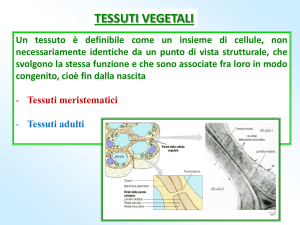
TESSUTI VEGETALI
Sono:
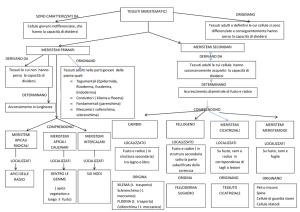
tessuti meristematici:
embrionali – primari – secondari – avventizi
tessuti di rivestimento: epidermide – rizoderma – endoderma – sughero
tessuti parenchimatici: clorofilliano – di riserva – di riempimento
tessuti di conduzione: xilema (legno) – floema (libro)
tessuti di sostegno:
collenchima – sclerenchima
tessuti segregatori (o secernenti)
MERISTEMI
Tessuti indifferenziati con cellule in continua divisione. Le cellule non presentano parete
secondaria, hanno vacuoli piccoli e numerosi.
I diversi meristemi si possono distinguere in base a:
• origine e periodo di attività: embrionali, primari, secondari
• localizzazione: apicali, laterali (intercalari)
• durata dell’attività: indefinita, definita, scalare (a gradino)
A questi si possono aggiungere meristemi avventizi che non rientrano nel programma di
crescita della pianta e si osservano, per esempio, in caso di lesioni.
• I meristemi embrionali sono presenti sin dalle prime divisioni cellulari dell’embrione e
sono localizzati ai poli dell’apice del germoglio e dell’apice della radice.
• Il meristema apicale del germoglio è in posizione effettivamente terminale e presenta
due abbozzi fogliari laterali. Il meristema apicale della radice è in posizione subterminale,
protetto dalla “cuffia radicale”, e non presenta abbozzi fogliari; in diverse specie
comprende un centro quiescente (con cellule a lentissima divisione).
• I meristemi primari, derivanti direttamente da quelli embrionali e parzialmente
coincidenti con essi, sono responsabili principalmente dell’accrescimento primario, lungo
l’asse maggiore della pianta (generalmente in altezza), mentre i meristemi secondari sono
responsabili dell’accrescimento secondario, perpendicolarmente all’asse maggiore
(diametrico). I principali meristemi secondari sono il cambio cribro-vascolare e il
fellogeno (o cambio subero-fellodermico). Entrambi danno origine a tessuti differenziati
sia verso l’interno sia verso l’esterno della pianta. I meristemi secondari sono esclusivi
delle gimnosperme delle dicotiledoni; salvo poche eccezioni mancano nelle
monocotiledoni.
• Il cambio cribro-vascolare dà origine allo xilema (legno) verso l’interno e al floema
(cribro o libro) verso l’esterno. Ha due tipi di cellule, un tipo disposto secondo la
longitudine, fusiforme che danno origine agli elementi di conduzione (vasi, tracheidi, tubi
cribrosi), e un altro disposto lungo il raggio della pianta che origina gli elementi dei raggi
midollari disposti radialmente. Nei climi stagionali l’attività del cambio presenta periodi di
pausa durante la stagione avversa.
• Il fellogeno è un tipico meristema “a gradino” che nei climi stagionali cessa la sua
attività e si riforma nella successiva stagione favorevole in posizione più interna rispetto
alla precedente. Esso dà origine al sughero verso l’esterno e al felloderma internamente.
• Tipici meristemi definiti sono quelli fogliari, con cellule per lo più raggruppate in “isole”
lungo i margini e le nervature.
TESSUTI DI RIVESTIMENTO
I tessuti di rivestimento hanno funzione protettiva e, soprattutto, evitano alla pianta
un’eccessiva perdita di acqua.
L’epidermide riveste le parti aeree, giovani, della pianta. E’ costituito da uno strato (più
raramente da pochi strati) di cellule vive particolarmente appiattite, con parete esterna
ispessita, coperta da uno strato di cuticola più o meno spessa e talora anche da cere.
Mancano spazi intercellulari, ma sono presenti delle aperture regolabili chiamate stomi.
Gli stomi presentano l’apertura circondata da due cellule di guardia a conformazione
particolare e generalmente provviste di cloroplasti. L’aumento di turgore nelle cellule di
guardia permette l’apertura dello stoma, mentre la diminuzione ne consente la chiusura;
ciò è indispensabile per la traspirazione e la fotosintesi. All’epidermide possono essere
annessi tricomi (peli) con funzione di rivestimento e/o peli ghiandolari con funzione di
secrezione.
Il rizoderma riveste la parte giovane della radice, corrispondente alla zona pilifera. Non
ha solo funzione di rivestimento, ma anche di assorbimento dell’acqua e dei sali minerali.
E’ caratterizzato da un solo strato di cellule dotate di prolungate estroflessioni chiamate
peli radicali, che aumentano enormemente la superficie della radice che viene a contatto
con l’acqua nel terreno. Le cellule del rizoderma, fortemente usurate dal contatto con le
particelle del terreno, muoiono rapidamente e nella parte inferiore della radice vengono
sostituite da nuove cellule prodotte dal meristema radicale.
L’endoderma è un tessuto particolare che non riveste la pianta perifericamente, ma
“internamente”. Si ritrova tipicamente nella radice, ma anche nei fusti sotterranei
trasformati (rizomi) e nelle foglie aghiformi e primitive delle gimnosperme. L’endoderma
è costituito da un singolo strato di cellule posto fra la corteccia e il cilindro centrale. Nelle
dicotiledoni tutte le cellule dell’endoderma presentano caratteristici ispessimenti di
suberina-lignina chiaramente osservabili al microscopio (bande o punti di Caspary). In
questo modo l’acqua non può entrare nel cilindro centrale attraverso gli spazi intercellulari
e dalle pareti ma è costretta a passare attraverso le cellule da cui può essere meglio
“controllata”. Inoltre l’endoderma, separando il cilindro centrale dall’esterno, separa due
comparti a concentrazione molto diversa, condizione necessaria per il richiamo dell’acqua
nei vasi e la sua salita alle foglie.
Il sughero si osserva soprattutto nelle porzioni non più giovani di fusto e radici. E’ un
tessuto morto, generalmente pluristratificato, con cellule, regolarmente impilate, a parete
suberificata. La notevole suberificazione conferisce un forte potere impermeabilizzante e
coibente che limitano fortemente la perdita d’acqua e gli sbalzi termici. Il sughero presenta
comunque aperture non regolabili chiamate lenticelle, attraverso le quali possono avvenire
limitati scambi gassosi; di regola le lenticelle si formano in corrispondenza dei preesistenti
stomi e vanno incontro a cicli di occlusione e riapertura coincidenti col variare delle
stagioni.
TESSUTI PARENCHIMATICI O PARENCHIMI
Sono considerati i tessuti “fondamentali” delle piante perché comprendono i tessuti
deputati alla fotosintesi. la loro specializzazione non è molto spinta e pertanto sono in
grado di tornare allo stato meristematico, dando poi luogo a tessuti diversi. Le cellule sono
tondeggianti o a spigoli arrotondati e distinte fra loro da ampi spazi intercellulari.
I parenchimi clorofilliani hanno la funzione di svolgere la fotosintesi e si trovano
perifericamente in tutte le parti verdi della pianta, principalmente nella foglia e nei fusti
giovani. Nella foglia si possono presentare sotto due forme differenti.
Nel parenchima a palizzata, con cellule allungate, disposte parallelamente in uno o pochi
strati, ricche di numerosi cloroplasti dotati di sistema tilacoidale particolarmente
sviluppato, è privilegiata la fase luminosa della fotosintesi, mentre nel parenchima
lacunoso, caratterizzato da ampi spazi intercellulari, cellule arrotondate a minor numero di
cloroplasti, viene privilegiata la fase oscura. In diverse specie, tuttavia, non è possibile il
parenchima clorofilliano non è nettamente distinguibile in questi due aspetti.
I parenchimi di riserva hanno la funzione di accumulare, conservare e fornire sostanze
diverse. Essi sono localizzati più internamente rispetto ai precedenti, particolarmente nel
fusto (soprattutto in quelli sotterranei come tubero, rizoma e bulbo), nelle radici, nei semi,
nei frutti. La maggior parte di essi va incontro a fasi di accumulo e fasi di utilizzo delle
riserve, rappresentate da amido (parenchima amilifero, ricco di amiloplasti), proteine
(parenchima aleuronico ricco di corpi proteici), lipidi (parenchima oleifero, ricco di
oleosomi), acqua (parenchima acquifero). Quest’ultimo, tipico delle piante di climi caldi
ed aridi conserva grandi quantità d’acqua per lo più grazie a molecole fortemente idrofile
(emicellulose e mucillaggini) che la trattengono. Il parenchima aerifero è invece un
particolare tessuto con ampi spazi intercellulari, che permette di portare ossigeno alle
radici, nei piccioli e nei fusti e concorre anche al galleggiamento delle foglie nelle piante
acquatiche.
Tra i parenchimi di riempimento possiamo segnalare il midollo posto in zona centrale
nei fusti e le zone periferiche dei raggi midollari che consentono l’aumento diametrico del
fusto senza che si verifichino spaccature.
TESSUTI DI CONDUZIONE
Non si tratta di tessuti semplici, ma complessi che costituiscono dei veri e propri sistemi di
trasporto.
Lo xilema o legno ha il compito di trasportare acqua e sali minerali dalle radici alle foglie
e può presentare diversi elementi di conduzione a seconda del tipo di pianta e dell’organo
e dell’età di questo. Ogni elemento di conduzione deriva dalla sovrapposizione lungo un
asse di cellule allungate che hanno subito una progressiva lignificazione delle pareti sino
alla morte. Nelle gimnosperme e nelle porzioni giovani ancora in crescita delle altre piante
(giovanissimi fusticini, parti giovani della foglia, ecc.) troviamo elementi di conduzione
xilematici più primitivi: le tracheidi, che hanno diametro molto ridotto e presentano setti
trasversali obliqui (a becco di flauto); esse non svolgono esclusivamente funzione di
trasporto, ma anche di sostegno e per questo sono chiamate fibrotracheidi. Nelle
angiosperme (eccetto alcune parti più giovani), prevalgono invece le trachee, a diametro
notevolmente maggiore e con setti trasversali quasi completamente riassorbiti, con
funzione quasi esclusivamente di conduzione. Le trachee sono definiti vasi aperti, sono
lunghe da pochi mm a diversi m (nelle liane) e sono molto più efficienti delle tracheidi
(definite vasi chiusi) per il trasporto dell’acqua.
La lignificazione delle pareti aumenta progressivamente in modo da permettere nelle
prime fasi l’allungamento degli organi, alla fine rimangono punti non lignificati detti
“punteggiature” che permettono il passaggio dell’acqua da una trachea o tracheide ad
un’altra. La lignificazione è indispensabile per conferire rigidità e resistenza alla pressione
dell’acqua. Gli elementi di conduzione sono affiancati da cellule parenchimatiche e da
fibre di sostegno. Nelle regioni con climi stagionali, la produzione dello xilema da parte
del cambio è differenziata per periodi: all’inizio della stagione favorevole vengono
prodotti maggiori elementi di conduzioni (legno primaverile), mentre alla fine vengono
prodotti meno elementi di conduzione molte più fibre di sostegno (legno estivo). La parte
periferica dello xilema (vicina al cambio), più giovane è la parte realmente funzionante per
la conduzione, mentre la parte più interna, più vecchia, svolge funzione esclusivamente di
sostegno; la prima è chiamata alburno e la seconda duramen.
Il floema o cribro o libro, al contrario dello xilema, è un tessuto vivo e ha la funzione di
condurre le sostanze elaborate con la fotosintesi dalle foglie agli altri organi. Gli elementi
caratterizzanti sono le cellule cribrose (più primitive e localizzate nelle parti in crescita) e i
tubi cribrosi. Questi sono privi di nucleo e di altri organuli; derivano dalla sovrapposizione
lungo un asse di più cellule con pareti dotate di placche cribrose fornite di numerose
perforazioni. Ogni unità (derivante da una cellula) di un tubo cribroso è accompagnata da
una cellula compagna, provvista di grosso nucleo e di numerosi organuli. Ciascuna unità
di tubo cribroso e la sua cellula compagna derivano dalla divisione della stessa cellula
iniziale e non può sopravvivere autonomamente. I tubi cribrosi non necessitano di rinforzi
di lignina come lo xilema perché la densità della linfa che vi viene trasportata è molto
maggiore di quella dell’acqua. Gli elementi conduttori del floema restano funzionanti per
un breve periodo (generalmente non più di una stagione), dopodiché cessano di
trasportare. La stagione favorevole alla ripresa vegetativa è pertanto accompagnata dalla
produzione di nuovo floema. Anche nel floema si riscontrano elementi parenchimatici e
fibre di sostegno.
Xilema e floema si dispongono negli organi delle piante in modo peculiare, spesso
differenziato a seconda dei gruppi sistematici, a formare i fasci cribro-vascolari. Nel caso
delle dicotiledoni prevalgono fasci collaterali con floema e xilema adiacenti, con uno
strato di cambio interposto. Nelle monocotiledoni, salvo poche eccezioni, i fasci hanno
floema e xilema a diretto contatto, senza l’interposizione del cambio. Anche la
disposizione dei fasci cambia a seconda che si osservi la radice o il fusto di una
monocotiledone o di una dicotiledone.
TESSUTI MECCANICI O DI SOSTEGNO
I principali tessuti meccanici o di sostegno si rinvengono nelle parti aeree e in modo più
limitato, negli organi sotterranei.
Il collenchima è un tessuto vivo caratterizzato da cellule con parete ispessita di cellulosa.
Le cellule collenchimatiche possono essere brevi o più o meno allungate e affusolate alle
estremità (fibre). Il collenchima conferisce resistenza, ma permette una certa flessibilità; si
ritrova perifericamente (di solito immediatamente sotto gli strati epidermici) in particolare
nei fusti erbacei o poco lignificati e nei piccioli. Spesso le fibre del collenchima sono
riunite in cordoni disposti lungo il fusto per irrobustirlo senza irrigidirlo. Le cellule del
collenchima sono vive e l’irregolarità degli ispessimenti della parete permette la
comunicazione intercellulare. In alcuni casi il collenchima può svolgere
contemporaneamente altre funzioni, come per esempio la fotosintesi (collenchima
clorofilliano). Quando il deposito di cellulosa aumenta le cellule collenchimatiche
muoiono e assumono il carattere di sclerenchima.
Lo sclerenchima è un tessuto morto con cellule la cui parete si è progressivamente e
completamente ispessita di cellulosa e poi si è lignificata. Lo sclerenchima irrobustisce gli
organi in cui è localizzato e ne aumenta la rigidità. Esso comprende due tipi di cellule: le
fibre sclerenchimatiche particolarmente allungate e le sclereidi (non allungate). Le fibre
sono spesso aggregate in cordoni o guaine che possono circondare completamente o
parzialmente il fusto o i fasci svolgendo una funzione di irrobustimento e protezione con
lunghezza variabile da pochi mm a qualche metro. Esse sono sfruttate industrialmente per
tessuti, cordami, ecc. (lino, canapa, juta, ecc.)
TESSUTI SEGREGATORI
Fra i tessuti segregatori si possono evidenziare quelli che formano strutture di secrezione
interna come per esempio tasche e canali resiniferi (provvisti di resine) o laticiferi (con
latice) e di secrezione esterna (peli ghiandolari, nettàri, ecc.).