La respirazione cellulare (seconda parte)
LA LEZIONE
Introduzione
Terza fase: catena di trasporto degli elettroni e fosforilazione ossidativa
Il ciclo di Krebs è seguito da un processo che ha come risultato finale la sintesi di ATP
a partire da ADP e fosfato inorganico. In estrema sintesi, tale processo utilizza come
materiale di partenza i cofattori NADH e il FADH2 prodotti durante il ciclo di Krebs, i
quali fungono da donatori di elettroni lungo un percorso che termina con l’ossigeno
molecolare e che consiste in una catena formata da molecole che trasportano gli
elettroni. In questo modo, si crea un flusso di elettroni e parte dell'energia liberata è
recuperata per sintetizzare molecole di ATP, attraverso la fosforilazione ossidativa. I
cofattori, tornati allo stato ossidato, possono poi rientrare nel ciclo di Krebs. È bene
sottolineare che il ciclo non usa ossigeno, utilizzato invece nella fosforilazione
ossidativa.
I protagonisti
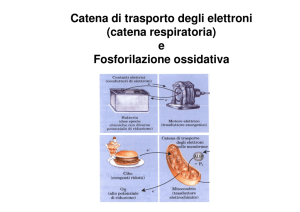
Nella membrana mitocondriale interna, schematizzata in fig.1, è localizzato un insieme
di proteine con funzione enzimatica che formano la catena di trasporto degli elettroni,
detta anche catena respiratoria.
Tali proteine costituiscono quattro complessi integrati nella membrana:
■ complesso I, noto anche come NADH-CoQ reduttasi o coenzima Q reduttasi o NADH
deidrogenasi;
■ complesso II, noto anche come succinato-CoQ reduttasi;
■ complesso III, noto anche come citocromo c reduttasi o CoQ-citocromo c reduttasi;
■ complesso IV, noto anche come citocromo ossidasi o citocromo c ossidasi.
Sono inoltre presenti due complessi enzimatici relativamente mobili (il coenzima Q, o
ubichinone, e il citocromo c) e altri enzimi, come glicerofosfato deidrogenasi, ETF
(Electron-transferring flavoprotein) deidrogenasi ecc., che cedono elettroni
direttamente al coenzima Q.
fig.1 Rappresentazione schematica del mitocondrio con
i principali processi biochimici alla base della
respirazione cellulare
La NADH deidrogenasi (fig.2) e la succinato-CoQ reduttasi (fig.3) hanno una natura
lipoproteica, sono costituite da numerose subunità e contengono, rispettivamente, i
cofattori FMN (flavinmononucleotide) e FAD. Si tratta di cofattori molto simili,
impegnati nell'ossidazione del NADH. Nei due complessi enzimatici sono presenti
inoltre proteine Fe-S (gruppi prostetici o cluster) che, grazie all'ossidazione del ferro,
permettono il trasferimento di elettroni all'ubichinone.
fig.2 Modello tridimensionale del complesso I; fig.3 Modello tridimensionale del complesso II nel batterio Escherichia coli
Per quanto riguarda i citocromi, ne esistono diversi tipi. Nella catena di trasporto degli
animali superiori sono presenti i citocromi a, a3, b, c, c1, che sono differenziati in base
allo specifico spettro di assorbimento della luce visibile. Il sistema redox dei citocromi
è il gruppo eme, contenente uno ione ferro, che può passare da uno stato ferroso,
ridotto (Fe2+), a uno ferrico, ossidato (Fe3+), e viceversa. La citocromo c reduttasi
(fig.4) è un complesso proteico contenente più citocromi, di tipo b e c1, oltre a
proteine Fe-S.
fig.4 Modello del complesso III
Invece, la citocromo c ossidasi (fig.5) è formata dai citocromi a e a3, oltre a proteine
Cu-S.
Il citocromo c (fig.6), al contrario dei precedenti, non è parte della membrana interna,
bensì in posizione periferica; inoltre è di piccole dimensioni e soprattutto è mobile,
ossia può passare da una parte all'altra della membrana. Questa proprietà gli
permette di costituire un ponte tra i complessi III e IV, poiché è in grado di trasferire
elettroni dall'uno all'altro.
fig.5 Modello del complesso IV; fig.6 Modello del citocromo c, con il gruppo eme in posizione centrale
Infine, il coenzima Q (fig. 7) è un benzochinone liposolubile dotato di una lunga
catena isoprenoide. Ha piccole dimensioni e può facilmente muoversi attraverso la
membrana interna. Trasferisce elettroni tra i complessi I e II e nella forma ridotta
prende il nome di ubichinolo.
fig.7 Le diverse forme del coenzima Q
durante il trasporto degli elettroni: in A, la
forma ossidata; in B, la forma semichinonica;
in C, la forma ridotta
Oltre che nel trasporto degli elettroni, i complessi I, III e IV funzionano anche come
pompe protoniche, perché con la loro attività permettono la formazione di un
gradiente di protoni a livello della membrana interna del mitocondrio, necessario alla
formazione di ATP. Tutti i costituenti della catena respiratoria si presentano in una
disposizione estremamente ordinata, tale che essi si susseguono in funzione
dell’aumento del proprio potenziale di ossidoriduzione (−320 mV il potenziale del
NADH, +820 mV il potenziale dell’O2, con una differenza totale di 1.140 mV). In tal
modo, si viene a creare una catena di trasporto degli elettroni nella quale ogni
costituente è fisicamente posto tra il suo riducente (che lo precede) e il suo ossidante
(che lo segue), permettendo un flusso di elettroni dal substrato (NADH o FADH 2)
all’accettore finale degli elettroni stessi (O2). Dal momento che gli elettroni sono
trasportati da uno stato energetico più alto a uno più basso si ha una diminuzione di
energia libera pari a 52,6 kcal.
È bene sottolineare che, in condizioni fisiologiche, l’ossidazione dei substrati e la
fosforilazione dell'ADP sono accoppiate; infatti, i protoni generati dall’ossidazione del
NADH (o del FADH2) si accumulano tra la matrice racchiusa dalla membrana
mitocondriale interna e la
matrice esterna alla
membrana, creando, come già
detto, una differenza di
concentrazione di ioni H+ (e
quindi un potenziale di
membrana). La presenza di
questo potenziale di membrana
permette la sintesi dell’ATP a
opera di un sistema enzimatico
complesso denominato ATP
sintetasi (o complesso V; fig.
8).
fig.8 Modello del funzionamento dell'ATP
sintetasi
Si tratta di una proteina di membrana costituita da due subunità: F 0 e F1. La subunità
F0 è un polipeptide che attraversa la membrana del mitocondrio (negli Eucarioti) o
della cellula (nei Procarioti) e costituisce un canale ionico per il passaggio di H +, dal
momento che la membrana interna del mitocondrio è impermeabile ai protoni; F 0 è
formata a sua volta da una subunità a, due b e 14 c, queste ultime organizzate a
corona. Invece F1 consiste di tre subunità α e tre β, accoppiate tra loro a formare
dimeri intorno alla subunità γ, collegata alla porzione F0; sono anche presenti subunità
δ ed ε. Il passaggio di protoni attraverso il canale ionico costituito dalle subunità c
determina la rotazione della subunità γ che, a sua volta, causa un cambiamento
conformazionale dei dimeri, esponendo il sito che lega ADP e Pi e permettendo la
sintesi di ATP.
Tappe della catena di trasporto degli elettroni
Nella prima tappa della catena respiratoria il NADH è ossidato con la liberazione di due
elettroni e la riduzione di FMN a FMNH2, che a sua volta si riossida e passa due
elettroni al gruppo prostetico (costituito da quattro centri Fe-S) del complesso I. Da
qui, gli elettroni si trasferiscono da Fe 2+ (che diviene Fe3+) al coenzima Q, che è
ridotto a QH2 (ubichinolo). L'energia liberata nel corso di questa tappa serve per
trasportare quattro protoni fuori dalla matrice mitocondriale interna; inoltre, per ogni
NADH sono mobilitati due elettroni. Esiste anche una via alternativa di ossidazione,
nella quale, al posto dell'FMNH2, è utilizzato il FADH2, successivamente ossidato dal
gruppo prostetico come sopra. L'enzima catalizzante la reazione di questa via è il
complesso II. In questo caso, non c'è trasporto di protoni.
Nella seconda tappa (nota anche come ciclo Q), il QH2 riduce a Fe2+ il Fe3+ contenuto
nel citocromo b, che a sua volta torna allo stato ossidato riducendo il Fe 3+ presente
nel citocromo c1; questo trasmette la sua capacità riducente al ferro contenuto nel
citocromo c. Il sistema enzimatico che catalizza questa tappa è il complesso III.
Durante la catena di reazioni nello spazio intermembrana del mitocondrio sono
rilasciati quattro protoni. Nella terza e ultima tappa, il Fe 2+ nel citocromo c si ossida
riducendo il Fe3+ del citocromo a, che a sua volta riduce il Fe3+ del citocromo a3, che
a sua volta riduce a Cu+ il Cu2+ presente nel complesso IV. Infine, il rame ridotto
reagisce con l'ossigeno molecolare, riducendolo a H 2O e utilizzando due H+ per ogni
atomo di ossigeno. Nello spazio intermembrana del mitocondrio sono trasferiti due
protoni.
Tirando le somme, al termine del processo nello spazio intermembrana sono stati
espulsi dieci protoni ogni due elettroni trasferiti all'ossigeno. Poiché la membrana del
mitocondrio è impermeabile ai protoni, questi escono dalla matrice attraverso i
complessi I, III e IV. Tale trasferimento genera il gradiente di concentrazione
protonica tra i due lati della membrana interna.
Fosforilazione ossidativa
I protoni possono rientrare nella matrice racchiusa dalla membrana mitocondriale
interna solo grazie ai canali ionici costituiti dal complesso V. Secondo la teoria
chemiosmotica, ormai definitivamente accettata, è proprio questo flusso di protoni
generato dall’ossidazione dei substrati ad agire da attivatore del sito catalitico del
complesso V, che così può catalizzare la sintesi di ATP a partire da ADP e P i. Si tratta
quindi di un processo endoergonico in cui l'energia elettrochimica è trasformata
nell'energia chimica necessaria alla fosforilazione dell'ADP, con la sintesi di una
molecola di ATP ogni quattro protoni. Una conferma indiretta della correttezza di tale
ipotesi è data dai cosiddetti agenti disaccoppianti della fosforilazione (per es., il 2,4dinitrofenolo), i quali inducono la fuoriuscita dalla membrana mitocondriale interna
degli ioni H+ prodotti dalla catena respiratoria, impedendo quindi la produzione di ATP.
In loro presenza, l’energia libera rilasciata dal trasporto degli elettroni viene dispersa
sotto forma di calore.
La regolazione della fosforilazione ossidativa è basata soprattutto sulle esigenze
energetiche della cellula. Infatti, quando aumenta la quantità di lavoro che questa
deve svolgere (ossia cresce l'utilizzazione dell'ATP e quindi diminuisce la sua
concentrazione cellulare), aumenta la velocità della catena di trasporto degli elettroni
e della fosforilazione ossidativa; avviene il contrario quando la cellula è in stato di
riposo (situazione in cui diminuisce la necessità di energia).
Bilancio finale del processo respiratorio
Al termine del processo di ossidazione completa in aerobiosi di una mole di glucosio a
CO2 e H2O, sono prodotte 32 moli di ATP, 28 delle quali generate dalla fosforilazione
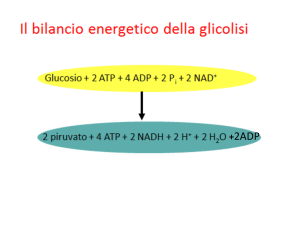
ossidativa. Infatti, partendo dalla glicolisi aerobica, la resa energetica è:
glucosio + 2 NAD+ + 2 ADP → 2 acido piruvico + 2 NADH + 2 ATP
Il processo di decarbossilazione ossidativa produce
2 acido piruvico + 2 NAD+ → 2 acetilCoA + 2 NADH + 2 CO2
Nel ciclo di Krebs, l'acetilCoA è demolito ad anidride carbonica e la resa finale è:
2 acetilCoA + 2 ADP + 6 NAD+ + 2 FAD → 4 CO2 + 2 ATP + 6 NADH + 2 FADH2
In totale, si ricavano dieci NADH, due FADH2 e quattro ATP.
Quando i coenzimi cedono il proprio potere riducente, si ha il rilascio di dieci ioni H +
per ogni molecola di NADH e sei H+ per ogni molecola di FADH2. Poiché la sintesi di
una molecola di ATP richiede l'entrata nella matrice mitocondriale di quattro protoni, e
poiché l’ossidazione di una molecola di NADH produce circa 2,5 molecole di ATP,
mentre quella del FADH2 ne produce circa 1,5, si avrà una resa finale di
10 NADH + 2 FADH2 + 6 O2 + 28 ADP → 10 NAD+ +2 FAD + 6 H2O + 28 ATP
Per quanto riguarda le molecole di ATP, 25 derivano dal NADH e 3 dal FADH 2. Al
termine del processo di ossidazione completa in aerobiosi di una mole di glucosio a
CO2 e H2O, sono quindi prodotte 32 molecole di ATP, 28 delle quali generate dalla
fosforilazione ossidativa. Questo imponente risultato rende chiaro che, in termini
energetici, la respirazione cellulare è un processo molto più efficiente della glicolisi.
Respirazione cellulare nelle piante: Caratteristiche generali
Nelle piante, il processo della respirazione cellulare è fondamentalmente simile a
quello che avviene negli animali superiori. Per una rapida rassegna al riguardo, si può
vedere questo video .
Inoltre, anche nelle piante si ha produzione di calore, ma in quantità molto meno
elevata che negli animali superiori, tanto più che gran parte del calore prodotto si
disperde rapidamente attraverso la grande superficie delle lamine fogliari; tuttavia, in
certi casi si può constatare un aumento notevole della temperatura (termogenesi). In
generale, una pianta superiore verde emette nell’atmosfera, in 24 ore, da 5 a 10 volte
il proprio volume di CO2, equivalente a 1/5-1/3 della quantità di CO2 che essa ha
fissato per fotosintesi durante la giornata.
Respirazione cellulare nelle piante: Ciclo dell'acido gliossilico
Molte piante e batteri hanno la capacità di utilizzare acidi grassi o acido acetico come
unica fonte di carbonio, soprattutto per produrre carboidrati. Questo è possibile grazie
a un processo (assente nella maggior parte dei viventi) simile al ciclo di Krebs e che è
noto come ciclo dell'acido gliossilico o del gliossilato. Nelle piante, i primi due passaggi
di tale ciclo avvengono in organelli cellulari specializzati chiamati gliossisomi,
appartenenti al gruppo dei perossisomi.
Come nel ciclo di Krebs, l'acetilCoA, derivante dalla β-ossidazione degli acidi grassi,
insieme all'acido ossalacetico porta alla formazione di acido citrico e acido isocitrico.
Successivamente, quest'ultimo composto è scisso in acido succinico e acido gliossilico,
in una reazione catalizzata dall'isocitrico liasi. L'acido succinico entra nei mitocondri e
imbocca il ciclo di Krebs, per essere trasformato in acido malico che, passato nel
citosol, è convertito in acido ossalacetico, intermedio della gluconeogenesi. Invece
l'acido gliossilico si combina con acetilCoA per produrre acido malico secondo una
reazione catalizzata dalla malico sintetasi. Questo acido malico fomerà poi acido
ossalacetico, garantendo la continuità del ciclo di Krebs.
Si sottolinea che, negli organismi incapaci di svolgere il ciclo dell'acido gliossilico, sono
assenti i suoi enzimi chiave, ossia l'isocitrico liasi e la malico sintetasi. Il risultato netto
del ciclo del gliossalato è il seguente:
2 acetilCoA + NAD+ + H2O → acido succinico + 2 CoA + NADH + H+
Respirazione cellulare nelle piante: Fotorespirazione
Le piante, soprattutto quelle C3, sono dotate di un processo stimolato dalla luce
chiamato fotorespirazione, grazie al quale una parte del carbonio organicato ottenuto
attraverso la fotosintesi viene immediatamente ossidata ed eliminata in forma di
anidride carbonica. La fotorespirazione è legata in modo particolare alle proprietà
dell'enzima ribulosiodifosfato carbossilasi (o RuDP carbossilasi o Rubisco), la cui
attività è di fondamentale importanza nei processi fotosintetici di fissazione del
carbonio.
Nel ciclo di Calvin il Rubisco interagisce con l'anidride carbonica, catalizzando la
trasformazione del ribulosio 1-5-difosfato in due molecole di acido 3-fosfoglicerico.
L'enzima è però in grado di interagire anche con l'ossigeno, comportandosi come
un'ossidasi e determinando la sintesi di acido 3-fosfoglicerico e acido 2-fosfoglicolico.
Quest'ultimo composto non entra nel ciclo di Calvin, ma viene ossidato a CO 2 e acido
3-fosfoglicerico (che continuerà il ciclo). Questo doppio aspetto del Rubisco è dovuto
al fatto che la sua attività è modulata dalla concentrazione delle molecole dei due
diversi gas, in una sorta di competizione: quando la pressione cellulare di O 2 è molto
elevata, il Rubisco lo trasforma in CO2, che rappresenta una modo per eliminare
l'ossigeno in eccesso, abbassandone la concentrazione in prossimità dei cloroplasti e
forse proteggendoli da processi fotossidativi; quando è elevata la pressione cellulare di
CO2, l'enzima fissa il carbonio nel processo fotosintetico.
Le proprietà del Rubisco sono responsabili del cosiddetto effetto Warburg (ovvero
l’inibizione della fotosintesi da parte di concentrazioni crescenti di O 2), noto sin
dall’inizio del XX secolo. Il ciclo fotorespiratorio è comunque molto complesso e si
completa nelle cellule delle foglie, principalmente C3, interessando tre diversi
comparti: i cloroplasti, i perossisomi e i mitocondri.
Bibliografia
Berg J., Tymoczko J. L. e Stryer L. "Biochimica", 2008, Zanichelli, Bologna
Garrett R. H., Grisham C. M. "Principi di biochimica", 2004, Ed. Piccin, Padova
Nelson D. L. e Cox M. M., "Principi di biochimica di Lehninger", 2006, Zanichelli,
Bologna
Voet D., Voet J. G. e Pratt C. W., "Fondamenti di biochimica", 2001, Zanichelli, Bologna