Il fusto
Il fusto funzioni e struttura
Nelle cormofite, la presenza di tessuti
conduttori e di sostegno, ha reso
possibile lo sviluppo di un organo
epigeo, il fusto o caule che ,
assolve a numerose funzioni:
1. Sostiene le porzioni fotosintetizzanti,
rappresentate in gran parte dalle
foglie e le espone alla luce nella
giusta angolazione;
2. Garantisce il trasporto delle sostanze
nutritive dalle radici e dalle foglie a
tutte le cellule della pianta;
3. Costituisce un organo di riserva,
seppure in misura molto minore
rispetto alle radici.
Osservando l’estremità del fusto è possibile mettere
in evidenza tre zone che dall’apice, verso il basso
sono dette rispettivamente:
1 Zona di accrescimento per divisione
Le cellule del meristema apicale, sono in continua divisione,
vi è produzione di nuovi elementi che allontanandosi dal
meristema, cessano di dividersi e vanno incontro ad un
processo di distensione e differenziazione.
2 Zona di accrescimento per distensione e di
differenziazione,
È la zona in cui avviene l’allungamento del giovane fusto
3 Zona di struttura primaria
Dove le cellule sono ormai morfologicamente e
funzionalmente adulte.
L’accrescimento primario in lunghezza è a questo punto
terminato.
Il meristema apicale del fusto è pertanto responsabile dell’ accrescimento
primario della pianta, e cioè della crescita in lunghezza delle porzioni aeree.
In alcune piante (es.: graminacee), sono presenti alla base degli internodi, i
meristemi intercalari, che determinano un ulteriore allungamento del fusto.
In tutti i fusti, inoltre, nelle zone di differenziazione, rimangono delle aree
meristematiche residue da cui, in corrispondenza delle bozze fogliari, si
originano i fasci conduttori delle foglie.
Nelle piante che presentano solo un accrescimento primario,
(pteridofite, quasi tutte le monocotiledoni ed alcune
dicotiledoni), la struttura primaria è quella definitiva.
Nelle gimnosperme e nella maggior parte delle dicotiledoni
invece, i tessuti primari del corpo della pianta, sono in
gran parte sostituiti, durante
l’accrescimento secondario in spessore,
dai tessuti secondari.
Alla zona di struttura primaria, segue dunque una
zona di struttura secondaria.
È da tener presente che non esiste una netta demarcazione tra
la varie zone, ma il passaggio avviene gradatamente.
Si ricorda ad esempio che già nella zona di accrescimento
per distensione esistono e sono funzionanti il protoxilema
ed il protofloema, costituiti da cellule adulte.
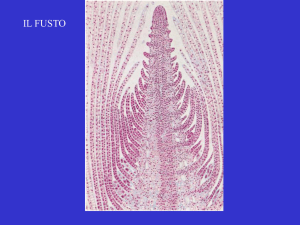
Struttura primaria del fusto
Effettuando una sezione trasversale di un
giovane fusto di dicotiledone ad una
certa distanza dall’apice (a livello della
struttura primaria), è possibile osservare
al microscopio ottico, dall’esterno verso
l’interno
• Epidermide
• Corteccia
• Cilindro centrale.
Phalangium
esculentum Nutt.
= Camassia quamash
Fusto
monocotiledone mais
DicotiledonePhaseolus vulgaris
La corteccia
La corteccia è formata da più di un tipo di tessuto.
Negli strati più superficiali, nei quali la radiazione
solare penetra facilmente, è presente un parenchima
clorofilliano; più all’interno, dove la radiazione
solare non arriva, i plastidi si saranno differenziati
in leucoplasti e il parenchima che li contiene sarà
essenzialmente un parenchima di riserva.
Nella corteccia trovano la collocazione più adeguata
anche i tessuti meccanici.
Le sollecitazioni cui va incontro un fusto sono infatti
tipicamente sforzi di flessione, che si traducono in
uno sforzo di compressione su un lato e in uno di
trazione su quello opposto.
Le sollecitazioni saranno quindi massime in periferia e
nulle nella zona centrale.
E’ nella zona periferica che andranno quindi collocate
le “ zone resistenti”, costituite dai collenchimi e/o
sclerenchimi
(nella radice, i tessuti meccanici, sono maggiormente
dislocati al centro dell’organo, in quanto essa è
sottoposta a sforzi di trazione più che di
piegamento e torsione).
C
Lo spessore della corteccia è in genere
limitato, presenta solitamente una zona
sottoepidermica collenchimatica alla quale
segue una zona più o meno spessa di
cellule parenchimatiche nel cui spessore si
possono trovare fibre nonché tessuti
segregatori di vario tipo (Canali
resiniferi,Tubi laticiferi ecc.).
Se si osserva ad esempio al microscopio la
sezione trasversale di un fusto di sedano,
le cellule collenchimatiche, formano dei
cordoni longitudinali ai vertici di fusti
erbacei a sezione quadrangolare .
Mentre i tessuti meccanici, danno
resistenza e flessibilità al fusticino, le
cellule parenchimatiche, svolgono sia
una funzione fotosintetica (più
sviluppata nelle cellule sottostanti
l’epidermide) che di riserva.
Lo strato più interno della corteccia è in
genere costituito da cellule più
compatte, con prevalente funzione di
riserva.
Nei fusti sotterranei o nei fusti sommersi
di piante acquatiche assume l’aspetto
di un vero e proprio tessuto
tegumentale endodermico.
Cinnamomum zeylanicum
Sezione trasversale di
fusto di Nimphaea
sp.ricco di
parenchima
aerifero.
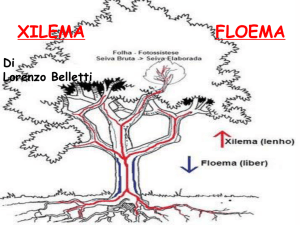
Cilindro centrale o stele
Il cilindro centrale o stele, di una pianta, è la zona più interna
del fusto o della radice in cui si trovano i tessuti conduttori,
che nella struttura primaria, sono riuniti in cordoni verticali
detti fasci.
Un fascio può essere definito come un insieme di tessuti
adibiti sia al trasporto di sostanze organiche sia di acqua e
sali minerali .
I fasci xilematici e floematici sono in stretta relazione ma, a
seconda della posizione occupata nello spazio possono
essere suddivisi in diverse categorie.
1.
Fasci concentrici
Sono divisi in:
a.
Fascio perixilematico, in cui lo xilema circonda il floema; è presente in
certi rizomi e nei fusti di alcune monocotiledoni;
b.
Fascio perifloematico, in cui il floema circonda lo xilema; è presente
nella maggior parte delle felci.
il perifloematico, è stato progressivamente modificato nel corso dell’evoluzione
delle piante terrestri e deve essere cosiderato l’antenato dei moderni fasci,
quelli collaterali.
È probabile che il successo in ambiente terrestre del tipo perifloematico sia
dovuto a:
1)
La funzione svolta dallo xilema è prioritaria rispetto a quella del floema ed
il fascio perifloematico, assicura maggiore protezione allo xilema; una
ferita, anche profonda, può danneggiare al massimo un elemento cribroso
ma difficilmente un più prezioso vaso;
2)
Lo xilema quando non è più funzionante, è più utile del vecchio floema,
potendo servire anche come elemento di sostegno; è vantaggioso quindi
avere uno xilema interno, che si conserva anche con l’eliminazione dei
tessuti esterni.
Uno svantaggio dei fasci concentrici è però che, essendo sempre privi di tessuto meristematico, non
consentono un accrescimento diametrico secondario: il sistema concentrico non è vincente se
persiste come tale!
La frammentazione del grande anello di xilema e floema e in alcuni casi, l’interposizione cambiale
nelle “isole” così formatesi sono stati i processi evolutivi che hanno portato alla presenza dei
fasci collaterali e bicollaterali delle spermatofite.
Fasci collaterali
Lo xilema è situato sullo stesso raggio del
floema, internamente a questo.
Se tra i due tessuti conduttori è interposta una
zona meristematica primaria (il futuro
cambio intrafascicolare) il fascio collaterale
è aperto (gimnosperme e dicotiledoni)(C)
Quando invece xilema e floema sono a stretto
contatto senza strati meristematici
intercalati, il fascio collaterale è chiuso (D )
È tipico delle piante che non presentano
accrescimento secondario in spessore
(monocotiledoni e molte piante erbacee) ma
è anche presente in molte nervature fogliari.
Fascio bicollaterale
È un fascio APERTO caratterizzato dalla presenza di
una zona floematica interna, oltre a quella esterna
normale.
Lo xilema, quindi, risulta sempre separato dal floema
per mezzo di due zone meristematiche
(Solanaceae, Cucurbitaceae, Apocinaceae (E))
Fascio radiale
I cordoni xilematici e floematici, in numero
variabile, sono posti su raggi diversi, cioè
affiancati lateralmente in modo alternato.
Questi sono denominati come arche (F).
La differenziazione degli elementi vasali
avviene in modo centripeto dalla periferia
verso il centro, portando, a volte, alla
formazione di una massa legnosa a forma di
stella. È tipico delle radici delle cormofite.
Teoria stelare
Le piante vascolari hanno organizzato e
disposto i vari tipi di fascio all’interno del
cilindro centrale per ottenere, nel corso
dell’evoluzione, la soluzione migliore per
utilizzare lo spazio e quindi la luce.
Per colonizzare la terra si sono superati degli
ostacoli specializzando e sviluppando
determinati tessuti per la conduzione(già
nelle briofite sono presenti cellule come
gli idroidi ed i leptoidi).
I fasci conduttori compaiono a livello delle
pteridofite.
La presenza di tessuti conduttori lignificati
(tracheidi) ha permesso di incrementare il
sostegno del fusto ed il trasporto a
distanza dell’acqua il che, ha consentito a
differenza delle briofite uno sviluppo in
altezza e diventare addirittura arboree.
I tessuti conduttori si trovano nel cilindro
centrale (STELE).
La struttura più primitiva tipica di molte pteridofite è
la protostele (A), nella quale è presente, al centro
del fusto, un unico fascio perifloematico.
Il passo evolutivo successivo ha permesso di sfruttare
meglio lo spazio ed ha posto al riparo da problemi
derivanti da possibili danneggiamenti dell’unico
fascio presente nella protostele.
Dopo una divisione dicotomica dalla stele centrale si
dipartono più fasci che seguono i germogli laterali
e si ottiene una actinostele (B), fascio radiale,
presente nella maggior parte delle radici e nei fusti
di alcune licofite.
Un migliore utilizzo dello spazio e della luce ha
richiesto uno sviluppo in altezza con conseguenti
problemi di staticità.
Nella sifonostele (C) gli elementi xilematici e
floematici vengono utilizzati sia per la conduzione
che per garantire la staticità e la resistenza alla
flessione del fusto, che può così aumentare in
altezza.
È come un tubo interno al fusto perché è caratterizzato
da un anello continuo di fasci collaterali o
bicollaterali (Selaginella) che ha al centro un
tessuto parenchimatico di riserva che costituisce il
midollo. È presente nei fusti di molte felci e di
alcune gimnosperme e angiosperme.
Una semplice sifonostele non assicura però una buona ramificazione
necessaria alla formazione di germogli laterali.
Il problema viene superato nella sifono-eustele (presente in alcune felci)
(D) dove si aveva un’iniziale frammentazione dell’anello di tessuto
conduttore, ma ancor meglio nell’eustele (E).
Presente in Equisetum, nella generalità delle gimnosperme ed in molte
dicotiledoni, si ha una disposizione circolare di più fasci collaterali
intervallati da raggi midollari, che permettono il collegamento tra
midollo e corteccia. L’ambiente terrestre è stato conquistato da
questa struttura.
Infine parliamo della atactostele delle monocotiledoni(F) derivante
dalla eustele in quanto in alcune specie di dicotiledoni sono presenti
steli intermedie alle due forme.
Nell’atactostele i fasci sono sempre di tipo collaterale chiuso e dispersi
nel tessuto fondamentale, tanto che non è possibile distinguere il
midollo dalla corteccia.
Sono più grandi verso l’interno , piccoli è più numerosi verso l’esterno.
La presenza di un gran numero di fasci circondati da una
guaina di fibre e la loro disposizione caratteristica, dà alle
monocotiledoni una rigidità affiancata all’elasticità, tali
che, sebbene prive di accrescimento secondario, fa sì che
molte di esse possano svilupparsi in misura maggiore
rispetto alle dicotiledoni erbacee.
Inoltre, viene perfezionato il trasporto idrico in quanto le
foglie sono innervate direttamente dai fasci collaterali
autonomi, che decorrono longitudinalmente nel fusto.
Questo sistema di conduzione in cui ogni fascio è collegato ad
una singola foglia, favorisce notevolmente l’attività
fotosintetica.
DicotiledonePhaseolus vulgaris
Fusto
monocotiledone mais
Cilindro centrale o stele
È posto in posizione centrale,
occupa gran parte del fusto
ed in esso, si distinguono i
fasci conduttori ed una zona
centrale di parenchima
midollare con funzione di
riserva.
La distribuzione dei fasci nelle
dicotiledoni è variabile, ma nelle
regioni internodali, è
generalmente riconducibile
alla sifonostele o alla
eustele.
Sezione trasversale del giovane fusto di tiglio (Tilia Americana)
che presenta una tipica sifonostele-nel particolare (b) si mette in
evidenza la presenza di collenchima sottoepidemico, dei dotti
mucipari e dei fasci conduttori primari.
Fusto di Aristolochia sp. (Dicotiledoni) in
sezione trasversale. Esempio di eustele
1.
2.
3.
4.
5.
6.
7.
8.
9.
Altro esempio di struttura eustelica: fusto di
Clematis vitalba (Dicotiledoni) in sezione
trasversale.
Libro
Legno
Epidermide
Corteccia
Periciclo
Endoderma
Zona perimidollare
Raggi midollari
Midollo centrale
A livello dei nodi, tuttavia, la struttura anatomica si complica, in quanto alcuni
fasci conduttori della stele si dirigono verso le appendici caulinari,
lasciando in molti casi nella stele una zona più o meno ampia priva di
tessuto conduttore, la lacuna fogliare.
L’insieme dei fasci che, attraversano la corteccia, si dirigono verso l’appendice
laterale è detto traccia fogliare.
Le lacune fogliari, evidenti nella sifonostele sono difficilmente individuabili
nella eustele, in cui si confondono con i raggi midollari.
Nelle
MONOCOTILEDONI
invece, i fasci di tipo
collaterale chiuso, sono
uniformemente sparsi nel
tessuto fondamentale,
costituendo una
atactostele in cui non è
visibile la distinzione,
tipica delle dicotiledoni,
tra corteccia e midollo.
plectostele
Quindi ci sono piante che hanno un unico cilindro
centrale nel fusto (o stele) e parleremo di fusti
monostelici ed altre il cui fusto possiede più
cilindri centrali ed in questo caso si parla fusti
polistelici.
SIFONOSTELE (o solenostele, siphon e solen dal
greco, tubo) la struttura del fascio è tubulosa
l’unica stele, è caratterizzata da un unico fascio
concentrico perifloematico con midollo al
centro.
Se al nome viene fatto seguire da un aggettivo
qualificativo,
ECTOFLOICA, si precisa che il floema si trova sul
solo lato esterno del legno;
ANFIFLOICA è caratterizzata da un unico fascio
presentante midollo al centro e una porzione
xilematica ad anello racchiusa da due cerchi
floematici uno all'esterno ed uno all'interno.
APLOSTELE o PROTOSTELE
cauli monostelici, si possono avere un unico fascio
concentrico perifloematico; il legno è
compatto cosicché non esiste un midollo
ACTINOSTELE
l’unica stele è costituita da un unico fascio raggiato
PLECTOSTELE
l’unica stele è costituita da strati di legno e di libro
sifonostele
anfifloica
Dennstaedtia punctilobula
(Filicales) in sezione
trasversale.
La stele è caratterizzata
da un unico fascio
presentante midollo
al centro e una
porzione xilematica
ad anello racchiusa da
due cerchi floematici
uno all'esterno ed
uno all'interno
Fusto di Botrychium virginianum (Ophioglossales) in sezione trasversale sopra
un nodo.
A livello dei nodi, la stele è
caratterizzata da un unico
fascio concentrico
perifloematico con
midollo al centro:
sifonostele ectofloica.
Particolare della stele
Fusto di Piper geniculatum (Dicotiledoni) in sezione
trasversale.
Esempio di una struttura
particolare di
Dicotiledone in cui i
fasci sono dislocati su
due cerchi:
- quello esterno
formante un anello
continuo con possibilità
di accrescimento
secondario,
- quello interno con
disposizione più
irregolare che ricorda la
struttura atactostelica delle
Monocotiledoni.
eustele di stelo di Lupinus arboreus
A differenza delle Dicotiledoni, nelle
Monocotiledoni i fasci sono disposti
"disordinatamente" nel cilindro
centrale (struttura atactostelica).
Essi sono di tipo collaterale chiuso, cioè
senza cambio interposto tra xilema e
floema.
Tali fasci sono immersi in un parenchima
con cellule a pareti lignificate,
l'ispessimento delle pareti è via via
maggiore dall'interno verso l'esterno.
Nella corteccia si nota abbastanza bene
l'endodermide e quindi, verso
l'esterno, un parenchima
assimilatore che lascia il posto a un
collenchima in posizione
sottoepidermica.
L'epidermide mostra una spessa cuticola.
atactostele di stelo di Zea mays.
Fusto di Zea mays (Monocotiledoni) in sezione trasversale.
Altro esempio di
struttura
atactostelica, con i
fasci cribro-vascolari
che si spingono a
ridosso
dell’epidermide
Nelle monocotiledoni e nelle dicotiledoni erbacee, nonostante ci
sia solo accrescimento primario, cioè vi è crescita dovuta ai
soli meristemi primari, si ha un limitato accrescimento in
spessore dovuto essenzialmente all’ingrossamento delle
cellule dei vari tessuti definitivi.
Solo in rari casi si ha un ragguardevole aumento delle
dimensioni del fusto grazie a particolari modalità di
accrescimento primario in spessore osservabile nelle palme
tra le monocotiledoni, e nei cactus e nei tuberi di patata tra le
dicotiledoni.
A parte queste eccezioni, di solito l’accrescimento in spessore è
di tipo secondario ed inizia solo al termine di quello primario,
protraendosi fino alla morte della pianta.
Le cotiledoni con fusti con accrescimento secondario
Monocotiledoni arborescenti come la palma da
cocco(Cocos sp.) e l’albero di Joshua (Yucca
brevifolia) possono raggiungere dimensioni
ragguardevoli. In realtà esistono 3 tipi di
monocotiledoni arborescenti:
1.
Le palme,come il cocco, che non si
ramificano e non hanno un vero
accrescimento secondario:
2.
I pandàni (Pandanus sp.) che esibiscono
fusti ramificati, ma sono anch’essi privi di
accrescimento laterale
3.
Le liliacee arboree (come Yucca)e altre che
portano rami, possiedono un cambio e
dunque hanno un vero accrescimento
secondario.
I fusti delle palme possiedono un gran numero di
fasci di conduzione, molti dei quali sono
tracce fogliari. Le foglie sono grandi e la
loro base abbraccia il fusto; a ogni foglia
afferiscono molte tracce fogliari.
a
b
c
Alla base delle palme avviene un certo
ispessimento che però deriva da radici
avventizie basali.
Un ingrossamento del fusto nella sua
interezza in effetti avviene, ed è opera
della divisione e dell’espansione delle
cellule parenchimatiche responsabili del
cosiddetto accrescimento secondario
diffuso. Non è un vero e proprio
accrescimento secondario, perché non è
opera di un cambio, che è assente.
Diverse monocotiledoni, incluse Agave e
Dracaena(albero del sangue di drago),
mostrano invece un accrescimento
secondario vero e proprio, derivante
dall’azione di un cambio; i loro fusti sono
affusolati, più sottili in alto e più spessi
alla base.
Questa architettura deriva dalla disposizione
spaziale delle cellule prodotte dal
cambio.
Il cambio delle monocotiledoni è il solo che
produca la maggior parte delle cellule
parenchimatiche con i fasci conduttori
immersi in questo tessuto fondamentale.
I fasci vascolari secondari differiscono dai
primi perché in essi, lo xilema circonda il
floema.
Nelle monocotiledoni, qualche fusto più
vecchio mostra uno strato simile al
sughero che rimpiazza l’epidermide.
Questo strato, detto sughero stratificato,
non è considerato appartenente al
periderma perché non deriva dall’attività
di un cambio subero fellodermico(CHE
NON C’E’), ma dalla proliferazione delle
cellule parenchimatiche della periferia
del fusto.
Struttura secondaria del fusto
In alcune pteridofite oggi estinte, era presente un accrescimento
secondario in spessore che si è però oggi affermato nelle
gimnosperme e nelle dicotiledoni.
Perché è necessario che la pianta abbia questo tipo di accrescimento
secondario?
Necessità primaria di una pianta è quella di svolgere un’efficiente attività
fotosintetica, che richiede l’allungamento del fusto in maniera da portare le
foglie sempre + in alto.
Vi sarebbero problemi di staticità se non intervenisse l’accrescimento in
spessore, che permette anche lo sviluppo di nuovi fasci conduttori in
collegamento con le foglie neo formate.
Nelle gimnosperme e nelle dicotiledoni l’accrescimento secondario in spessore è
assicurato dall’attività di due meristemi:
1.
Cambio cribro vascolare, che porta all’accrescimento del tessuto vascolare
2.
Cambio subero fellodermico, che origina un nuovo tessuto di rivestimento,
il periderma.
1-Cambio cribro vascolare
Sia nel fusto che nella radice, il cambio cribro
vascolare è costituito da un anello continuo di
due tipi di cellule :
1.
Iniziali fusiformi che sono allungate
longitudinalmente, appiattite ed affusolate
alle estremità e per mezzo di divisioni
periclinali aumentano gli strati di cellule
che formano il corpo della pianta.
Quando una si divide, una delle due cellule figlie,
dopo aver prodotto per mitosi un numero
variabile di derivate, si differenzia, mentre
l’altra cellula figlia rimane meristematica ed in
grado di dividersi ancora.
Le cellule disposte internamente al cambio
cribro-vascolare si differenziano in
tracheidi, elementi delle trachee o altre
cellule dello XILEMA (o legno)
SECONDARIO, mentre quelle esterne si
differenziano in cellule cribrose o
elementi dei tubi cribrosi, cellule annesse
o altre cellule del FLOEMA (o libro)
SECONDARIO.
Cosi lo xilema secondario si trova in contatto
con quello primario, più interno, e il
floema secondario è in contatto con il
primario più esterno. Questa è detta
attività dipleuretica
Iniziali dei raggi, sono
molto più corte delle iniziali
fusiformi, si dividono e
producono raggi midollari.
Questi sono costituiti da cellule
vive parenchimatiche che
hanno funzione di riserva e
di trasporto delle sostanze
nutritive.
Il numero delle derivate
xilematiche è in genere
maggiore di quello delle
derivate floematiche e ciò fa
si che dal cambio si origini
molto + xilema secondario
verso l’interno ed un
quantitativo inferiore di
floema secondario verso
l’esterno.
2.
L’attività del cambio cribro vascolare permette al
fusto ed alla radice di accrescersi in spessore, e
di conseguenza deve essere compensato da un
aumento della circonferenza del cambio stesso,
mediante divisioni anticlinali (divisioni
perpendicolari alla superficie dell’organo) delle
sue cellule.
I tessuti esterni al cambio, in seguito alla sua attività,
vengono spostati e schiacciati verso la periferia
del fusto tanto che del floema restano evidenti
solo le fibre.
Tali tessuti, non riescono a seguire l’accrescimento
in spessore del fusto; una conseguenza sarebbe
la loro lacerazione se non si differenziassero nel
frattempo, per attività di meristemi residui siti
nei raggi midollari, nuove cellule
parenchimatiche in grado di occupare gli spazi
vuoti creati.
Poiché le formazioni del floema secondario vecchie
e lontane dal cambio, subiscono una dilatazione
maggiore di quelle giovani e vicine al cambio,
assumono una caratteristica forma a cuneo detti
RAGGI DI DILATAZIONE DEL FLOEMA.
Questo cambio rimane in attività fino alla morte della pianta, assicurando un
accrescimento diametrico secondario potenzialmente illimitato.
Nei fusti con sifonostele, il cambio cribro vascolare appare fin dall’inizio come un
anello continuo di cellule meristematiche interposte tra i fasci di XILEMA e
FLOEMA primari perciò è detto CAMBIO INTERFASCICOLARE di origini
primaria.
Nella maggior parte delle gimnosperme e delle dicotiledoni tuttavia il tipo di stele
presente è l’eustele, dove è distinguibile un cambio intrafascicolare di origine
primaria, interposto tra xilema e floema primari e, un cambio interfascicolare
che si origina da un meristema secondario a partire da cellule parenchimatiche dei
raggi midollari.
Anche qui il cambio è circolare sin dall’inizio e produce xilema verso l’interno e
floema verso l’esterno.
Cambio subero-fellodermico
A differenza del cambio cribro-vascolare il fellogeno è sempre in tutta la sua
estensione, un meristema di origine secondaria in quanto deriva da cellule già
differenziate che riacquistano le caratteristiche meristematiche.
L’attività del fellogeno permette di sostituire i tessuti primari di rivestimento di
fusto e radice, che a causa della dilatazione tendono a lacerarsi, con un tessuto
secondario: il PERIDERMA.
Il fellogeno (≠ dal cambio cribro vascolare) è formato da un solo tipo di cellule
iniziali, ma anch’esso presenta attività dipleurica, in quanto dalla divisione
delle sue cellule origina sughero(+) verso l’esterno e felloderma(-) verso
l’interno.
Sughero, fellogeno e felloderma vanno a costituire il PERIDERMA
Il primo cambio subero fellodermico, si origina nei fusti a
partire da cellule parenchimatiche sottoepidermiche o a
volte dalle stesse cellule epidermiche (es.melo, pero).
I tessuti esterni al fellogeno, sono soggetti a stiramento e solo
in alcune piante (acero, rosa, vischio) l’epidermide si
accresce per dilatazione e può quindi persistere per diversi
anni al di sopra del periderma.
D’altronde, poiché il sughero, prodotto verso l’esterno dal
fellogeno, è impermeabile alle sostanze trasportate dai
fasci conduttori del cilindro centrale, tutte le cellule esterne
ad esso sono comunque destinate a morire.
Solo in alcune angiosperme il primo fellogeno è attivo per anni (es. betulla) o
addirittura per tutta la vita della pianta (es faggio, quercus suber).
Di solito invece, il primo fellogeno cessa ben presto la propria attività
trasformandosi anch’esso in sughero.
Un nuovo fellogeno si origina allora da strati più profondi della corteccia, ma
morirà anche questo.
Si ripete nel corso degli anni questo processo e il cambio s.fellodermico arretra
nell’origine, e il fellogeno si forma anche a livello del floema primario e, in
seguito, addirittura dal floema secondario.
In questo modo il floema più vecchio, dato che è esterno al fellogeno muore e
viene eliminato con la caduta del ritidoma.
RITIDOMA indica tutti i tessuti morti, esterni all’ultimo fellogeno formato e
corrisponde a quella che comunemente è indicata come SCORZA O
CORTECCIA degli alberi.
In una struttura con accrescimento secondario il termine corteccia è usato
propriamente per indicare tutti i tessuti, sia vivi sia morti , esterni al cambio
cribro-vascolare.
Il fellogeno, oltre a contribuire all’accrescimento diametrico, interviene nei
processi di cicatrizzazione.
Nei tessuti lacerati le cellule parenchimatiche vive presenti in prossimità della
ferita iniziano a produrre un “callo da ferita”; in seguito alla periferia del callo,
entra in attività un fellogeno che produce uno strato di sughero protettivo verso
l’esterno.
Gli anelli di crescita
Molto evidenti, nelle sezioni trasversali di fusti di piante che vivono nelle zone temperate, sono
un gran numero di anelli concentrici chiamati anelli di crescita. Osservando la struttura di
questi anelli al microscopio ottico, si può notare che essi si originano in quanto le cellule
conduttrici dello xilema, non hanno tutte lo stesso diametro.
Ad elementi conduttori con lume molto grande, fanno seguito, procedendo dal centro verso il
cambio c.v. elementi conduttori dal diametro progressivamente minore, evidenziabili come
zone più scure perché con minore contenuto di aria, finché ad elementi molto piccoli,
seguono bruscamente elementi con diametro molto grande.
Ciò si spiega in quanto il cambio di piante legnose di zone temperate, alla ripresa in primavera
dell’attività vegetativa, sotto lo stimolo di ormoni prodotti dalle gemme, produce elementi
con pareti sottili, facilmente estensibili e dal diametro grande, per garantire l’appropriato
apporto di acqua alle gemme che si devono aprire.
Con il sopraggiungere dell’estate, invece, il cambio inizia a produrre elementi sempre più piccoli
e con pareti fortemente ispessite e lignificate, finché al cessare dell’attività meristematica del
cambio (in genere ad Agosto), si interrompe la produzione di nuovo xilema. Nella primavera
successiva, il cambio riprende a produrre elementi grandi che, a stretto contatto con quelli
piccoli e con pareti spesse dell’anno precedente, creano un confine netto tra il legno
primaverile e quello estivo. Ogni anno un nuovo anello si aggiunge ai precedenti.
(45X-SEM) Pinus echinata(il pino pece) evidente il legno
primaverile(larghe tracheidi)ed il legno estivo(strette). I
raggi midollari sono uniseriati, formati da un’unica fila
di cellule, parenchimatiche.
Si notano canali resiniferi.
Il legno delle conifere è detto omoxilo perché formato da un
unico tipo di cellule che hanno funzione sia di sostegno
che di conduzione.
Per la sua uniformità il legno omoxilo è adatto alla
conduzione delle vibrazioni e vieni quindi preferito per
la costruzione di strumenti musicali.
Questo legno è privo di fibre ed è spesso denominato
softwood (nelle angiosperme ed in alcune conifere
come Pinus elliottii c’è l’hardwood).
La formazione di un legno estivo diverso da
quello primaverile, non è tanto dovuta alla
mancanza d’acqua in estate quanto ad una
variazione, sia qualitativa che quantitativa,
nella produzione di ormoni dalla parte delle
gemme, che in questo periodo cessano di
svilupparsi.
Se si eliminano le gemme apicali di una pianta,
l’attività del cambio cessa, per riprendere se
al suo apice si usano ormoni come le
giberelline o le auxine.
Esiste quindi uno stretto rapporto tra la
formazione di foglie e l’attività del cambio.
Ovviamente il fenomeno degli anelli di crescita
è presente solo in quelle piante che vivono in
zone dove il cambio è attivo periodicamente
e dunque dove esiste l’alternanza delle
stagioni.
Nelle zone equatoriali, dove la temperatura è
praticamente costante durante tutto l’anno, il
cambio è sempre attivo e non si hanno quindi
anelli di crescita.
Il legno privo di anelli è molto pregiato ne è un
esempio l’ebano. Se in una stagione
vegetativa (corrispondente ad un anno nelle
regioni temperate) si forma un solo anello di
crescita, questo viene detto anche cerchia
annuale.
(SEM 100X Quercus rubra) Si vedono elementi
conduttori del legno primaverile(poche
tracheidi e numerose trachee) con ampio lume
e, quelle estive con lume ridotto.
Il legno delle angiosperme è detto eteroxilo per la
presenza delle trachee e delle fibre deputate al
sostegno e, per la natura dei raggi midollari
che possono essere uniseriali(come nelle
gimnosperme) o multiseriali, ossia formati da
più file di cellule.
La presenza di grossi vasi (trachee) e di lunghe
fibre, fanno si che il legno delle angiosperme
sia costituito da file disordinate di cellule.
Il legno delle angiosperme è definito hardwood
anche se solo in alcune angiosperme il legno è
veramente duro(bosso, mogano, olivo,
ligustro). In altre angiosperme, il legno è dolce
come nel caso del legno di balsa.
Tutto dipende dalla proporzione relativa di
elementi di conduzione, fibre e cellule
parenchimatiche.
Nel legni duri quindi prevalgono le fibre, i vasi
con lume piccolo e le cellule parenchimatiche
sono poco numerose.
Legno secondario
Il legno secondario prodotto dall’attività del
cambio cribro vascolare svolge
principalmente 3 funzioni:
1.
Lo xilema prossimo al cambio,
recentemente differenziato, assicura il
TRASPORTO dell’acqua e sali in essa
disciolti.
2.
Allontanandosi dal cambio, andando
verso il centro, si inizia a perdere
gradualmente la funzione di trasporto e
inizia quella di SOSTEGNO (tille,
protuberanze delle cell.
parenchimatiche, occludono i vasi ed
entra aria facendo perdere funzionalità).
3.
RISERVA: le cellule parenchimatiche
durante il riposo vegetativo,
immagazzinano le sostanze elaborate
dall’azione fotosintetica. Il legno +
interno, + vecchio, non svolge questa
funzione, perché qui le cellule sono
morte.
La sezione di alcuni tronchi (Prunus, mogano, teak, ebano…) presentano
2 zone facilmente distinguibili:
Alburno: esterno e prossimo al cambio cribro vascolare, è la zona+
chiara, è lo xilema funzionante nel trasporto dell’acqua e svolge
funzione di riserva per le cellule parenchimatiche vive.
Duramen: interna e lontana dal cambio cribro vascolare e costituita dal
legno più scuro, più vecchio. Lo xilema non è più funzionante, le
pareti cellulari sono impregnate di tannini, oli essenziali, resine che
sono antidegradative e antifermentative, che non fanno marcire il legno
e danno ad esso un colore particolare, ne incrementano la durezza, il
peso l’impermeabilità.
È quello commercialmente + pregiato.
In alcuni alberi, mancano queste impregnazioni del duramen (salice) ed il
tronco può per l’azione di funghi, divenire cavo.
Nell’Acer pseudoplatanus, Acer platanoides ad esempio è presente solo
l’alburno.
Libro secondario
Il floema funzionante è solo quello dell’ultimo anno, più
prossimo al cambio.
Esternamente a questo troviamo del tessuto floematico che
mantiene solo la funzione di riserva.
In ogni caso, lo spessore totale del floema è sempre ridotto in
quanto, con il procedere dell’accrescimento diametrico
della pianta, esso viene schiacciato verso i margini del
fusto e di esso rimangono solo le fibre.
Anche qui il lume degli elementi cribrosi viene occluso da un
callo in modo da divenire non funzionale.
A seguito dell’attività del fellogeno, inoltre, il vecchio floema
viene eliminato con la caduta del ritidoma.
Nelle piante erbacee, l’asse caulinare non
possiede cellule con pareti lignificate e
prende il nome di fusto. Negli arbusti e
negli alberi al contrario l’asse lignifica,
diventa rigido e prende il nome di tronco.
La porzione dell’asse compresa fra il colletto
e le foglie cotiledonari viene detta asse
ipocotile. La prima parte del fusto che si
forma dopo la germinazione .
Fra i cotiledoni e la prima foglia è situato
l’asse epicotile.
Il punto di inserzione della foglia sull’asse è
spesso ingrossato e va a formare un nodo.
La porzione di fusto fra due nodi
consecutivi si dice internodo.
La crescita intercalare dovuta ai meristemi
permette l’allungamento del fusto e
l’allontanamento dei nodi. Il fusto dove
gli internodi hanno raggiunto la loro
massima lunghezza viene detto
macroblasto.
Nelle piante primitive la ramificazione era
biforcata(dicotomica). La ritroviamo
ancora nelle briofite e nelle pteridofite
attuali(Psilotum).
Ha inizio a livello di una cellula apicale
che si divide longitudinalmente in
determinati momenti
dell’accrescimento del fusto. I due
rami della forcella si sviluppano a
partire da due cellule figlie.
I fusti delle spermatofite, danno origine a
ramificazioni che nascono dalle
gemme laterali (o ascellari).
Una gemma corrisponde ad un cono
vegetativo racchiuso da foglie
modificate con funzione protettiva. Le
gemme laterali sono situate sul fusto
all’ascella delle foglie
Le foglie possono presentare
all’ascella una o + gemme a
seconda della specie.
Le ramificazioni del fusto si
sviluppano da gemme ascellari.
Se un ramo laterale non si allunga
viene detto brachiblasto.
In parecchie specie arboree, ad es.
negli alberi da frutta si osservano
brachiblasti che portano fiori e
successivamente frutti. Questi rami
sono detti fruttiferi. Le foglie di
larici e pini sono alla stessa stregua
inserite su brachiblasti.
Il fusto di numerose piante legnose si
allunga con macroblasti durante il
primo anno mentre l’anno seguente
si sviluppano brachiblasti laterali
Le modalità con le quali il fusto può ramificarsi possono essere raggruppate essenzialmente in
due tipi principali:
Ramificazione monopodiale
Ramificazione simpodiale
La forma del cono è determinata dalla posizione delle gemme laterali che si svilupperanno.
Si distinguono un accrescimento:
Acrotono
Basitono
Le regole che si applicano alla ramificazione del fusto valgono anche per le infiorescenze
Tipi di fusto metamorfosato
RIZOMA:
E' un fusto strisciante e sotterraneo che può ricordare per il suo aspetto,
l'apparato radicale. La sua vera natura tuttavia si manifesta anche
esternamente per la presenza di foglie squamiformi , di radici avventizie e di
gemme.(Esempio: gramigna)
TUBERO:
Anche questo è un organo ipogeo che per di più si trova allo stesso livello
delle radici , quindi più facilmente può confondere un osservatore superficiale.
Dimostra però la sua origine caulinare per la presenza delle gemme.(Esempio:
patata)
BULBO:
E' un fusto accorciato e claviforme (girello) attorno al quale si trovano
numerose foglie divenute carnose, perché ricche di materiale di riserva. Da
esso, in basso, si dipartono le radici, in alto le foglie e lo scapo fiorifero.
(esempio : Cipolla, Giacinto , ecc..
STOLONI:
Sono rami striscianti, simili ai rizomi; si trovano però sulla superficie del
suolo. Si osservano frequentemente nelle fragole e possono, staccandosi dalla
pianta madre, originare piante indipendenti.
CLADODI:
Sono rami appiattiti e con l'aspetto di foglie. Portano tuttavia il fiore e poi il
frutto nella loro porzione più centrale e non alla loro base. (Esempio:
Pungitopo)
FUSTI SFERICI E COLONNARI:
Si trovano nelle piante grasse, cioè in quei vegetali adattati agli ambienti
caldo-secchi. La loro forma globosa o cilindrica è in funzione del fatto che
essi rappresentano organi di riserva acquifera . Sono generalmente verdi
perchè mancano di foglie funzionanti, essendosi queste trasformate in spine.
VITICCI:
Sono rami trasformatisi in organi di presa; si trovano comunemente nella vite
e servono per sostegno alla pianta.
SPINE:
In alcuni casi i rami assumono l'aspetto di spine più o meno grosse. Queste si
distinguono dagli aculei (propri ad esempio della Rosa) che sono semplici
produzioni epidermiche , perché presentano la struttura interna del fusto.