402
IL SISTEMA RESPIRATORIO
sono invaginazioni dell’esoscheletro rivestite di cuticola
(o Figura 9.19).
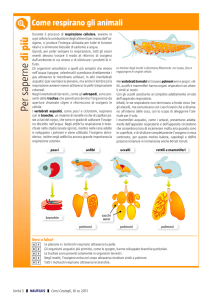
I polmoni lamellari (quattro paia negli scorpioni, uno o
due paia nei ragni, due paia negli amblipligi e negli uropigi) sono contenuti in un’invaginazione sclerificata a forma
di tasca della parete addominale ventrale (atrio) che comunica con l’esterno tramite uno spiracolo a fessura (stigma).
Dalla parete anteriore dell’atrio originano numerose evaginazioni secondarie di forma lamellare e fogliacea che massimizzano l’area della superficie di scambio respiratorio a
contatto con l’emolinfa di cui sono riccamente irrorate.
Corte barre simili a pioli sporgono dalle lamelle e impediscono che queste ultime collassino l’una sull’altra, mantenendo così aperti al passaggio dell’aria gli spazi interlamellari. L’aria entra ed esce dal polmone principalmente
per diffusione. La ventilazione può avvenire per mezzo di
un muscolo che contraendosi dilata l’atrio determinando
l’inspirazione, mentre il ritorno elastico delle pareti atriali
produce l’espirazione.
Le trachee sono strutture tubulari che si ramificano da
uno stigma sulla superficie del corpo fino all’emocele e si
trovano soprattutto negli aracnidi di piccole dimensioni, nei
quali l’elevato rapporto superficie/volume comporterebbe
un’eccessiva perdita di acqua dai polmoni a libro. Alcuni ragni, pseudoscorpioni e ricinulei sono provvisti di cosiddette
trachee a setaccio, simili ai polmoni a libro ma con strutture
tubiformi ramificate al posto delle lamelle. Opilionidi, acari,
solifugi e la maggior parte dei ragni possiedono invece trachee tubulari (ramificate e non).
La maggior parte dei ragni possiede sia un paio di polmoni lamellari (anteriori) sia una serie di trachee tubulari
(posteriori). Alcuni aracnidi di piccole dimensioni (palpigradi e alcuni acari) sono sprovvisti di superfici respiratorie
specializzate e pertanto utilizzano la respirazione cutanea.
Le trachee si ramificano in tubi cuticolari endocellulari
del diametro di circa 0,2 µm detti tracheole, che sono situate
nel citoplasma delle cellule tracheolari e si prolungano per
prendere contatto con le singole cellule (o Figura 9.20).
La parte terminale delle tracheole è piena di un liquido (il
liquido tracheolare) nel quale l’O2 si scioglie prima di diffondere nelle cellule tracheolari. In alcuni casi, come nei
muscoli deputati al volo, esse penetrano all’interno del citoplasma distribuendo O2 direttamente ai mitocondri.
Il flusso di aria all’interno delle trachee avviene per ventilazione attiva e per diffusione: la prima interviene nelle trachee di maggiore diametro grazie alla presenza di muscolatura, la seconda si realizza all’interno delle tracheole. Alla ventilazione attiva contribuiscono i sacchi aerei, i quali collassano
e si espandono in relazione all’attività di muscoli esterni. Un
contributo alla ventilazione attiva viene dato dai movimenti
ritmici di compressione ed espansione della parete dell’addome, mentre i movimenti della testa e del protorace spingono
l’aria nelle trachee. La ventilazione è ulteriormente aumentata dalla compressione del torace, prodotta dall’attività dei
muscoli del volo. Recenti studi effettuati su grilli, scarabei e
formiche, utilizzando raggi X a elevata intensità, hanno dimostrato che le trachee presenti nella testa e nel torace sono
compresse indipendentemente dal movimento di questi insetti, ma il meccanismo non è ancora noto.
Alcune insetti molto piccoli (proturi e collemboli) sono
privi di trachee e gli scambi gassosi avvengono attraverso la
superficie del corpo.
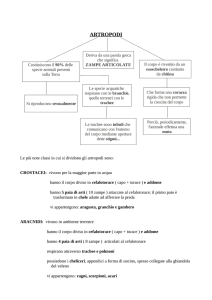
Crostacei I decapodi terricoli come i granchi delle
noci di cocco sono dotati di branchie ridotte e presentano
camere branchiali trasformate in polmoni riccamente vascolarizzati deputati agli scambi gassosi. Come nelle specie
acquatiche, lo scafognatite genera la corrente di ventilazione, costituita ovviamente da aria. Negli isopodi terrestri, la
porzione esterna di alcuni pleopodi (esopodite) ha sviluppato internamente un sistema di invaginazioni epidermiche simili alle trachee (pseudotrachee) che rappresenta la
superficie di scambio respiratorio, integrata con la respirazione cutanea.
Insetti Gli scambi gassosi negli insetti avvengono grazie
ad un sistema di trachee che si apre all’esterno con stigmi
(la cui apertura in alcune specie è occlusa da lobi sclerotizzati collegati a muscoli) e che distribuisce O2 direttamente
ai tessuti senza l’intermediazione del sistema emale. L’entità di scambio respiratorio è quindi strettamente correlata all’attività metabolica e al consumo di O2. Gli stigmi si
aprono generalmente in un atrio e sono dotati di un meccanismo di chiusura e di un filtro, per limitare la perdita d’acqua e l’ingresso di particellato e parassiti. In alcuni insetti
di grandi dimensioni (per es. le api) le trachee si dilatano in
alcuni punti formando sacchi aerei.
o FIGURA 9.20 Negli insetti, l’aria che entra in una trachea
attraverso lo spiracolo fluisce tramite le tracheole fino alle terminazioni a fondo cieco che sono in contatto con tessuti specifici
come i muscoli.