valore affettivo personale'' dei rinforzi (Kringelbach e Rolls
2004). “Si presume che il valore affettivo negativo di uno stimolo doloroso o un'esperienza
dolorosa cronica può essere codificato in questa regione” (Preißler et al., 2013). Inoltre è stata
trovata un’altra area con modificazioni strutturali, la corteccia prefrontale dorsolaterale,
maggiormente a destra. Anche questa zona è implicata nell’elaborazione del dolore, in particolare
nell’elaborazione del dolore cronico. Sussistono differenze a livello di modificazione strutturale in
quest’area tra pazienti con dolore da arto fantasma cronico e pazienti con dolore lieve. La riduzione
è avvenuta solo nei pazienti con dolore da arto fantasma cronico. Infine i risultati hanno mostrato la
riduzione di un’altra struttura, ossia, il collicolo inferiore centrale e posteriore dei pazienti con
dolore dell’arto fantasma medio-alto rispetto ai pazienti con dolore lieve. Attraverso la VAS rating
è stato dimostrato che più il paziente percepiva il dolore, maggiore era la riduzione di materia grigia
nel collicolo inferiore di destra. Differentemente dai pazienti con SPLP, quelli con HPLP vanno
incontro ad una plasticità “disadattiva” attraverso l’attivazione di aree che elaborano il dolore (Katz
e Melzack 1990;. Apkarian et al 2009). La sensazione persistente di dolore porta nel tempo ad un
processo degenerativo, con conseguente morte neuronale, riduzione di dendriti, riduzione delle
dimensioni delle cellule, fino alla riduzione della regione corticale deputata alla codifica visiva
(Draganski e May 2008; May 2008, 2009). Tuttavia, diversamente dai pazienti con SPLP, in questi
pazienti si riscontra un aumento di materia grigia nella corteccia cingolata anteriore (ACC). È da
26
notare che quest’area si attiva in risposta a stimoli emotivamente salienti, come ad esempio il dolore
(Bentley et al. 2003), inibisce il movimento, le risposte e valuta il livello di dolore percepito. Studi
di neuroimaging funzionale hanno dimostrato un’attivazione dell’ACC in risposta a stimoli
nocicettivi (Peyron et al. 2000). Preißler et al. (2013) hanno suggerito due possibilità alla base di un
maggior volume corticale nella corteccia cingolata anteriore. La prima ipotizza che l’aumento di
volume della ACC prima dell’operazione chirurgica, possa essere un fattore di vulnerabilità per lo
sviluppo di un forte dolore cronico post operazione. La seconda ipotesi è che questo aumento sia
dovuto ad una costante sensazione negativa prodotta dal dolore permanente o dal costante tentativo
da parte del paziente di evitare il dolore, oppure l’insieme di entrambe le possibilità (Preißler et
al.,2013). Complessivamente queste modificazioni strutturali di materia grigia possono essere
interpretate in termini di atrofia cerebrale collegata al dolore (Apkarian et al 2004;. Rocca et al
2006;. Kim et al 2008).
Langer et al. (2012), hanno effettuato uno studio per verificare come anche cambiamenti transitori
della periferia del corpo possano indurre meccanismi di compensazione e plasticità a livello
cerebrale. Lo studio è stato effettuato mediante l’immobilizzazione dell’arto superiore destro per 14
giorni e la compensazione con l’arto sinistro (non dominante) per mediare le funzioni svolte
quotidianamente dall’arto dominante. Gli autori ipotizzavano che una riduzione della mobilità
dell’arto superiore destro inducesse una diminuzione di volume (misurato attraverso Surface-based
morphometry analysis) delle aree somatosensoriali e dell’area motoria primaria e dei fasci di fibre
all’interno del tratto corticospinale (misurata attraverso l’analisi delle mappe di Fractional
Anisotropy, FA), deputata all’elaborazione dei movimenti fini. Al contrario, la compensazione per
opera dell’arto superiore di sinistra avrebbe dovuto comportare un aumento di materia grigia a
livello dell’area sensorimotoria dell’emisfero destro e anche un aumento di FA nel tratto
corticospinale di destra. Mediante test di valutazione di abilità motorie lo studio ha dimostrato un
effettivo miglioramento della mano non dominante e questo effetto si evidenzia anche a livello
corticale. Infatti, è stato riscontrato un aumento di materia grigia nella corteccia motoria e
premotoria di destra. Tale cambiamento strutturale può riflettere o un’acquisizione di nuove abilità o
la conseguenza di una costante stimolazione dell’arto superiore di sinistra durante i 14 giorni di
immobilizzazione del braccio destro (Langer et al., 2012). La precoce plasticità cerebrale evidenzia
che il sistema nervoso si evolve per far fronte a improvvisi cambiamenti ambientali. Questi risultati
sono importanti perché possono indirizzare futuri approcci terapeutici per la compensazione
cerebrale in pazienti con danni centrali o periferici (Langer et al., 2012).
27
CAPITOLO 3
Rappresentazione corporea: uno studio fMRI in soggetti con
alterazioni periferiche.
3.1. Introduzione
La comprensione dei meccanismi attraverso i quali il cervello rappresenta il corpo è uno
degli oggetti d’indagine che più ha stimolato le neuroscienze negli ultimi anni.
Il nostro corpo è, infatti, un oggetto speciale d’indagine poiché i modi in cui entriamo in
relazione con esso sono molteplici: il tatto, la vista, la propriocezione, il comportamento motorio, la
comprensione semantica, le emozioni, i sentimenti. Riceviamo quindi molte più informazioni dal
nostro corpo che da qualsiasi altro oggetto è, inoltre, l’unico che percepiamo dall’interno
(deVigmenount, 2011).
In altri termini, il nostro cervello elabora costantemente informazioni, sia top-down che
bottom-up, riguardanti il nostro corpo che interagiscono per creare rappresentazioni più astratte del
corpo. La complessità di modi in cui entriamo in relazione con il nostro corpo rende quindi ragione
della varietà di modi in cui il cervello rappresenta il corpo e della varietà di alterazioni delle
rappresentazioni corporee che possono conseguire sia a lesioni cerebrali sia alla perdita di
informazioni derivante da deafferentazione sensorimotoria.
Per quanto concerne nello specifico quest’ultimo caso, in seguito a modifiche delle afferenze
somatiche o dell’output motorio possono verificarsi cambiamenti neuroplastici in aree
sensorimotorie del sistema nervoso (Jones, 2000; Sanes e Donoghue, 2000). Se, da una parte,
infatti, l’apprendere nuove abilità motorie determina un ampliamento della rappresentazione dei
muscoli coinvolti in un certo compito (Pascual-Leone et al., 1995) e cambiamenti strutturali in
specifiche aree cerebrali (Draganski et al., 2004), dall’altra, una deafferentazione a lungo termine di
tipo sensorimotorio, come nel caso di amputazione di un arto, risulta in un’espansione delle
rappresentazioni circostanti (Chen et al., 1998; Cohen et al., 1991) e nella riduzione di materia
grigia (Draganski et al., 2006).
Diverse prove sperimentali sono state raccolte sulla neuroplasticità conseguente a
deafferentazione sensorimotoria e sulle conseguenti modificazioni della rappresentazione corporea.
Nella maggior parte dei casi si tratta di studi comportamentali che hanno valutato
modificazioni nella rappresentazione corporea attraverso compiti di rotazione mentale di parti del
corpo (es. mani/piedi). Questi compiti richiedono, ad esempio, di stabilire se la figura di una mano
28
ruotata di una certa angolazione è una mano destra o sinistra (Parsons, 1987). Inoltre, per essere
risolti correttamente, richiedono un’implicita rotazione della propria parte del corpo (Parsons,
1987), valutano quindi una specifica rappresentazione corporea, cioè lo “schema corporeo” o
rappresentazione sensorimotoria del nostro corpo (Schwoebel & Coslett, 2005). Studi
neuropsicologici mostrano, infatti, che l’immaginare di compiere un movimento richiede il
richiamo dello schema corporeo esattamente come se si stesse realmente svolgendo il movimento
(de Lange et al., 2005; Parsons, 1994; Schwoebel et al., 2001). Inoltre, studi di neuroimaging
funzionale confermano tale parallelismo tra azione immaginata e realmente eseguita poiché
mostrano sovrapposizioni nelle attivazioni della corteccia parietale superiore, nel solco
intraparietale e nella porzione più rostrale del lobulo parietale inferiore (Bonda et al., 1995; Parsons
et al., 1995) durante i due compiti.
Attraverso l’utilizzo di un compito di rotazione di mani, Nico e colleghi (2004) trovano che i
pazienti che hanno subito l’amputazione di un arto superiore sono più lenti e meno accurati in un
compito di rotazioni mentali di mani rispetto a individui sani di controllo. Gli autori ipotizzano una
modificazione nello schema corporeo per spiegare l’aumento dei tempi di risposta e la minore
accuratezza; i soggetti amputati non possono più confrontare la rappresentazione del proprio arto
con il disegno della mano presentato.
Curtze e colleghi (2010) trovano, invece, che negli amputati, a differenza dei controlli, non è
presente l’effetto di lateralità. In altri termini, i soggetti di controllo sono più veloci nel giudicare
l’arto dominante ma questo effetto si perde in seguito ad amputazione. Secondo gli autori, tale dato
suggerisce che gli amputati non utilizzano nello svolgimento del compito la rappresentazione del
proprio corpo (schema corporeo), ma una strategia visiva che fa riferimento a una rappresentazione
più astratta e prototipica del corpo che è stata definita da alcuni autori “descrizione strutturale del
corpo” (Corradi Dell’Acqua et al., 2009), ovvero, una “mappa visuo-spaziale” dei diversi distretti
corporei. Ad ogni modo, l’elicitazione, durante il compito, dell’arto fantasma in alcuni dei soggetti
amputati suggerisce che almeno questi soggetti abbiano utilizzato l’imagery motoria. In questa
condizione il soggetto pensa a ciò che deve fare, immagina la sequenza delle azioni da svolgere e ne
vede il risultato finale. E’ dunque un’attività immaginativa in cui il compito viene svolto, in assenza
di movimento osservabile, includendo una rappresentazione egocentrica del proprio corpo mediante
l’utilizzo dello schema corporeo. Studi condotti da Corradi-Dell’Acqua (2009) per l’identificazione
delle differenti basi neurali dello schema corporeo (BD) e dello schema strutturale del corpo (BSD)
sostengono questa ipotesi. Studi fMRI hanno mostrato che i soggetti, durante i compiti di rotazione
mentale, possono utilizzare due diverse strategie: la prima riguarda una rappresentazione
egocentrica (BD) in cui il soggetto codifica la posizione delle parti del proprio corpo in relazione
29
con l’ambiente; la seconda strategia fa riferimento ad una rappresentazione allocentrica in cui i
soggetti codificano la posizione delle parti del proprio corpo mediante il confronto con un corpo
standard (BSD) (Corradi-Dell’Acqua, 2009). Altri studi hanno dimostrato che i soggetti possono
eseguire un compito su parti del corpo mediante l’utilizzo di strategie diverse. Ad esempio Parsons
(1987) ha notato che i partecipanti ad un compito di rotazione mentale di mani confrontavano
spontaneamente la parte del corpo presentata con la parte del proprio corpo mediante l’uso
dell’imagery motoria. Tuttavia questo compito poteva essere risolto anche mediante una diversa
strategia, con l’utilizzo di immagini visive richiamando una rappresentazione allocentrica (Kosslyn
et al, 2001; Tomasino e Rumiati, 2004).
Sebbene compiti di rotazione mentale di parti del corpo siano stati utilizzati in studi di
neuroimaging funzionale per localizzare la sede delle diverse rappresentazioni corporee (Schwoebel
& Coslett, 2005; Corradi-Dell’Acqua et al., 2009), a nostra conoscenza nessun lavoro di
neuroimaging funzionale ha indagato se la differenza nella prestazione in compiti di rotazione
mentale di parti del corpo tra soggetti di controllo e amputati riportata in alcuni studi
comportamentali (Nico et al., 2004; Curtze et al., 210) sia da imputare a una modificazione dello
schema corporeo o all’utilizzo di una diversa strategia, che presuppone il richiamo di una diversa
rappresentazione corporea.
L’obiettivo del nostro studio è indagare, attraverso uno studio di neuroimaging funzionale, i
pattern di attivazione di pazienti con amputazione di un arto durante un compito di rotazione di parti
del corpo, al fine di verificare se queste attivazioni siano o meno sovrapponibili a quelle di soggetti
di controllo. Questo confronto ha permesso di verificare se la perdita d’informazioni
sensorimotorie, conseguenti all’amputazione, ha determinato cambiamenti nelle attivazioni delle
aree deputate all’elaborazione dello schema corporeo e/o l’attivazione di altre aree cerebrali
coinvolte nell’elaborazione di altre rappresentazioni corporee (ad esempio quelle deputate
all’elaborazione della “mappa visuo-spaziale del corpo”) che indicherebbero il ricorso a una
differente strategia per eseguire il compito. Inoltre abbiamo indagato attraverso un’analisi Voxel
Based Morphometry (VBM) se la perdita di informazioni sensorimotorie potesse modificare la
corteccia a livello strutturale.
Lo studio è stato condotto presso la Fondazione Santa Lucia ed è stato approvato dal
comitato etico indipendente della struttura.
30
3.2 Metodologia
Soggetti
Gruppo sperimentale
Un gruppo di 9 pazienti maschi con amputazione dell’arto sinistro, ovvero l’arto non dominante (età
media 40,88; Deviazione Standard-DS 19,68) e senza storia di disturbi neurologici e/o psichiatrici,
è stato coinvolto nello studio. Ad ogni partecipante è stata richiesta un’adesione scritta di
partecipazione alla ricerca (consenso informato -D.lgs. del 30 giugno 2003, n. 196).
Gruppo di controllo
Un gruppo di 11 volontari maschi sani (età media 41,18 e DS 15,73), senza storia di disturbi
neurologici e/o psichiatrici, paragonabili, età e scolarità al gruppo sperimentale (Età: T= 0.037, p=
0.970; Scolarità: T= 0.765, p=0.455), è stato coinvolto nello studio. Ad ogni partecipante è stata
richiesta un’adesione scritta di partecipazione alla ricerca (consenso informato -D.lgs. del 30 giugno
2003, n. 196) (si veda la tabella 1 per le informazioni demografiche).
Soggetti Età
Controlli 41.18 (15.73)
Amputati 40.88(19.68)
Scolarità
14.72 (2.79)
13.55 (4.06)
P.E. raven
33.54 (2.20)
30.66 (4.38)
Tabella 1. Media e DS del campione di ricerca.
Stimoli
Al fine di indagare eventuali modifiche nella rappresentazione mentale del corpo (schema corporeo)
nei pazienti con amputazione dell’arto inferiore sinistro, è stato sviluppato un set di stimoli
costituito da stimoli inerenti a parti del corpo (mano e piede) ed oggetti non corporei (specchietto
di motocicletta) che hanno una destra e una sinistra intrinseca; gli stimoli (mano, piede e specchietto
destro e sinistro) sono stati ruotati di 60 gradi in 60 gradi, ottenendo così per ogni categoria 36
possibili rotazioni, da 0° a 300°(Figura 1).
31
Orientamento Mano Destra__________________Orientamento Mano_Sinistra______
Orientamento_Specchietto_Destro____________Orientamento_Specchietto_Sinistro__
Orientamento_Piede_Destro__________________Orientamento_Piede_Sinistro_____
Figura 1. Set di stimoli utilizzati nell’esperimento.
32
Il set di stimoli utilizzati nell’ambito del paradigma sperimentale di fMRI comprende quindi 3
categorie: arti superiori (mano), arti inferiori (piede) e un oggetto non corporeo (specchietto di una
moto). Più nello specifico, il set di stimoli comprende 72 item per gli arti superiori (36 stimoli
relativi alla mano destra e 36 alla mano sinistra), 72 per gli arti inferiori (36 stimoli relativi al piede
destro e 36 al piede sinistro) e 72 item che rappresentano un oggetto non corporeo che può essere
ruotato realmente (36 stimoli relativi allo specchietto destro e 36 allo specchietto sinistro).
Inoltre sono stati utilizzati test comportamentali carta e matita per indagare eventuali
modificazioni in altre rappresentazioni corporee (rappresentazione semantica e mappa visuospaziale).
Test
Versione modificata del test di Semenza e Goodglass (1985)
Il test è costituito da 30 item, in ciascuno dei quali si chiede al soggetto di indicare le parti del
corpo sia destre che sinistre denominate dall’esaminatore, prima ad occhi chiusi e successivamente
ad occhi aperti (dopo un intervallo durante il quale il soggetto è impegnato in un altro compito). È
stato calcolato il numero di errori.
Test dello Schema Corporeo (Durat-Hmeljiak et al., 1978)
Il test si compone di diversi subtest. Nel nostro paradigma abbiamo utilizzato il subtest di
evocazione del corpo visto di fronte, il cui materiale è costituito da una tavoletta di lavoro su cui è
raffigurata la testa di un bambino e 9 tessere, ciascuna rappresentante una parte del corpo umano
(collo, braccio destro e sinistro, gamba destra e sinistra, emitorace destro e sinistro, mano destra e
sinistra). E’ stato chiesto al soggetto di riconoscere, nominandoli e indicandone la lateralità, le
diverse parti del corpo per poi posizionarle sulla tavoletta in cui l’unico punto di riferimento è la
testa del bambino. Si è registrato il tempo di posizionamento della parte del corpo e prima di
presentare una nuova figura, la posizione della tessere è stata registrata sovrapponendo una griglia
sulla tavoletta. Ogni tessera viene rimossa prima di presentare la successiva. Il punteggio è dato
dalla somma delle risposte corrette al riconoscimento (punteggio massimo: 9) e alla localizzazione
(punteggio massimo: 9).
Test di semantica
Ogni trial del test è costituito da immagini rappresentanti parti del corpo da abbinare ad
immagini rappresentanti capi di abbigliamento. La risposta è stata classificata per: contigua,
concettuale, random e corretta. Ai soggetti venivano presentate, una per volta, immagini raffiguranti
33
capi di abbigliamento per ciascuno dei quali il soggetto doveva indicare tra le quattro alternative
raffiguranti parti del corpo, quale tra le immagini presentate corrispondeva allo specifico capo di
abbigliamento.
Procedura
Per ciascun soggetto la sessione sperimentale fMRI è stata preceduta da una breve sessione di
addestramento all’esterno della macchina al fine di permettere la comprensione e familiarizzazione
con il compito. Lo studio fMRI è stato progettato come un disegno a eventi. La presentazione del
set di stimoli è stata randomizzata e ciascun item è stato presentato per un massimo di 3000 msec,
seguito da un punto di fissazione di durata variabile tra 1300 e 3100 ms (Figura 2). Durante
l’acquisizione delle immagini in risonanza è stato richiesto ai partecipanti allo studio di eseguire un
compito di giudizio di lateralità dello stimolo (destra/sinistra) attraverso l’utilizzo di una pulsantiera
compatibile con l’ambiente di risonanza.
Sono state acquisite 5 sessioni fMRI, ciascuna della durata di 9 minuti circa.
Figura 2. Paradigma Sperimentale
Acquisizione delle immagini
Lo studio è stato eseguito presso il servizio di Radiologia della Fondazione Santa Lucia
utilizzando uno scanner di risonanza magnetica a 3 Tesla Siemens “Allegra”
Le immagini sono state acquisite per l’intera corteccia usando l’imaging BOLD (30 fette, in-plane
risoluzione = 3 x 3 mm, slice spacing = 4.5 mm, tempo di ripetizione è [TR] = 2 sec,tempo di eco [
TE ] = 30 msec, flip angle = 70 gradi); per ognuna delle 5 scansioni fMRI sono stati acquisiti 276
volumi. Inoltre le scansioni funzionali sono state seguite dall’acquisizione di scansioni anatomiche
ad alta risoluzione pesate in T1 (Siemens MPRAGE, 176 fette, piano di risoluzione = 0,5 x 0,5 mm,
spessore fetta = 1 mm, TR = 2 sec, TE = 4.38 ms, flip angle= 8 gradi); le immagini strutturali
tridimensionali ad alta risoluzione sono state utilizzate per la co-registrazione delle immagini EPI,
34
permettendo una localizzazione anatomica più precisa delle attivazioni. La durata complessiva di
ogni acquisizione non superava i 60 minuti.
Preprocessamento delle immagini
I
dati
di
risonanza
sono
stati
pre-processati
mediante
il
software
SPM8
(http://www.fil.ion.ucl.ac.uk/spm) combinato con un software interno (BrainShow) implementato in
MATLAB (The MathWorks Inc., Natick, MA, USA). I primi quattro volumi di ogni scansione sono
stati scartati per consentire la stabilizzazione del campo magnetico. Tutte le immagini sono state
corrette per i movimenti della testa (realignment) usando il primo volume di riferimento. Le
immagini di ogni partecipante sono state poi co-registrate sulla loro stessa immagine in T1. Le
immagini co-registrate sono state poi normalizzate utilizzando MNI-152 EPI standard e l’immagine
riallineata media come fonte. È stato infine applicato un processo di smoothing utilizzando un
kernel isotropo Gaussiano di 8 mm.
3.3 Analisi dei test carta e matita.
I test comportamentali carta e matita (versione modificata del test di Semenza e Goodglass,
sub-test Evocazione Fronte Corpo, test di semantica) sono stati somministrati ai soggetti prima
dello svolgimento del compito di rotazione mentale in risonanza. Al fine di individuare differenze
significative tra il gruppo di controllo e il gruppo sperimentale, è stata effettuata un’analisi T-test.
3.4 Analisi dei dati comportamentali.
I dati comportamentali del paradigma di rotazioni mentali in fMRI sono stati analizzati per
mezzo di ANOVA a struttura fattoriale mista, sull’accuratezza e sui tempi di risposta. I dati sono
stati dapprima filtrati per i tempi, eliminando le risposte < 300 ms (Nico et al., 2004) ed è stata
calcolata la proporzione di risposte corrette per ogni ripetizione di ogni item ed è stata applicata una
trasformazione per Arcoseno (Nico et all., 2004). Successivamente abbiamo filtrato le risposte
corrette e calcolato la media dei tempi di risposta, per ciascuna condizione in ciascun soggetto. I
valori mancanti sono stati sostituiti con la media dei tempi di risposta del gruppo in quella
condizione. È stata quindi applicata una trasformazione logaritmica.
Abbiamo eseguito due ANOVA con struttura fattoriale mista 3×2×6×2 con categoria
(mano/piede/oggetto), lato (destra/sinistra), angolo di rotazione (0°/60°/120°/180°/240°/300°) come
fattori within-subject e il gruppo (controllo/sperimentale) come fattore between-subject sia
sull’accuratezza che sui tempi di risposta. In base ai dati presenti in letteratura ci aspettiamo che i
35
pazienti con amputazione dell’arto inferiore sinistro mostrino un rallentamento nell’esecuzione del
compito di rotazione mentale riguardante la categoria che rappresenta l’arto amputato. Se l’ipotesi è
corretta, dovremmo osservare un effetto d’interazione tra il gruppo e la categoria, con effetti
specifici dell’arto amputato.
I confronti post hoc sono stati condotti utilizzando la correzione di Bonferroni per i
confronti multipli, sia per quanto riguarda le analisi sull’accuratezza che sui tempi di risposta.
3.5 Analisi dei dati fMRI
Le immagini funzionali sono state analizzate separatamente per ciascun partecipante
utilizzando un’analisi voxel per voxel in accordo con il General Linear Model (Nelder,
Wedderburn, 1972). Abbiamo inserito dei regressori per ogni condizione sperimentale, mano
destra-mano sinistra, piede destro-piede sinistro, sottraendo lo stimolo di controllo, ovvero, lo
specchietto. L’analisi di gruppo è stata condotta sulle immagini così stimate relativamente alle
singole condizioni sperimentali, trattando il soggetto come un fattore casuale. In particolar modo, a
livello del gruppo, abbiamo condotto dei Two Sample T-test per confrontare l’attivazione dei due
gruppi (controlli, pazienti con amputazione dell’arto inferiore di sinistra) sulle singole condizioni
(mano destra-sinistra, piede amputato-non amputato). Data la scarsa numerosità dei soggetti in
questo primo stato di analisi, abbiamo deciso di applicare una soglia non troppo conservativa
(punc<0.001) e una dimensione del cluster maggiore di 10 voxel.
Per indagare eventuali differenze strutturali tra pazienti con amputazione dell’arto non
dominante e soggetti di controllo abbiamo condotto una Voxel Based Morphometry (VBM) sulle
immagini MPRAGE utilizzando VBM 8 toolbox, implementata in SPM 8. La presenza di artefatti
grossolani e anormalità è stata controllata da un esperto di neuroimmagini. Nell’analisi sono stati
inclusi 8 amputati sinistri e 11 soggetti di controllo. Le immagini di un partecipante sono state
escluse da questa analisi poiché non rispettavano i parametri della qualità dell’immagine per
l’esecuzione della VBM (esse si discostavano di 2 deviazioni standard rispetto alla media). Le
immagini sono quindi state normalizzate utilizzando la normalizzazione di Dartel, segmentate in
materia grigia, materia bianca e liquor cerebrospinale; dunque è stato effettuato un processo di
smoothing utilizzando un full width at half minimum (FWHM) a 8 mm. Abbiamo utilizzato un two
sample T-test per comparare le immagini dei pazienti amputati con quelle dei soggetti di controllo.
36
3.5 Risultati
T-test nei compiti carta e matita
L’analisi dei dati comportamentali, mediante T-test, atta ad individuare eventuali
modificazioni nella rappresentazione corporea all’interno del gruppo sperimentale confrontato con
quello di controllo non ha evidenziato risultati significativi.
ANOVA: accuratezza
L’ANOVA mista 3×2×6×2 con categoria (mano/piede/oggetto), lato (destra/sinistra), angolo
di
rotazione
(0°/60°/120°/180°/240°/300°)
come
fattori
within-subject
e
il
gruppo
(controllo/sperimentale) come fattore between-subject, è stata eseguita in un primo momento
sull’accuratezza per testare se ci fossero differenze significative tra i due gruppi. Dai risultati è
emerso un effetto principale significativo per il fattore categoria con [F(2; 38) =7.839, p=0.001].
L’analisi post-hoc mostra una maggior accuratezza dei soggetti per la categoria piede rispetto alla
categoria oggetto evidenziando una facilitazione nel riconoscimento della prima p=0.004 (Tabella
2).
Gruppo
Amputati Sx
Controlli
Categoria
Mano
Oggetto
Piede
Mano
Oggetto
Piede
Media
1.148
1.036
1.279
1.318
1.180
1.424
Deviazione Standard Errore
0.103
0.135
0.076
0.098
0.129
0.072
Tabella 2. Media e Deviazione standard del gruppo sperimentale e del gruppo di controllo per il
fattore categoria nell’analisi dell’accuratezza.
È inoltre significativo l’effetto principale rotazione [F (2.465; 46.833 )=10.634, p=0.000]. I
confronti post-hoc mostrano una maggior accuratezza dei soggetti in rotazioni di 0° rispetto alla
rotazione di 120°(p=0.036) e 180°(p=0.000); un’accuratezza maggiore a 60° rispetto a
180°(p=0.004); una minor accuratezza a 120° rispetto a 0°(p=0.036); una minor accuratezza a 180°
rispetto a 0°(p=0.00), 60°(p=0.004), 240°(p=0.044) e a 300°(p=0.000); una maggior accuratezza a
37
240° rispetto a 180°(p=0.044) e infine una maggior accuratezza a 300° rispetto a 180°(p=0.000)
(Figura 3). Inoltre è stato riscontrato un effetto di interazione significativo tra i fattori categoria ×
lato × rotazione [F (5.7; 108.295) =2.607, p=0.023]. L’analisi post hoc mostra che nell’interazione
il fattore lato è significativo solo nelle categorie mano e piede e non nella categoria oggetto. Questo
risultato è in linea con la presenza dell’effetto dominanza che in letteratura si riscontra per le parti
del corpo ma non per gli oggetti (Courtze, 2010).
Figura 3. Accuratezza e TR nell’interazione mano × rotazione, oggetto × rotazione e piede ×
rotazione.
38
ANOVA: tempi di risposta.
Abbiamo
effettuato
un’ANOVA
(3×2×6x2)
con
categoria
(mano/piede/oggetto),
lato
(destra/sinistra), angolo di rotazione (0°/60°/120°/180°/240°/300°) come fattori within-subject e il
gruppo (controllo/sperimentale) come fattore between-subject.
I risultati ottenuti mostrano un effetto principale significativo per il fattore Categoria [F(2;
38)= 4561, p=0.017] e dall’analisi post-hoc si evidenzia che i soggetti sono più rapidi nella
condizione mano rispetto a piede (p=0.007) e non vi sono differenze significative rispetto all’
oggetto (Tabella 3).
Gruppo
Amputati Sx
Controlli
Categoria
Mano
Oggetto
Piede
Mano
Oggetto
Piede
Media
3.185
3.177
3.146
3.119
3.117
3.094
Deviazione Standard Errore
0.037
0.034
0.034
0.035
0.033
0.032
Tabella 3. Media e Deviazione standard del gruppo sperimentale e del gruppo di controllo per il
fattore categoria nell’analisi dei tempi di risposta.
L’effetto della rotazione risulta essere significativo [F(5; 95)=83.156, p=0.00]. Dai confronti posthoc si evidenzia, come per l’analisi dell’accuratezza, che i soggetti sono più veloci, nei gradi di
rotazione 0° e 300°, mentre risultano con tempi di risposta più lunghi nelle rotazioni a 180°.
Effettuando confronti post-hoc la differenza media è significativa a livello dell’angolo di 0° e gli
angoli 120°,180°e 240° con p=0.00. L’effetto di facilità è in linea con le analisi effettuate
sull’accuratezza (Tabella 4).
39
Gruppo
Amputati Sx
Controlli
Rotazione
0°
60°
120°
180°
240°
300°
0°
60°
120°
180°
240°
300°
Media
3.118
3.143
3.181
3.260
3.177
3.137
3.070
3.072
3.113
3.220
3.126
3.060
Deviazione Standard Errore
0.037
0.037
0.037
0.032
0.030
0.034
0.035
0.035
0.036
0.031
0.028
0.032
Tabella 4. Media e Deviazione standard del gruppo sperimentale e del gruppo di controllo per il
fattore rotazione nell’analisi dei tempi di risposta.
Infine abbiamo riscontrato un effetto significativo nell’interazione categoria × rotazione con
[F(5.725; 108.771)= 4.063, p=0.001]. Dai confronti post-hoc i risultati mostrano che i soggetti
impiegano molto più tempo nella risposta della categoria mano, oggetto e piede nella rotazione a
180° (Figura 3).
Aree attivate durante il compito di rotazione mentale.
Nel paradigma sperimentale utilizzato, mediante dei Two Sample T-test abbiamo confrontato
l’attivazione dei due gruppi (controlli, pazienti con amputazione dell’arto inferiore di sinistra) sulle
singole condizioni (mano destra-sinistra, piede amputato-non amputato). Nella condizione di
rotazione dell’arto amputato (ovvero del piede sinistro), si evidenzia un incremento del segnale
BOLD nel Giro Sopramarginale di sinistra, che si estende marginalmente fino al giro temporale
superiore, nell’Area Supplementare Motoria sinistra e nel Verme Cerebellare destro dei pazienti
amputati rispetto al gruppo di controllo (Figura 4; Tabella 5). Dagli altri confronti condotti sulle
rimanenti condizioni sperimentali (piede destro, mano sinistra e mano destra) non emergono
differenze significative tra pazienti e controlli.
40
Laterale_________________________________________________________________________
Mediale_________________________________________________________________________
Figura 4. Attivazioni cerebrali in pazienti con amputazione dell’arto inferiore sinistro durante il
compito di rotazione mentale.
41
Condizione Piede Sinistro (attivazione A>C)
Regione
Area motoria supplementare
Giro Sopramarginale
Giro temporale superiore
(posteriore)
Verme 4-5
cluster
equivk
14.00
30.00
10.00
T
5.102
4.775
4.586
p
p(unc)
0.000
0.000
0.000
x
mm
-3
-57
-48
y
mm
-10
-22
-31
z
mm
52
16
16
4.709
0.000
6
-55
-5
Tabella 5. Informazioni dettagliate rispetto all’attivazione delle aree cerebrali durante l’attività di
rotazione mentale. Risultati ottenuti a seguito dell’analisi condotta con Two Sample T-test
confrontando l’attivazione tra i due gruppi con punc<0.001 e una dimensione del cluster >10 voxel.
Voxel Based Morphometry
I risultati dell’analisi VBM mostrano in pazienti con amputazione dell’arto inferiore sinistro
un volume maggiore di materia grigia a livello del Giro angolare (43.5, -55.5, 33 e 49.5, -61.5,
34.5), del Giro Frontale Medio (30, 27, 43.5) e del Cuneo (18, -84, 40.5 rispetto ai controlli )
(Figura 5; Tabella 6).
Aumento materia grigia in Amputati sinistri
Regione
Cuneo
Giro Angolare
Giro Angolare
Giro Medio Frontale
cluster
equivk
55
108
11
T
4.965
4.537
4.297
3.973
p
p(unc)
0.000
0.000
0.000
0.000
x
mm
18
43.5
49.5
30
y
mm
-84
-55,5
-61,5
27
z
mm
40.5
33
34.5
43.5
Tabella 6. Informazioni dettagliate rispetto all’aumento del volume cerebrale in pazienti con
amputazione dell’arto inferiore sinistro.
42
Laterale_________________________________________________________________________
Mediale_________________________________________________________________________
Figura 5. Aree con aumento del volume corticale in pazienti con amputazione dell’arto inferiore
sinistro.
43
3.5 Discussione.
“La rappresentazione mentale del nostro corpo è un processo che richiede il reclutamento di
diverse informazioni propriocettive, enterocettive, visive e senso-motorie in interazione con
l’ambiente” (Palermo et al., 2014). Nel presente studio abbiamo indagato attraverso uno studio di
risonanza magnetica funzionale come la mancanza d’informazioni sensorimotorie, in seguito
all’amputazione di parti del corpo, in particolare dell’arto inferiore sinistro, modifichi la
rappresentazione corporea. Abbiamo indagato se i pattern di attivazione di pazienti con
amputazione di un arto durante un compito di rotazione di parti del corpo, fossero o meno
sovrapponibili a quelli di soggetti di controllo. Questo confronto ci ha permesso di verificare
l’effetto della perdita d’informazioni sensorimotorie, conseguenti all’amputazione, sulla
rappresentazione neurale dello schema corporeo o l’attivazione di altre aree cerebrali coinvolte
nell’elaborazione di altre rappresentazioni corporee (ad esempio quelle deputate all’elaborazione
della “mappa visuo-spaziale del corpo”) che indicherebbero il ricorso a una differente strategia per
eseguire il compito.
Abbiamo confrontato un gruppo di 9 soggetti con amputazione dell’arto inferiore di sinistra
con un gruppo di 11 soggetti sani di controllo attraverso compiti comportamentali carta e matita e
attraverso un compito di rotazione mentale in fMRI. I risultati ottenuti attraverso le analisi effettuate
sul gruppo sperimentale e il gruppo di controllo, non mostrano effetti significativi nello
svolgimento delle prove comportamentali carta e matita. Il confronto tra le prestazioni dei due
gruppi al sub test “Evocazione-Fronte corpo” non riporta un valore significativo. Questo dato si
riscontra in letteratura (Palermo et al., 2014) dove l’effetto significativo risulta solo nei pazienti con
amputazione dell’arto dominante e non per quelli di arto non dominante, a dimostrazione che
evidentemente solo nel primo caso può avvenire una modificazione della mappa topologica del
corpo. Nel nostro caso, come riportato anche nello studio di Palermo et al. (2014), il risultato nel
sub test “Evocazione-Fronte corpo” può suggerire che anche se avviene una riorganizzazione,
questa non è tale da poter modificare in modo significativo la mappa topologica del corpo.
Abbiamo valutato la presenza di eventuali differenze significative tra gruppo di controllo e
gruppo sperimentale in una prova di rotazione mentale di parti del corpo, mani e piedi. I risultati
mostrano una maggior accuratezza dei soggetti per la categoria piede rispetto alla categoria oggetto
evidenziando una facilitazione nel riconoscimento della prima; è stato riscontrato anche un effetto
principale significativo per il fattore rotazione in cui tutte e tre le categorie (mano, piede ed oggetto)
risultavano meno accurate per l’angolo di rotazione a 180°. Questo risultato è in accordo con i dati
riportati da Nico et al. (2004) che considerano l’angolazione di 180° per entrambi gli arti superiori,
come una posizione innaturale e difficile da raggiungere e mantenere e ciò può essere ipotizzato
44
anche per la rotazione del piede. Inoltre, è in accordo con i dati classici sulle rotazioni mentali che
indicano aumento progressivo dei tempi e diminuzione progressiva dell’accuratezza all’aumentare
dell’angolo di rotazione dalla posizione canonica. Infine nell’analisi dell’accuratezza abbiamo
riscontrato un effetto di interazione significativo di terzo livello (categoria × lato × rotazione), in cui
il fattore lato risulta significativo solo per le parti del corpo e non per l’oggetto. Questo dato è in
linea con il fatto che nel corpo è presente il fattore dominanza che si verifica molto presto nella
nostra vita. Il risultato rispetto alla lateralità degli stimoli mano e piede ci fornisce la prova che i
soggetti durante la rotazione mentale fanno riferimento ad una rappresentazione del proprio corpo
(Courtze, 2010).
L’ANOVA effettuata sui tempi di risposta mostra un effetto principale per il fattore
categoria, in cui i soggetti rispondono più velocemente quando viene presentato lo stimolo mano
rispetto allo stimolo piede. Inoltre è emerso un effetto significativo della rotazione. E’ emerso,
come per l’analisi dell’accuratezza, che i soggetti sono più veloci, e quindi trovano il compito più
semplice, nei gradi di rotazione 0° e 300°, mentre risultano con tempi di risposta più lunghi nelle
rotazioni a 180° per tutte e tre le categorie (mano, oggetto e piede). L’effetto di facilitazione
rispecchia i risultati dell’ANOVA per l’accuratezza in linea con i dati presentati da Nico et al.
(2004) in cui i soggetti erano più lenti nel raggiungimento della rotazione a 180°, considerato come
orientamento non naturale.
Le due ANOVA effettuate rispetto all’accuratezza e ai tempi di risposta non riportano un
main effect del gruppo. Questo è un dato che non ci sorprende in quanto i compiti sono stati
costruiti con una struttura semplice in modo tale da permetterne il completo svolgimento da parte di
entrambi i gruppi, al fine di valutarne le attivazioni. Differentemente da quanto riscontrato in
letteratura (Nico et al., 2004), gli amputati non differivano dai controlli nella prova di rotazione
mentale della parte del corpo amputata. Questo è probabilmente dovuto al fatto che nei soggetti del
nostro studio l’arto amputato non era quello dominante.
In genere, le differenze riportate nell’analisi dei dati fMRI, possono essere interpretate come
diverse strategie utilizzate dai pazienti amputati nell’affrontare il compito di rotazione mentale. In
questi pazienti, durante lo svolgimento delle prove, è evidente un coinvolgimento dello schema
corporeo che però presenta caratteristiche parzialmente diverse da quelle dei soggetti di controllo,
poiché si osserva un’attivazione a livello cerebrale maggiore nell’emisfero sinistro a livello
dell’Area Motoria Supplementare, del Giro Sopramarginale, del Giro Temporale superiore e del
Verme. In particolare, i risultati mostrano un incremento del segnale BOLD nella parte posteriore
dell’Opercolo Parietale, precisamente nel Giro Sopramarginale di sinistra che si estende
marginalmente fino al giro temporale superiore, nei pazienti con amputazione dell’arto inferiore
45
sinistro rispetto al gruppo di controllo, durante l’attività di rotazione mentale del piede sinistro.
Quando immaginiamo di compiere una data azione vari parametri fisiologici corporei si comportano
come se noi stessimo effettivamente eseguendo quella stessa azione (Decety, 1996). Confrontando
questo dato con la letteratura (Corradi-Dell’Acqua, 2009), possiamo ipotizzare che per la
risoluzione del compito di rotazione mentale i pazienti amputati utilizzano come i soggetti di
controllo la rappresentazione dello schema corporeo, ma utilizzano una strategia che coinvolge
maggiormente il processo di imagery motoria.
Inoltre si evidenzia un’attivazione dell’Area Supplementare Motoria nei pazienti amputati
legata probabilmente alla strategia utilizzata per la rotazione mentale di parti del corpo. La SMA è
deputata alla coordinazione e alla pianificazione dei movimenti complessi, comprende una
rappresentazione somatotopica del corpo. Uno studio sulla somatotopia funzionale attraverso l’uso
di compiti complessi (Cunningham, 2015), ha rilevato un’attivazione a livello della SMA in
corrispondenza di una rappresentazione specifica degli arti. L’autore conferisce alla SMA un ruolo
di primo piano nell’esecuzione e nel controllo posturale dei movimenti prossimali e assiali. Nel
nostro studio, l’incremento del segnale BOLD in quest’area potrebbe dipendere da un maggior
carico di imagery motoria necessaria a ruotare mentalmente un arto che non è più presente a livello
fisico.
Infine si evidenzia un’attivazione nel Verme Cerebellare. Questa zona comprende quella
parte della corteccia cerebellare che riceve fibre somatosensoriali, e propriocettive provenienti dal
tronco e dagli arti; svolge una funzione di controllo nell’esecuzione dei movimenti e probabilmente,
anche in questo caso, l’attivazione può essere ricollegata ad un utilizzo dell’imagery motoria per
l’esecuzione del compito.
Dai risultati riportati sembrerebbe plausibile ipotizzare che l’attivazione di queste aree
nell’emisfero sinistro sia il risultato di un’attività mentale complessa che implicherebbe un
coinvolgimento dell’arto dominante destro utilizzato per mediare la rotazione mentale dell’arto
mancante attraverso un processo di traslazione, poiché i soggetti del gruppo sperimentale non
possono più far riferimento a quella parte del proprio corpo.
Abbiamo inoltre condotto un’analisi VBM per indagare eventuali differenze nel volume
della sostanza grigia tra pazienti amputati e controlli. I risultati hanno dimostrato nei pazienti con
amputazione dell’arto inferiore sinistro un volume maggiore di materia grigia a livello del Giro
Angolare, del Giro Frontale Medio e del Cuneo rispetto ai controlli. Nello specifico, la zona di
maggior volume localizzata nel cuneo dovrebbe corrispondere alla parte caudale del lobulo parietale
superiore PEc del macaco. Quest’area è fortemente coinvolta nella rappresentazione dell’azione e
nell’elaborazione dell’informazione riguardo la posizione degli arti nello spazio. Mostra, inoltre,
46
connessioni anatomiche con le cortecce motoria e premotoria. Quest’area sembra inoltre essere
coinvolta nella coordinazione dei movimenti degli arti nell’ambiente (Careb e coll., 2010). Bakola
et al. (2010) hanno effettuato uno studio mediante cellule marcate per individuare l’attivazione
cerebrale nei macachi durante lo svolgimento di azioni motorie degli arti. I risultati hanno suggerito
che la parte caudale del lobo parietale superiore (PEc) elabora le informazioni circa gli arti
mediante il legame che ha con altre aree coinvolte nella rappresentazione motoria, come le aree
premotorie e motorie insieme al collegamento con altre aree importanti per la navigazione spaziale
e l’analisi visiva. Questo circuito che collega il lobo parietale superiore con il lobo frontale
sembrerebbe quindi implicato nel controllo scheletro motorio atto a guidare l’azione nello spazio.
Quindi possiamo ipotizzare che in seguito all’amputazione dell’arto inferiore sinistro, avviene un
cambiamento strutturale che implica un’espansione delle zone deputate al controllo dell’azione,
ossia, il Cuneo insieme al Giro Frontale Medio (una regione associata con funzioni esecutive di alto
livello e processi decisionali) e al Giro Angolare (deputato ad elaborare la collocazione degli oggetti
nello spazio).
Probabilmente una deafferentazione a lungo termine di tipo sensorimotorio porta ad una
riduzione di materia grigia nelle aree motorie dell’arto amputato (Preißler et al., 2013) e una
riorganizzazione strutturale del cervello per far fronte alla nuova situazione. L’aumento di volume
di materia grigia nelle nostre aree è probabilmente dovuto ad un riadattamento cerebrale mediante il
maggior uso di aree quali il Giro Frontale Medio, corrispondente alla SMA, che si attiva durante
compiti di immaginazione motoria (Hanakawa, 2008). Si potrebbe ipotizzare che durante la vita
quotidiana il soggetto, non potendo far più riferimento al suo arto mancante nello schema corporeo,
utilizzi costantemente una strategia visiva che fa riferimento a una rappresentazione più astratta e
prototipica del corpo, definita da alcuni autori “descrizione strutturale del corpo” (Corradi
Dell’Acqua et al., 2009), ovvero, una “mappa visuo-spaziale” dei diversi distretti corporei.
Il Cuneo è coinvolto nell’ laborazione visiva di base e media l’attenzione, la memoria di
lavoro e l’aspettativa futura. Possiamo ipotizzare che il suo aumento sia proporzionale all’aumento
di imagery motoria ed elaborazione visiva per far fronte, nel miglior modo possibile, alle difficoltà
che comporta la mancanza di un arto, applicando strategie motorie diverse in relazione
all’orientamento dell’azione da compiere nello spazio.
Il Giro Angolare sottende diversi processi cognitivi tra cui l’organizzazione spaziale, la
memoria e l’attenzione. Uno studio condotto sulle immagini visive mentali (Boccia et al., 2015), ha
riportato l’attivazione del Giro Angolare di sinistra durante l’immaginazione di una carta
geografica; immaginare un ambiente o un oggetto implica l’attivazione di aree cerebrali coinvolte
nell’immaginazione visiva. Questo dato è utile per capire che i pazienti con amputazione utilizzano
47
maggiormente le aree deputate all’organizzazione del movimento nello spazio, attraverso la
maggior attivazione di quelle aree che richiedono il reclutamento di specifiche informazioni, quali
visione, attenzione e memoria, mediante un aumento di imagery motoria necessaria alla navigazione
nello spazio andando a compensare l’alterazione della rappresentazione del corpo determinata
dall’assenza di un arto.
In conclusione l’aumento di materia grigia a livello del Giro Angolare, del Giro Frontale
Medio e del Cuneo di sinistra sembrerebbe testimoniare una riorganizzazione strutturale dovuta ad
un’integrazione e un maggior reclutamento di informazioni sensorimotorie per orientare l’azione
dell’arto amputato nello spazio.
48
Conclusioni.
Il nostro studio, condotto su pazienti con alterazioni corporee periferiche, parte da diverse
ipotesi e interrogativi che abbiamo voluto indagare e approfondire mediante uno studio di
neuroimaging funzionale. Nonostante in letteratura, compiti di rotazione mentale siano stati già
utilizzati al fine di indagare mediante neuroimaging funzionale dove hanno sede le rappresentazioni
corporee (Schwoebel & Coslett, 2005; Corradi-Dell’Acqua et al., 2009), a nostra conoscenza nessun
lavoro di fMRI ha indagato se durante compiti di rotazione mentale di parti del corpo tra soggetti di
controllo e pazienti amputati riportata in alcuni studi comportamentali (Nico et al., 2004; Curtze et
al., 210), sia dovuta ad una modificazione della rappresentazione dello schema corporeo o
all’utilizzo di una diversa strategia per eseguire il compito che presuppone il richiamo di una diversa
rappresentazione corporea.
L’obiettivo del nostro studio è stato quello di far luce, attraverso uno studio fMRI, sui pattern
di attivazione cerebrale in pazienti con amputazione dell’arto inferiore sinistro durante un compito di
rotazione mentale di parti del corpo e di un oggetto non corporeo, comparandole con le attivazioni
dei soggetti di controllo. Ciò ha permesso di determinare come la perdita di informazioni
sensorimotorie, conseguenti all’amputazione, sia in grado di influenzare il richiamo di una
determinata strategia di rappresentazione corporea al fine di riuscire a completare il compito
sperimentale. A seguire, mediante un’analisi Voxel Based Morphometry, abbiamo indagato
eventuali differenze a livello di materia grigia confrontando il gruppo sperimentale con il gruppo di
controllo.
Rispetto al gruppo di soggetti sani, il gruppo di pazienti amputati, durante lo svolgimento
delle prove sperimentali, mostra una maggior attivazione cerebrale a livello dell’emisfero sinistro, in
particolare a livello dell’Area Motoria Supplementare, del Giro Sopramarginale, del Giro Temporale
superiore e del Verme. Queste aree sono deputate alla coordinazione e alla pianificazione dell’azione
includendo una rappresentazione somatotopica del corpo. Questi risultati ci portano ad ipotizzare che
i pazienti amputati utilizzano, come i soggetti sani, una strategia di richiamo della rappresentazione
dello schema corporeo, ma probabilmente, a causa di un mancato confronto tra immagine del corpo
presentata e controparte fisica, il cervello cerca di compensare andando a reclutare in misura
maggiore le strutture coinvolte nell’imagery motoria. Inoltre, sembrerebbe plausibile ipotizzare che
l’attivazione dell’emisfero sinistro nel gruppo sperimentale sia imputabile ad un’attività mentale
complessa che implicherebbe un coinvolgimento dell’arto dominante destro utilizzato per mediare la
rotazione mentale dell’arto mancante attraverso un processo di traslazione, processo reso necessario
49
dal fatto che i soggetti del gruppo sperimentale non possono più far riferimento a quella parte del
proprio corpo.
Anche i risultati dell’analisi VBM hanno mostrato importanti cambiamenti di volume in
alcune strutture della corteccia cerebrale. In particolare, i pazienti amputati hanno riportato un
volume maggiore di materia grigia a livello del Giro Angolare, del Giro Frontale Medio e del
Cuneo rispetto ai controlli. Sono aree deputate rispettivamente all’elaborazione della collocazione
degli oggetti nello spazio, alle funzioni esecutive di alto livello e ai processi decisionali e infine il
Cuneo, che sembrerebbe corrispondere alla parte caudale del lobulo parietale superiore (PEc) del
macaco coinvolta nella rappresentazione dell’azione e nell’elaborazione dell’informazione riguardo
la posizione degli arti nello spazio, media, nell’essere umano, l’attenzione e l’elaborazione visiva di
base. In sintesi possiamo ipotizzare che l’aumento di materia grigia in queste zone sia la causa di un
ricorso costante al sistema visivo e ad un richiamo maggiore di imagery motoria. Di conseguenza
avviene nella corteccia una riorganizzazione strutturale dovuta ad un’integrazione e un maggior
reclutamento di informazioni sensorimotorie utili per orientare l’azione dell’arto amputato nello
spazio.
Lo studio dei pazienti amputati ci ha permesso di capire quanto le rappresentazioni del corpo
siano plastiche. Questi risultati nel loro insieme possono essere il punto di partenza per elaborazioni
future più approfondite. Per ora le nostre sono solo ipotesi, poiché il gruppo di partecipanti allo
studio non è molto ampio. Con un ampliamento del campione potranno essere effettuati più
confronti sia a livello funzionale che strutturale portando a risultati più forti e concreti. Inoltre
potrebbe essere effettuata un’indagine approfondita per verificare eventuali differenze di sesso e
lateralità tra gli amputati dal momento che il nostro campione è caratterizzato da soggetti di sesso
maschile e con amputazione del solo arto sinistro. Un ulteriore approfondimento potrebbe
riguardare la distanza dall’amputazione dell’arto sui processi di compensazione cerebrale per
verificare se il cambiamento fisico improvviso porti ad una rapida riorganizzazione che rimane o
meno immutata nel tempo. Sarebbe interessante anche indagare come la presenza/assenza di una
protesi influenza la rappresentazione dello schema corporeo e approfondire e validare l’ipotesi di
come una protesi estetica o mioelettrica (Nico et al., 2004) incida in modo significativo su tale
rappresentazione.
50
BIBLIOGRAFIA.
Apkarian AV, Baliki MN, Geha PY. (2009). Towards a theory of chronic pain. Prog Neurobiol.
87:81--97.
Bakola S., Michela Gamberini M., Lauretta Passarelli L., Patrizia Fattori P. and Claudio Galletti C.
(2010). Cortical Connections of Parietal Field PEc in the Macaque: Linking Vision and Somatic
Sensation for the Control of Limb Action. Cerebral Cortex 20:2592—2604.
Bentley DE, Derbyshire SW, Youell PD, Jones AK. (2003). Caudal cingulate cortex involvement in
pain processing: an inter-individual laser evoked potential source localisation study using realistic
head models. Pain. 102:265--271.
Berlucchi G, Aglioti S (1997) The body in the brain: neural bases of corporeal awareness. Trends
Neurosci 20:560–564
Berlucchi G., Aglioti S.M. (2010). The body in the brain revisited. Exp Brain Res 200:25–35 DOI
10.1007/s00221-009-1970-7.
Birbaumer N., Lutzenberger W., Montoya P., Larbig W., Unertl K., To¨ pfner S., Grodd W., Taub
E., Flor H. (1997). Effects of regional anesthesia on phantom limb pain are mirrored in changes in
cortical reorganization. J Neurosci. 17:5503--5508.
Boccia M., Piccardi L., Palermo L., Nemmi F., Sulpizio V., Galati G. and Guariglia C. (2015). A
Penny for Your Thoughts! Patterns of Fmri Activity Reveal the Content and the Spatial Topography
of Visual Mental Images. Human Brain Mapping 36:945–958.
Bonda E., Petrides M., Frey S., and Evans A. (1995). Neural correlates of mental transformations of
the body-in-space.Proc Natl Acad Sci 92,11180–11184.
Botvinick M. & Cohen J. (1998). Rubber Hands “feel” touch that eyes see. Nature, vol. 39, pp. 756.
Boulanger-Balleyguier, G. (1967). Les &apes de la reconnaissance de soi devant le miroir [Stages
of self-recognition in front of the mirror]. Enfance, 1, 91-116.
Buxbaum, L. J., Giovannetti, T., & Libon, D. (2000). The role of the dynamic body schema in
praxis: Evidence from primary progressive apraxia. Brain and Cognition, 44, 166–191.
Buxbaum LJ, Coslett HB (2001) Specialized structural descriptions for human body parts: evidence
from autotopagnosia. Cogn Neuropsychol 18:363–381.
Calvo-Merino, B., Grezes, J., Glaser, D. E., Passingham, R. E., & Haggard, P. (2006). Seeing or
doing? Influence of visual and motor familiarity in action observation. Current Biology, 16(19),
1905–1910.
Cardoso-Leite P, Gorea A. (2010). On the perceptual/motor dissociation: a review of concepts,
theory, experimental paradigms and data interpretations. Seeing Perceiving. 23:89--151.
51
Carrie Armel K. and Ramachandran V.S. (2003). Projecting sensations to external objects: evidence
from skin conductance response. Proc Biol Sci 270(1523): 1499–1506.
Cavina-Pratesi C., Goodale M.A., Culham J.C. (2007). FMRI reveals a dissociation between
grasping and perceiving the size of real 3D objects. PLoS One. 2:e424.
Chan AW, Peelen MV, Downing PE (2004) The effect of viewpoint on body representation in the
extrastriate body area. NeuroReport 15:2407–2410.
Chen R, Corwell B., Yaseen Z., Hallett M., and Cohen L.G. (1998). Mechanisms of Cortical
Reorganization in Lower-Limb Amputees. J. Neurosci. 118, 3443–50
Cohen N.R., Cross E.S., Tunik E., Grafton S.T., Culham J.C. (2009). Ventral and dorsal stream
contributions to the online control of immediate and delayed grasping: a TMS approach.
Neuropsychologia. 47:1553--1562.
Corradi-Dell’Acqua C., Hesse M. D., Rumiati R.I. and Fink G.R. (). Where is a Nose with Respect
to a Foot? The Left Posterior Parietal Cortex Processes Spatial Relationships among Body Parts.
Cerebral Cortex 2008;18:2879—2890 doi:10.1093/cercor/bhn046.
Corradi-Dell’Acqua C., Tomasino B., and Fink G.R. (2009). What Is the Position of an Arm
Relative to the Body? Neural Correlates of Body Schema and Body Structural Description. The
Journal of Neuroscience 29(13):4162– 4171.
Coslett, H. B. (1998). Evidence for a disturbance of the body schema in neglect. Brain and
Cognition, 37, 527–544.
Coslett, H. B., Saffran, E. M., & Schwoebel, J. (2002). Knowledge of the human body: A distinct
semantic domain. Neurology, 59, 357–363.
Costantini M. & Haggard P. (2007). The rubber hand illusion: Sensitivity and reference frame for
body Ownership. Consciousness and Cognition, pp. 229–240.
Critchley Macdonald (1979). The Divine Banquet of the Brain. New York, USA: Raven Press
Books, Ltd. ISBN 0890043485.
Curtze C., Bert Otten B., Postema K. (2010). Efects of lower limb amputation on the mental
rotation of feet. Exp Brain Res 201:527–534.
Cunningham D.A., Machado A., Yue G.H., Carey J.R., Plow E.B. (2015). Functional somatotopy
revealed across multiple cortical regions using a model of complex motor task. J.Brainres 07.050.
Daurat-Hmeljiak, C., Stambak, M., & Berges, J. (1978). Il test dello schema corporeo. Una prova di
conoscenza e costruzione dell’immagine del corpo [The body schema test. A test of knowledge and
construction of body image]. Firenze, Italy: Organizzazioni Speciali.
Decety J. (1996). Do imagined and executed actions share the same neural substrate? Cognitive
Brain Research 3, 87-93.
52
De Vignemont F., Tsakiris M., Haggard P. (2005). Body mereology. In Knoblich G., Thornton
I.M., Grosjean M., Shiffrar M. The Body from inside out. New York, Oxford University Press;
pp.147-170.
De Vignemont F.(2010). Body schema and body image: Pros and cons. Neuropsychologia 48, 669–
680.
De Vignemont F. (2011). Embodiment, ownership and disownership. Consciousness and Cognition
20, 82-93.
De Lange FP, Hagoort P, Toni I (2005). Neural topography and content of movement
representations. J Cogn Neurosci 17:97–112.
Desmurget, M., Epstein, C. M., Turner, R. S., Prablanc, C., Alexander, G. E., & Grafton, S. T.
(1999). Role of the posterior parietal cortex in updating reaching movements to a visual target.
Nature Neuroscience, 2, 563–567.
Desmurget, M., & Grafton, S. T. (2000). Forward modeling allows feedback control for fast
reaching movements. Trends in Cognitive Science, 4, 423–431.
Dettmers C., Liepert J., Adler T., Rzanny R., Rijntjes M., van Schayck R., Kaiser W., Bruckner L.,
Weiller C. (1999). Abnormal motor cortex organization contralateral to early upper limb amputation
in humans. Neurosci Lett. 263:41--44.
Di Pellegrino G., Fadiga L., Fogassi L., Gallese V., Rizzolatti G. (1992). Understanding motor
events: a neurophysiological study, Exp. Brain Res, Vol. 91, pp. 176-180.
Downing P.E., Jiang Y., Shuman M., Kanwisher N. (2001). A cortical area selective for visual
processing of the human body. Science 293:2470–2473.
Draganski, B., Gaser C., Busch V., Schuierer G., Bogdahn U., May A. (2004). Neuroplasticity:
changes in grey matter induced by training. Nature. 427, 311–312.
Draganski, B., Moser T., Lummel N., Gänssbauer S., Bogdahn U., Haas F., May A. (2006).
Decrease of thalamic gray matter following limb amputation. Neuroimage. 31, 951-7.
Draganski B, May A. (2008). Training-induced structural changes in the adult human brain. Behav
Brain Res. 192:137--142.
Dominey P, Decety J, Broussolle E, Chazot G, Jeannerod M. (1995). Motor imagery of a lateralized
sequential task is asymmetrically slowed in hemi- Parkinson's patients. Neuropsychologia 33: 72741.
Elbert T., Flor H., Birbaumer N., Knecht S., Hampson S., Larbig W., Taub E. (1994). Extensive
reorganization of the somatosensory cortex in adult humans after nervous system injury.
Neuroreport. 5:2593--2597.
53
Ellison A, Cowey A. (2006). TMS can reveal contrasting functions of the dorsal and ventral visual
processing streams. Exp Brain Res. 175:618--625.
Ellison A, Cowey A. (2007). Time course of the involvement of the ventral and dorsal visual
processing streams in a visuospatial task. Neuropsychologia. 45:3335--3339.
Ellison A, Cowey A. (2009). Differential and co-involvement of areas of the temporal and parietal
streams in visual tasks. Neuropsychologia. 47:1609--1614.
Farrer C, Frith CD (2002) Gallagher S. (1986). Body image and Body schema: a conceptual
clarification. The journal of mind and behavior 541-554 ISSN 0271-0137.
Flor H., Elbert T., Knecht S., Wienbruch C., Pantev C., Birbaumer N., Larbig W., Taub E. (1995).
Phantom-limb pain as a perceptual correlate of cortical reorganization following arm amputation.
Nature. 375:482--484.
Gallagher, S. (2005). How the Body Shapes the Mind. Review of psyche 284, 0199271941.
Gallese, V., Fadiga, L., Fogassi, L., & Rizzolatti, G. (1996). Action recognition in the premotor
cortex. Brain, 119, 593–609.
Gaser C., Nenadic I., Weiss T., Miltner W.H.R., Sauer H. 2004. Gray matter volume loss after
upper limb amputation. Klinische Neurophysiologie. 35:144.
Gazzola, V., van der Worp, H., Mulder, T., Wicker, B., Rizzolatti, G., & Keysers, C. (2007).
Aplasics born without hands mirror the goal of hand actions with their feet. Current Biology,
17(14), 1235–1240.
Gentilucci, M., Daprati, E., & Gangitano, M. (1998). Right-handers and left-handers have different
representations of their own hand. Brain Research. Cognitive Brain Research, 6(3), 185–192.
Giummarra MJ, Gibson SJ, Georgiou-Karistianis N, Bradshaw JL. 2007. Central mechanisms in
phantom limb perception: the past, present and future. Brain Res Rev. 54:219-231.
Goodale, M. A., & Milner, A. D. (1992). Separate visual pathways for perception and action.
Trends in Neuroscience, 15, 20–25.
Hanakawa T., Dimyan M.A., Hallett M. (2008). Motor planning, imagery, and execution in the
distributed motor network: a time-course study with functional MRI. Cereb Cortex 18(12):2775-88.
Head H., Holmes G. Sensory disturbances from cerebral lesions. Brain,1911-1912, 34:102-254.
Ionta, S., Fourkas, A. D., Fiorio, M., & Aglioti, S. M. (2007). The influence of hands posture on
mental rotation of hands and feet. Experimental Brain Research, 183(1), 1–7.
Johansson G. (1973). Visual perception of biological motion and a model for its analysis. Percept
Psychophy 14: 201–11.
Jones EG. (2000). Cortical and subcortical contributions to activity-dependent plasticity in primate
somatosensory cortex. Annu. Rev. Neurosci. 23, 1–37.
54
Judica A., Zoccolotti P., Pizzamiglio L. (2006). La riabilitazione neuropsicologica. Premesse
teoriche e applicazioni cliniche. Milano: Mazzucchi, 2006.
Katz J., Melzack R. (1990). Pain memories in phantom limbs—review and clinical observations.
Pain. 43:319--336.
Kosslyn S.M., Ganis G., & Thompson W. (2001). Neural foundations of imagery. Nature Reviews
Neuroscience, 2, 635-642.
Lackner, J.R. (1988). Some proprioceptive influences on the perceptual representation of body
shape and orientation. Brain 111, 281-297].
Langer N., Ha¨nggi J., Mu¨ller N.A., Simmen H.P., Ja¨ncke L. (2012). Effects of limb
immobilization on brain plasticity. Neurology 78(3):182-8. doi: 10.1212/WNL.0b013e31823fcd9c
Le Boulch J. (1966). Educare con il movimento. Roma: Armando editore, 1966.
Le Boulch J. (2006). Movimento e sviluppo della persona. Roma: Musicalificio Grande Blu, 2006.
Lis A., Venuti P., Basile A. P., Finesso R. (1988), Lo sviluppo dello schema corporeo. Età
Evolutiva, 30, 23-34.
G. Moseley G., Olthof N., Venema A., Don S., Wijers M., Gallace A., and Spence C. (2008).
Psychologically induced cooling of a specific body part caused by the illusory ownership of an
artificial counterpart. PNAS 2008 105 (35) 13169-13173.
Marjolein P., Kammers M., Mulder J., De Vignemont F. and Dijkerman H. C. (2010). The weight
of representing the body: addressing the potentially indefinite number of body representations in
healthy individuals Exp Brain Res. 204(3): 333–342.
May A. (2008). Chronic pain may change the structure of the brain. Pain. 137:7--15.
May A. (2009). [Chronic pain alters the structure of the brain]. Schmerz. 23:569--575.
May A. (2011). Structural brain imaging: a Window into chronic pain. Neuroscientist. 17:209—
220.
Medina J., Coslett H.B. (2010). From maps to form to space: touch and the body schema.
Neuropsychologia, n.48(3); pp.645-654.
Meltzoff AN, Moore MK (1977) Imitation of facial and manual gestures by human neonates.
Science 198:75–78.
Melzack R. (1990). Phantom limbs and the concept of a neuromatrix. Trends Neurosci. 13:88--92.
Melzack R. (2005). Evolution of the neuromatrix theory of pain. The Prithvi Raj Lecture: presented
at the third World Congress of World Institute of Pain, Barcelona 2004. Pain Pract. 5:85--94.
Merleau-Ponty, M. (1945). Phénoménologie de la perception. Paris: Gallimard.
Milner D, Goodale MA (1995). The visual brain in action. Oxford University Press, Oxford.
55
Mishkin, M., Ungerleider, L. G., & Macko, K. A. (1983). Object vision and spatial vision: Two
cortical pathways. Trends in Neuroscience, 6, 414–417.
Molinari E., Riva G. (2004). Psicologia Clinica dell’Obesità: Ricerche e Interventi. Torino: Bollati
Boringhieri, 2004.
Morris J.P., Pelphrey K.A., McCarthy G. (2006). Occipitotemporal activation evoked by the
perception of human bodies is modulated by the presence or absence of the face. Neuropsychologia
44:1919–1927
Nico D., Daprati E.,Rigal F.,Parsons L. and Angela Sirigu A. (2004). Left and right hand
recognition in upper limb amputees. Brain 127, 120-132 DOI: 10.1093/brain/awh006.
Paillard, J. (1987) Cognitive versus sensori-motor encoding of spatial information. In: P. Ellen, C.
Thinus-Blanc (Eds), Cognitive processes and spatial orientation in animal and man. Vol. Il.
Neurophysiology and developmental aspects. Dordrecht: Martinus Nijhoff, 43-77.
Paillard J (1999) Body schema and body image—a double dissociation in deafferented patients. In:
Gantchev GN, Mori S, Massion J (eds) Motor control, today and tomorrow. Academic Publishing
House, Sophia.
Palermo L., Di Vita A., Piccardi L., Traballesi M., and Guariglia C. (2014). Bottom-Up and TopDown Processes in Body Representation: A Study of Brain-Damaged and Amputee Patients.
Neuropsychology Online Publication http://dx.doi.org/10.1037/neu0000086.
Parsons, L. M. (1987). Imagined spatial transformation of one’s body. Journal of Experimental
Psychology, 116, 172–191.
Parsons LM (1994) Temporal and kinematic properties of motor behavior reXected in mentally
simulated action. J Exp Psychol Hum Percept Perform 20:709–730.
Parsons L.M., Fox P.T., Downs H.J., Glass T., Hirsch T.B., Martin C.C., Jerabek P.A. & Lancaster
J.L. (1995). Use of implicit motor imagery for visual shape discrimination as revealed by PET.
Nature 375, 54–58.
Pascual-Leone A., Cammarota A., Wassermann E.M., Brasil-Neto J.P., Cohen L.G., Hallett M.
(1993). Modulation of motor cortical outputs to the reading hand of Braille readers. Ann Neurol.
34:33--37.
Pascual-Leone A., Nguyet D, Cohen LG, Brasil-Neto JP, Cammarota A, Hallett M. (1995).
Modulation of muscle responses evoked by transcranial magnetic stimulation during the acquisition
of new fine motor skills. J. Neurophysiol., 74, 1037–45.
Pavani, F., Spence, C., & Driver, J. (2000). Visual capture of touch: Out-of-the-body experiences
with rubber gloves. Psychological Science, 11(5), 353–359.
Peyron R., Laurent B., Garcia-Larrea L. (2000). Functional imaging of brain responses to pain. A
review and meta-analysis. Neurophysiol Clin. 30:263--288.
56
Pick A (1922) Störungen der Orientierung am eigenen Körper. Psychol Forsch 2:303–318
Plügge H. (1967). Der Mensch und sein Leib. University of Michigan: Niemeyer.
Preißler S, Dietrich C, Meissner W, Huonker R, Hofmann GO, Miltner WH, Weiss T. (2011).
Brachial plexus block in phantom limb pain: a case report. Pain Med. 12:1649--1654.
Preißler1 S.., Feiler J., Dietrich C., Hofmann G.O.,. Miltner W.H.R. and Weiss T. (2013).Gray
Matter Changes Following Limb Amputation with High and Low Intensities of Phantom Limb
Pain. Cerebral Cortex 23:1038-1048
Rizzolatti, G. and Craighero, L. (2004) The mirror neuron system. Ann. Rev. Neurosci. 27: 169192.
Ramachandran V.S., Ruskin D., Cobb S., Rogers-Ramachandran D., Tyler C.W. (1994). On the
Perception of Illusory Contours. Vision Res. Vol. 34, No. 23, pp. 3145-3152.
Ramachandran VS, Rogers-Ramachandran D. (1966). Synaesthesia in phantom limbs induced with
mirrors. Proc Biol Sci. 22;263(1369):377-86.
Ramachandran V.S. e Rogers-Ramachandran D. (2000). Phantom limbs and neural plasticity. Arch
Neurol 57: 317–20.
Ramachandran VS, Rogers-Ramachandran D. 2006. Touching illusions. Sci Am Mind. 17:18--20.
Rice N.J., Valyear K.F., Goodale M.A., Milner A.D., Culham J.C. 2007. Orientation sensitivity to
graspable objects: an fMRI adaptation study. Neuroimage. 36:T87--T93.
Rossetti Y., Pisella L., Vighetto A. (2003). Optic ataxia revisited: visually guided action versus
immediate visuomotor control. Exp Brain Res. 153:171--179.
Saxe R., Jamal N. and Powell L. (2005). My Body or Yours? The Effect of Visual Perspective on
Cortical Body Representations. Cerebral Cortex doi:10.1093/cercor/bhi095
Saxe R., Jamal N., Powell L. (2006). My body or yours? The effect of visual perspective on cortical
body representations. Cereb Cortex 16:178–182
Schwoebel J, Friedman R, Duda N, Coslett HB (2001) Pain and the body schema: evidence for
peripheral eVects on mental representations of movement. Brain 124:2098–2104
Schwoebel1 J.and Coslett H.B (2005). Evidence for Multiple, Distinct Representations
of the Human Body. Journal of Cognitive Neuroscience 17:4, pp. 543–553.
Sergent, J. (1982). The cerebral balance of power: Confrontation or cooperation? Journal of
Experimental Psychology: Human Perception and Performance, 8, 253–272.
Shilder p. (1935). Immagine di sé e schema corporeo. Tr. It. (1990) Milano: Franco Angeli.
Sirigu A, Grafman J, Bressler K (1991) Multiple representations contribute to body knowledge
processing: evidence from a case of autotopagnosia. Brain 114:629–642.
57
Sirigu A., Cohen L., Duhamel J.R., Pillon B., Dubois B., Agid Y., et al. (1995) Congruent unilateral
impairments for real and imagined hand movements. Neuroreport 6: 997-1001.
Sirigu A., Duhamel J.R., Cohen L., Pillon B., Dubois B., Agid Y. (1996). The mental
representation of hand movements after parietal cortex damage. Science 273: 1564-8.
Slaughter, V., & Heron, M. (2004). Origins and early development of human body knowledge.
Monographs of the Society for Research in Child Development, 69, vii, 1–102. doi:10.1111/j.0037976X.2004 .00287.x
Snyder, L. H., Grieve, K. L., Brotchie, P., & Andersen, R. A. (1998). Separate body and worldreferenced representations of visual space in parietal cortex. Nature, 394, 887–891.
Suzuki, K., Yamadori, A., & Fujii, T. (1997). Category-specific comprehension deficit restricted to
body parts. Neurocase, 3, 193–200.
Taylor J.C., Wiggett A.J., Downing P.E. (2007). Functional MRI analysis of body and body part
representations in the extrastriate and fusiform body areas. J Neurophysiol 98:1626–1633.
Tessari A, Tsakiris M, Borghi AM, Serino A. (2010). The sense of body: A multidisciplinary
approach to body representation. Neuropsychologia 48(3):643-4.
Tomasino B. & Rumiati R.I. (2004). Effects of strategies on mental rotation and hemispheric
lateralization: neuropsychological evidence. Journal of Cognitive Neuroscience, 5, 878-888.
Treede R.D., Kenshalo D.R., Gracely R.H., Jones AKP. (1999). The cortical representation of pain.
Pain. 79:105--111.
Ungerleider LG, Mishkin M (1982) Two cortical visual systems. In: Ingle DJ, Goodale MA,
Mansfield RJW (eds) Analysis of visual behavior. The MIT Press, Cambridge, MA, pp 549–586
Urgesi C., Berlucchi G., Aglioti S.M. (2004). Magnetic stimulation of extrastriate body area impairs
visual processing of nonfacial body parts. Curr Biol 14:2130–2134.
Urgesi, C., Calvo-Merino, B., Haggard, P., & Aglioti, S. M. (2007). Transcranial magnetic
stimulation reveals two cortical pathways for visual body processing. The Journal of Neuroscience,
27, 8023–8030.
Vallar G., Gilles Rode G. (2009). Commentary on Bonnier P. L’aschématie. Rev Neurol (Paris)
1905;13:605–9. Journal homepage: www.elsevier.com/locate/yebeh. Epilepsy & Behavior 16, 397–
400.
Van Elk M., Forget J., Blanke O. (2013). The effect of limb crossing and limb congruency on
multisensory integration in peripersonal space for the upper and lower extremities. Journal
homepage: www.elsevier.com/locate/concog.
58
Weiss T., Miltner W.H.R., Liepert J., Meissner W., Taub E. (2004). Rapid functional plasticity in
the primary somatomotor cortex and perceptual changes after nerve block. Eur J Neurosci.
20:3413--3423.
Wolpert, D. M., & Ghahramani, Z. (2000). Computational principles of movement neuroscience.
Nature Neuroscience, 3, 1212–1217.
Wolpert, D. M., Ghahramani, Z., & Jordan, M. I. (1995). An internal model for sensorimotor
integration. Science, 269, 1880–1882.
Zopf, R., Savage, G., & Williams, M. A. (2010). Crossmodal congruency measures of lateral
distance effects on the rubber hand illusion. Neuropsychologia, 48(3), 713–725.
59
Ringraziamenti
Un primo ringraziamento va alla Prof.ssa Cecilia Guariglia che mi ha dato la possibilità di
prendere parte ad un progetto in cui ho avuto una prima esperienza diretta con metodi e materiali al
fine di indagare alcuni aspetti per la ricerca neuropsicologica. Questo lavoro mi ha trasmesso
entusiasmo e voglia di approfondire le mie conoscenze. Grazie alla Dott.ssa Antonella Di Vita e alla
Dott.ssa Maddalena Boccia, due persone tenaci e appassionate che con la loro pazienza mi hanno
seguito durante questo percorso insegnandomi nuove cose e dandomi la possibilità di cimentarmi in
prima persona.
Grazie alla mia famiglia che mi è sempre stata vicina dal primo giorno in cui ho fatto il mio
ingresso all’università. Mi hanno spronata ad andare avanti e a non mollare mai. Specialmente nei
miei “soliti momenti di crisi” mi hanno aiutato ad essere forte e ad uscirne a testa alta.
Grazie al mio fidanzato, una persona speciale che è sempre stata al mio fianco. Se sono
riuscita ad affrontare le mie difficoltà, i miei blocchi e le mie paure è proprio grazie a lui che ha
sempre avuto fiducia in me e ha saputo indicarmi la giusta strada.
Grazie ai miei amici che hanno reso più bello il mio percorso universitario! Domenico, Irene,
Alessandro, Filippo e Pier, persone folli e fantastiche con cui ho condiviso giornate di studio intenso,
giornate spensierate e serate bellissime!
Grazie a Stefania, amica di sempre, che nonostante i chilometri di distanza mi è sempre stata
vicina con la sua enorme fiducia nei miei confronti e le sue strigliate per non farmi mollare.
Un piccolo grazie va anche a me stessa che ho cercato di dare il massimo accompagnata dal
mio motto “Renderò possibile l’impossibile”.