Il Ciclo Cellulare
Nel corso della loro vita tutte le cellule subiscono
una serie di cambiamenti molecolari e morfologici
definiti complessivamente ciclo cellulare.
Esso indica il periodo che intercorre tra l’origine di
una cellula da una precedente divisione ed il momento
in cui questa si divide nuovamente.
Il ciclo cellulare comprende quattro fasi sequenziali
denominate G1, S, G2 e M, oltre ad una fase G0. Nel
loro insieme le fasi G1, S, G2 costituiscono
l’interfase, mentre la fase M indica il processo di
divisione cellulare per mitosi o per meiosi. Le fasi G1
e G2 (G=Gap=intervallo) sono considerate fasi di
riposo in quanto in esse non c’è duplicazione del DNA.
Tuttavia, durante queste fasi, la cellula è
metabolicamente attiva e compie le sue funzioni
specializzate preparatorie alle successive fasi del
ciclo. Durante la fase S (S=sintesi) si attuano la
duplicazione del DNA e la sintesi di istoni, che
portano alla replicazione cromosomica.
Le cellule che vanno incontro ad un profondo differenziamento o che non si dividono
ulteriormente (es. neuroni e cellule muscolari cardiache) prolungano indefinitamente la
fase G che pertanto viene detta G0. Altri tipi cellulari, come le cellule epiteliali e gli
epatociti, possono rientrare nel ciclo cellulare dalla fase G0 e procedere con la divisione
mitotica in seguito ad appropriati stimoli quali: fattori di crescita, sostanze mitogeniche,
segnali da altre cellule e dalla matrice extracellulare.
Il sistema di controllo del ciclo cellulare è basato sull’attività di due gruppi di proteine: le cicline e le chinasi
ciclina-dipendenti (Cdk). Esse agiscono specialmente in due momenti critici del controllo del ciclo: il passaggio dalla
fase G1 alla fase S ed il passaggio dalla fase G2 alla fase M.
L’intero ciclo cellulare si svolge in un periodo compreso tra alcune ore e qualche giorno, in rapporto alla fase G1 la
cui durata varia a seconda della linea cellulare considerata.
Le prime fasi dello sviluppo embrionale sono caratterizzate da divisioni cellulari rapide, anche se con notevoli
differenze tra le diverse specie, mentre con il differenziamento delle cellule, durante l’organogenesi, la velocità
delle divisioni cellulari generalmente decresce.
Le cellule della linea germinale
(spermatogoni ed ovogoni) dopo
una serie di cicli cellulari in cui si
dividono per mitosi, vanno
incontro ad un ciclo che termina
con una divisione meiotica.
Le cellule somatiche e le cellule germinali (spermatogoni ed ovogoni) di
ogni specie hanno un corredo cromosomico completo per cui sono
denominate diploidi ed indicate con 2n.
Mitosi
Il termine mitosi è usato per descrivere la divisione equazionale di una
cellula con formazione di due nuove cellule con lo stesso corredo
cromosomico della cellula progenitrice da cui sono derivate. La mitosi è
essenziale per lo sviluppo embrionale e per riparare e rimpiazzare i
tessuti durante la vita. La mitosi può essere divisa in quattro fasi:
profase, metafase, anafase e telofase.
Durante la profase (A, B), i cromosomi, formati dai cromatidi fratelli
spiralizzano ed i nucleoli si disgregano. Nel citoplasma 2 coppie di
centrioli duplicati durante l’interfase, cominciano a formare il fuso
mitotico distinto in un fuso mantellare ed uno cromosomiale, responsabile
del successivo movimento dei cromatidi verso i poli opposti della cellula in
divisione. Segue una breve pro-metafase (C), caratterizzata dalla
disgregazione dell’involucro nucleare, e quindi la metafase propriamente
detta (D), caratterizzata dall’organizzazione dei cromosomi all’equatore
della cellula. Il cinetocore, un complesso proteico che si forma sui
centromeri alla fine della profase, agisce come una piattaforma di
attacco per i microtubuli del fuso. Si forma quindi un complesso
microtubulo-cinetocore che consente il movimento dei cromosomi che si
posizionano a metà strada tra i due poli della cellula, in una regione
denominata piastra metafasica o equatoriale. Ogni cromatide fratello è
attaccato al centrosoma mediante il proprio complesso microtubulocinetocore. Nell’anafase (E) si distinguono due stadi: anafase A in cui le
coppie di cromatidi fratelli si separano sincronicamente per la scissione
dei centromeri e per l’accorciamento dei complessi microtubulocinetocore. I due set di cromatidi appena separati vengono trascinati
verso i due poli opposti della cellula. Nell’anafase B lo spostamento dei
due cromatidi verso i poli opposti è accompagnato dall’allontanamento dei
poli. Nella telofase (F), i due gruppi di cromosomi identici, raggruppati ai
rispettivi poli, decondensano; inoltre, quelli con costrizione secondaria
ripristinano il nucleolo, attorno a ciascun set di cromosomi si ricostituisce
un involucro nucleare e termina la cariocinesi. Dopo la formazione dei
nuovi involucri nucleari, un anello contrattile di actina stringe la
membrana cellulare e divide il citoplasma in modo da separare
completamente le due cellule figlie. Quest’ultimo processo definito
citocinesi o citodieresi, normalmente porta alla formazione di due cellule
figlie di uguali dimensioni; occasionalmente differenti quantità di
citoplasma ed organuli possono essere distribuiti tra le due cellule figlie.
A
C
E
B
D
F
Meiosi
E’ il processo di divisione riduzionale, esclusivo delle cellule della linea
germinale e rappresenta l’ultimo evento di divisione della
gametogenesi.
La meiosi porta alla formazione di gameti aploidi a partire da cellule
diploidi in quanto consiste di una sola fase S seguita da due divisioni
consecutive. In conseguenza, dalla cellula di partenza diploide, si
formano, in due tappe, quattro cellule aploidi. Le due divisioni,
separate da una breve intercinesi, sono indicate come I e II
divisione meiotica, ognuna delle quali si può suddividere in fasi
corrispondenti a quelle della mitosi: profase, metafase, anafase e
telofase.
Tuttavia nella meiosi queste fasi presentano caratteristiche peculiari
anche se la cellula, che entra in profase I, presenta la stessa quantità
di DNA di una cellula che si avvia alla mitosi.
La profase della I divisione meiotica (A-D), durante la quale
avvengono molti eventi intracellulari cruciali, è molto lunga e può
essere ulteriormente divisa in cinque sottostadi: leptotene, zigotene,
pachitene, diplotene e diacinesi.
Questi sottostadi prendono il nome dagli eventi salienti che si
verificano nel corso della profase:
• leptotene, inizio della condensazione della cromatina che continua
per tutta la profase fino a formare cromosomi spessi e corti (A);
• zigotene, appaiamento dei cromosomi omologhi duplicati per formare
le tetradi, ad opera di un complesso di natura proteica detto
complesso sinaptinemale che persiste fino allo stadio successivo (B);
•pachitene, scambio di materiale genetico, per un processo di
frattura e successiva saldatura tra i cromatidi non fratelli dei due
cromosomi omologhi, definito crossing-over (C);
• diplotene, separazione dei cromosomi omologhi che restano ancora in
contatto nei punti in cui si è verificato il crossing-over (chiasmi) (D);
• diacinesi, condensazione completa dei bivalenti con separazione degli
stessi a livello dei chiasmi (terminalizzazione dei chiasmi).
E’ da notare che solo nella spermatogenesi, il processo di
citocinesi, successivo sia alla I che alla II telofase meiotica,
divide equamente il citoplasma tra le cellule. Nell’ovogenesi, al
contrario, dopo la I telofase una delle due cellule risultanti
trattiene la maggior parte del citoplasma divenendo ovocita
II, la più piccola delle due cellule è denominata I globulo
polare. Una distribuzione ineguale di citoplasma viene
mantenuta anche tra l’ovocita secondario ed il II globulo
polare, mentre nel caso in cui anche il I globulo polare vada
incontro alla seconda divisione meiotica, lo scarso citoplasma a
disposizione viene equamente ripartito.
In metafase I, le coppie di cromosomi omologhi si attaccano mediante i
loro cinetocori ai microtubuli del fuso e si posizionano in piastra
equatoriale. I centromeri dei due omologhi sono orientati verso i poli
opposti del fuso, mentre i due cromatidi di ciascuno omologo sono ancora
uniti a livello del centromero. Durante l’anafase I (E), le diadi, costituite
ognuna dai due cromatidi di ognuno degli omologhi, si spostano ai poli
opposti della cellula. A differenza di quanto avviene in anafase mitotica,
ai poli opposti della cellula migrano singoli cromosomi della coppia di
omologhi, ma ancora costituiti da due cromatidi che possono avere
effettuato il crossing-over. A questo punto la distribuzione dei
cromosomi omologhi di derivazione paterna e materna è casuale, ed è alla
base del principio mendeliano dell’assortimento casuale. In telofase I,
intorno a ciascun set di cromosomi separati si forma l’involucro nucleare,
mentre ha luogo la citodieresi. Le due cellule figlie derivate dalla prima
divisione meiotica contengono un numero aploide di cromosomi (un
rappresentante di ogni coppia di omologhi) con una quantità 2c di DNA
dal momento che ogni cromosoma è ancora costituito da due cromatidi.
Dopo una breve fase di riposo, chiamata intercinesi o interfase (F),
durante la quale si ha la formazione di un nuovo fuso, ciascuna cellula
figlia inizia la seconda divisione meiotica. La profase II è più breve
della profase I; il nucleo contiene un set di diadi non interamente
despiralizzate, composte ognuna da una coppia di cromatidi fratelli
connessi mediante un unico centromero. La metafase II è simile alla
metafase I poiché i cromosomi sono posizionati in piastra equatoriale dal
complesso microtubulo-cinetocore. In questo caso il centromero si è
sdoppiato in due cinetocori ognuno su ciascuno dei singoli cromatidi.
Questo consente ai microtubuli di attaccarsi a ciascun cromatide.
Durante l’anafase II (G) i singoli cromatidi legati al complesso
microtubulo-cinetocore migrano verso i poli opposti della cellula in
divisione. Alla fine della telofase II si riformano gli involucri nucleari
attorno a ciascun set di cromatidi ed il citoplasma si divide nuovamente.
Pertanto alla fine della citodieresi le cellule terminali della meiosi
posseggono n cromosomi ed una quantità c di DNA e sono quindi aploidi,
ma il loro corredo genico è rinnovato dal crossing-over e dalla
ripartizione casuale dei cromosomi (H).
GAMETOGENESI
I gameti sono cellule aploidi altamente differenziate derivanti, nel corso dello sviluppo embrionale, da cellule
germinali primordiali. Esse sono caratterizzate da un costituente citoplasmatico, il plasma germinale,
preesistente nell’uovo prima della fecondazione.
Nel corso della segmentazione (della gastrulazione nei Mammiferi), dallo zigote si formano numerose cellule
somatiche ed un limitato numero di cellule germinali primordiali in cui viene segregato il plasma germinale.
In sede extraembrionale (nel topo si rendono visibili nell’endoderma del sacco vitellino ad una settimana
dalla fecondazione, mentre nell’uomo alla terza settimana) si assiste alla proliferazione di tali cellule che
attraverso il circolo sanguigno, migrano attraverso il mesentere dorsale nei territori gonadici in sviluppo. Le
cellule germinali primordiali o protogoni sono mitoticamente attive fino alla nascita nella femmina e per tutta
la vita nel maschio. Le cellule germinali, definite spermatogoni nelle gonadi maschili e ovogoni nelle gonadi
femminili, vanno incontro ad una sequenza di sviluppo simile, che viene indicata come processo di
gametogenesi.
Durante
la
gametogenesi
avvengono
sostanzialmente 3 tipi di eventi anche se con
alcune differenze quantitative, qualitative e
cronologiche tra i due sessi:
• proliferazione mitotica dei goni, (fino alla
nascita nella femmina in cui tutti gli ovogoni
entrano in profase I; per tutta la vita nel maschio
in cui gli spermatogoni dalla maturità sessuale in
poi, possono dividersi sia per mitosi che entrare
in meiosi divenendo spermatociti primari);
• divisione meiotica, (discontinua e completa
solo in seguito a fecondazione nella femmina;
continua e completa nel maschio);
• trasformazioni morfo-fisiologiche dei futuri
gameti, (auxocitosi nella femmina durante la
profase I; spermiogenesi nel maschio dopo la
meiosi).
Ovogenesi
Spermatogenesi
Mitosi e Meiosi iniziano una volta
per l’intera popolazione cellulare di
ovogoni
Mitosi
e
continuamente
popolazione
spermatogoni
Meiosi
iniziano
per parte della
cellulare
di
La meiosi si arresta e riprende in
una popolazione cellulare più
esigua
La meiosi procede in modo continuo
Ogni meiosi produce solo 1 gamete
e solo in seguito a fecondazione,
che può avvenire mesi o anni più
tardi rispetto all’inizio della meiosi
Ogni meiosi produce 4 gameti e la
meiosi si completa in giorni o
settimane
Il differenziamento del futuro
gamete avviene nella profase I,
quando l’ovocita è diploide
Il differenziamento del gamete
avviene dopo la meiosi, a carico
dello spermatidi aploide
Tutti i cromosomi subiscono una
ricombinazione durante la profase I
I cromosomi sessuali sono esclusi
dalla ricombinazione durante la
profase I
Nella I e nella II citocinesi
una
delle due cellule risultanti trattiene
la maggior parte del citoplasma
Nella I e nella II citocinesi
il
citoplasma è equamente diviso tra le
cellule
Tutti gli eventi legati alla gametogenesi sono finalizzati all’acquisizione da parte dei gameti di una
struttura e una fisiologia in grado di conciliare tre funzioni:
• sopravvivere in ambienti diversi da quello delle gonadi;
• riconoscere cellule omologhe dell’altro sesso e cooperare in eventi legati alla fecondazione;
• fornire adeguate quantità di materiale nucleare e citoplasmatico per lo sviluppo del nuovo organismo.
SPERMATOGENESI
Le cellule germinali primordiali che colonizzano le gonadi maschili si organizzano in strutture cave definite tubuli seminiferi,
la cui parete è data da cellule somatiche chiamate cellule del Sertoli. Al di fuori del tubulo seminifero, nella compagine del
connettivo che lo avvolge, si rinvengono le cellule del Leydig che producono testosterone. In questa situazione, le cellule
germinali, rappresentate da spermatogoni di tipo A1, che hanno già subito una fase di moltiplicazione cellulare nel corso
dell’organogenesi, restano quiescenti fino alla maturità sessuale.
Le cellule del Sertoli sono in stretto contatto tra loro nella porzione basale attraverso giunzioni occludenti che nell’insieme
formano la barriera emato-testicolare. La presenza di tale barriera fa sì che il tubulo seminifero sia organizzato in due
compartimenti:
-compartimento basale in cui sono ospitati gli spermatogoni e gli spermatociti leptotenici,
-compartimento adluminale in cui sono ospitati gli spermatociti più maturi, gli spermatidi e gli spermatozoi.
La barriera emato-testicolare ha diversi significati funzionali, la sua presenza infatti permette di mantenere microambienti
diversi tra i due compartimenti in modo da favorire in quello adluminale la meiosi e la spermioistogenesi, oltrechè impedire
possibili risposte immunologiche in seguito all’esposizione di antigeni germinali o il passaggio di macromolecole provenienti
dal compartimento adluminale nella circolazione sanguigna dell’animale.
Le cellule del Sertoli svolgono anche altre
funzioni quali:
-sostegno meccanico per le cellule
germinali, le quali sono infatti impilate
lungo la parete laterale delle cellule del
Sertoli secondo il loro grado di
maturazione, avvolte da un sistema di
prolungamenti citoplasmatici delle cellule
del Sertoli,
-attività fagocitarla nei confronti di cellule
germinali degenerate e di corpi residuali,
-attività endocrina (ABP, inibina, fattore
antimülleriano).
Il fenomeno della spermatogenesi nei Mammiferi può essere suddiviso in tre fasi:
fase mitotica di moltiplicazione e maturazione degli spermatogoni A e B, in cui le cellule germinali vengono attivate e
attraverso una serie di divisioni mitotiche producono cloni di cellule denominate spermatogoni di tipo A1,A2,A3 ed A4. Gli
spermatogoni intermedi, derivanti dagli spermatogoni A4, dividendosi formano spermatogoni di tipo B, che sono in grado di
iniziare la meiosi evolvendo in spermatociti primari.
meiosi in cui si assiste al passaggio da spermatociti primari a spermatidi. ogni spermatocita di primo ordine o primario
duplica il proprio DNA e subito dopo inizia la divisione meiotica. Con la prima divisione meiotica da ogni spermatocita di primo
ordine si formano due spermatociti di secondo ordine o secondari, cellule aploidi per numero di cromosomi, ma con quantità
2c di DNA in quanto ogni cromosoma è ancora costituito da due cromatidi. Con la seconda divisione meiotica ogni
spermatocita di secondo ordine si divide in due spermatidi, cellule completamente aploidi (avendo ricevuto ognuna un
cromatide di ogni cromosoma) ma ancora indifferenziate.
spermioistogenesi in cui gli spermatidi si trasformano in spermatozoi.
E’ distinta in 4 fasi:
• fase del Golgi in cui dall’omonimo apparato si forma un
granulo acrosomico avvolto da membrana;
• fase del cappuccio in cui la vescicola acrosomica va a
rivestire i 2/3 anteriori del nucleo mentre dal centriolo distale
comincia a formarsi il flagello;
• fase dell’acrosoma in cui il nucleo si sposta alla periferia
della cellula, la cromatina si addensa, i mitocondri si
organizzano intorno alla parte prossimale del flagello;
• fase di maturazione in cui nucleo ed acrosoma assumono la
forma tipica della specie per l’azione di una struttura
microtubulare detta manchette, mentre la maggior parte del
citoplasma (corpo residuale) è fagocitata dalle cellule del
Sertoli.
Nei Mammiferi la lunghezza degli spermatozoi varia da 40 a 250 m, considerando un
nemaspermio di mammifero possiamo distinguere:
-una testa che contiene nucleo ed acrosoma (contenente spermiolisine per la digestione della
matrice della corona radiata e acrosina per la digestione della zona pellucida),
-un collo dove si trovano i centrioli di cui il distale origina il flagello,
-una coda divisibile nei tratti intermedio (assonema, guaina fibrosa, guaina
mitocondriale), principale (assonema, guaina fibrosa) e terminale (assonema).
Sotto lo stimolo dell’ormone ipotalamico FSH/LHRH, l’adenoipofisi rilascia FSH ed LH. L’FSH
agisce sulle cellule del Sertoli stimolandole a
produrre Androgen Binding Protein (ABP), una
proteina che ha alta affinità per il testosterone e ne
favorisce la concentrazione nel comparto
adluminale del tubulo seminifero. L’LH agisce sulle
cellule del Leydig stimolandole a produrre
testosterone. Questo giunto alle cellule del Sertoli
viene
trasformato
in
deidrotestosterone.
L’aumento
della
concentrazione
del
deidrotestosterone stimola le cellule del Sertoli a
produrre inibina, un ormone che agisce a livello
della adenoipofisi inibendo la produzione di FSH.
Analogamente l’aumento della concentrazione
ematica di testosterone agisce, con un
meccanismo di feed-back negativo, sull’ipotalamo
che diminuisce la produzione di FSH/LH-RH con
un conseguente effetto negativo sulla produzione
di testosterone ed ABP.
Gli spermatozoi maturi si staccano dalle cellule del Sertoli e vengono immessi nel lume del tubulo
seminifero, attraversano la rete testis del testicolo e raggiungono l’epididimo dove la loro motilità
viene incrementata da un processo di maturazione mediante cambiamenti morfo-fisiologici quali la
rimozione della goccia protoplasmatica e modificazioni nella composizione della membrana
plasmatica , la loro capacità fecondante viene tuttavia resa latente dal fattore di decapacitazione
secreto dalle cellule a pennacchio dell’epididimo stesso.
Dopo il passaggio attraverso l’epididimo gli spermatozoi si arricchiscono del liquido secreto dalle
ghiandole annesse alle vie genitali maschili. L’insieme degli spermatozoi e del liquido prodotto da tali
ghiandole, costituisce il liquido seminale o sperma prodotto in quantità e consistenza diversa nelle
differenti specie animali. Gli spermatozoi maturi vengono stoccati nella coda dell’epididimo prima di
essere eiaculati. La maggior parte degli spermatozoi non eiaculati viene gradualmente eliminata
attraverso il sistema urinario; la piccola percentuale che resta nell’epididimo va incontro a
degenerazione e viene fagocitata.
Gli spermatozoi dei Mammiferi acquistano la capacità di fecondare solo nel loro percorso lungo le vie
genitali femminili, dove vanno incontro ad un processo di capacitazione attraverso una serie di
modificazioni della membrana che facilita la reazione acrosomiale.
Specie
Volume in ml
Toro
2-10 (4)
Montone 0.7-2 (1)
Caprone 0.4-2.8 (1)
Verro
100-500 (250)
Stallone
30-300 (70)
Cane
2-15 (6)
Numero di Spermatozoi in milioni/mm3
0.3-2 (1)
2-5 (3)
3-5 (2,5)
0.025-0.3 (0.1)
0.03- 0.08 (0.12)
1-9 (3)
Consistenza
Colore
cremosa
grigio chiaro-giallastro
cremosa
bianco-giallastro
cremosa
bianco-giallastro
lattiginosa-flocculenta
bianco latte
gelatinosa-acquosa
bianco-grigiastro
acquosa
bianco latte
Dati comparativi relativi a volume, consistenza e colore dell’eiaculato in alcuni Mammiferi domestici.
OVOGENESI
Nei Vertebrati l’ovogenesi è di tipo follicolare in
quanto, terminata la fase di moltiplicazione, ogni
ovogonio si associa a cellule somatiche di forma
appiattita per formare il follicolo ovarico
primordiale. Poco prima della nascita tutti gli
ovogoni si differenziano in ovociti primari:
duplicano il DNA, iniziano la fase di
accrescimento e entrano nella profase I della
meiosi. Poco prima o subito dopo la nascita, a
seconda della specie, ogni ovocita entra in una
prolungata fase di riposo allo stadio di diplotene
della profase I, detta dictiotene. Essa dura fino
alla maturità sessuale dell’animale, quando lo
sviluppo riprende per induzione da parte delle
gonadotropine. Nei Mammiferi il processo di
ovogenesi procede in parallelo col processo di
follicologenesi,
pertanto,
durante
l’attività
sessuale delle diverse specie animali, a regolari
intervalli di tempo, avviene la maturazione di uno
o più follicoli e dei relativi ovociti. Inoltre, sia
durante la fase proliferativa, che durante quella di
riposo, una cospicua quota di follicoli primordiali
va incontro ad atresia (nella specie umana da
2000 protogoni migranti dal sacco vitellino, si
arriva a 6-7 milioni di ovogoni al 5° mese di vita
intrauterina, mentre alla pubertà la donna ha da
200.000 a 400.000 ovociti; in una vitella di tre
mesi sono presenti circa 75.000 follicoli, in una
vacca di tre anni circa 20.000, in una di nove anni
circa 2.500).
follicolo atresico
Follicolo di Graaf
ovulazione
corpo luteo
follicolo secondario
follicolo primario
follicolo primordiale
corpo luteo in regressione
Nella follicologenesi
riconoscono 3 fasi:
si
fase iniziale continua ed
ormono-indipendente in cui
l’ovocita secerne GPs e PGs
che formano la zona pellucida,
le cellule follicolari da piatte
divengono cuboidi e proliferano
fino a formare 3-4 strati noti
come cellule della granulosa. Il
follicolo contenente un ovocita
di 20-25 µm di diametro viene
ora definito primario, Gli ovociti
nutriti attraverso le cellule
follicolari
aumentano
di
dimensione superando i 150
μm.
seconda fase in cui fuori dal
follicolo si organizza una
struttura detta teca di natura
fibro-connettivale che sarà
importante nella formazione del
corpo luteo e sotto la spinta
dell’FSH e degli estrogeni
prodotti dalle cellule della
granulosa
alcuni
follicoli
crescono ancora, aumenta il
numero delle cellule follicolari e
si forma l’antro (l’ovocita viene
decentrato circondato da una
parte della granulosa indicata
come cumulo ooforo). Il
follicolo così modificato prende
il nome di follicolo secondario.
Le cellule della granulosa sono
così spinte alla periferia mentre
un piccolo gruppo di esse, che
circonda l’ovocita, protrude
nella
cavità
follicolare
costituendo il cumulo ooforo,
mentre il follicolo prende il
nome di follicolo terziario o
follicolo di Graaf.
fase finale durante la quale la periodica liberazione di FSH e soprattutto di LH provocano la completa maturazione del
follicolo (espansione del cumulo, brusco aumento del liquido follicolare, maturazione nucleare dell’ovocita, ovulazione). Il
grado di maturazione nucleare dell’ovocita varia tra le specie; nella maggior parte dei Mammiferi si arresta in metafase II,
mentre cane, cavallo e volpe hanno il blocco meiotico in metafase I. L’ovulazione consiste nel rilascio dell’ovocita da parte
del follicolo. Prima dell’ovulazione, l’ovocita e le cellule della corona radiata si staccano dal cumulo ooforo e fluttuano nel
liquido follicolare. La rottura del follicolo avviene grazie alla formazione di un’area vescicolosa sulla superficie ovarica posta
sopra al follicolo, detta stigma.
Auxocitosi
L’auxocitosi indica l’accrescimento dell’ovocita
durante la profase ed avviene in tre fasi distinte:
— auxocitosi I, che consiste nell’aumento del volume
citoplasmatico,
— auxocitosi II, in cui si ha un’intensa sintesi proteica
ed un accumulo di RNA a lunga durata,
— auxocitosi III (vitellogenesi o deutoplasmagenesi),
in cui si ha un accumulo, all’interno dell’ovocita, di
materiale plastico ed energetico (tuorlo) che dovrà
garantire lo sviluppo dell’embrione in caso di
fecondazione.
Negli ovociti, alla fine della fase di accrescimento, subito sotto la membrana ooplasmatica si
osservano dei granuli definiti granuli corticali. Essi vengono prodotti dall’azione combinata
del reticolo endoplasmatico e dell’apparato di Golgi e contengono materiale
mucopolisaccaridico ed enzimi proteolitici che entrano in funzione dopo la fecondazione come
meccanismo di blocco della polispermia.
In relazione alla quantità di vitello contenuto in esse, le uova
possono essere classificate in:
alecitiche, prive di tuorlo;
oligolecitiche, con poco tuorlo, (secondo alcuni autori
comprendenti anche le alecitiche);
mesolecitiche, con una quantità discreta di tuorlo;
macrolecitiche, con un’enorme quantità di tuorlo.
Inoltre in funzione della distribuzione dello stesso vitello le
uova possono essere classificate in:
isolecitiche, quando presentano una distribuzione
uniforme del tuorlo nel citoplasma attivo;
telolecitiche, quando, aumentando la quantità di tuorlo,
citoplasma e nucleo sono spostati verso uno dei poli della
cellula (polo animale), mentre l’altro polo è occupato dal
vitello (polo vitellino o vegetativo);
centrolecitiche, quando hanno tutto il deutoplasma
concentrato al centro intorno al nucleo, ed un sottile strato di
ooplasma libero da deutoplasma in periferia.
Oltre alla membrana plasmatica le uova possono presentare membrane accessorie che in base alla loro origine si
dividono in:
involucri primari, prodotti dall’ovocita e dalle cellule follicolari, rappresentati dal corion delle uova di Pesci, dalla
membrana vitellina degli Anfibi e degli Uccelli e dalla zona pellucida dei Mammiferi;
involucri secondari, secreti ed apposti durante la discesa lungo gli ovidutti, particolarmente sviluppati negli animali
ovipari ed ovovivipari. Essi comprendono il rivestimento gelatinoso nelle uova di Anfibi e nelle uova di alcuni
Mammiferi (cavallo, cane, coniglio), il rivestimento gelatinoso ed il guscio cheratinico prodotto dalla ghiandola
nidamentale negli Elasmobranchi, ed inoltre l’albume, la membrana testacea ed il guscio calcareo nelle uova di
Rettili e Uccelli. Anche i Mammiferi Monotremi presentano albume e guscio, coriaceo nell’echidna e calcificato
nell’ornitorinco. Tali involucri hanno funzione di protezione e di nutrizione.
CORPO LUTEO
Una volta avvenuta l’ovulazione, il follicolo collassato, che resta nell’ovaio, viene invaso dalle cellule della
teca ed insieme ad esse si trasforma in corpo luteo, organo secernente progesterone e 17β-estradiolo.
Se l’ovocita non viene fecondato il corpo luteo produce ormoni solo per un breve periodo (corpo luteo
ciclico o spurio) e poi degenera come corpus albicans. Se l’ovocita è fecondato si forma il corpo luteo
gravidico che, sotto lo stimolo dell’LH o delle gonadotropine corioniche e, in alcune specie, della
prolattina, permane per periodi più o meno lunghi della gravidanza (3 mesi nella donna, per tutta la
gravidanza nella vacca). Dopo il parto, il corpo luteo si trasforma in corpus albicans e poi degenera.
L’ovogenesi e la follicologenesi nei Mammiferi sono parte di un processo periodico a regolazione
ormonale a cui si dà il nome di ciclo sessuale.
Esso comporta trasformazioni a livello ovarico (ciclo ovarico) ed a livello della mucosa uterina (ciclo
uterino).
Il ciclo sessuale è suddiviso in 4 fasi che, a livello ovarico, sono caratterizzate da eventi peculiari:
proestro, maturazione del follicolo ovarico,
estro, ovulazione,
metaestro, formazione e successiva regressione del corpo luteo,
diestro, periodo di riposo.
In caso di fecondazione il ciclo si interrompe nella fase di metaestro per tutta la durata della gravidanza. Il
ciclo sessuale riprende dal proestro durante l’allattamento.
Il ciclo sessuale è caratterizzato nei Mammiferi domestici da manifestazioni esterne e comportamentali
durante la fase di estro (ciclo estrale), mentre nei Primati è caratterizzato dallo sfaldamento della
mucosa uterina nella fase di metaestro (ciclo mestruale).
Nelle specie domestiche i cicli si ripetono ogni 18-21 giorni per cui si parla di animali poliestrali. La vacca
e la scrofa sono poliestrali per tutto l’anno. La cavalla, la pecora e la capra sono poliestrali stagionali
(primavera ed autunno). Le specie selvatiche hanno sempre attività sessuale stagionale.
Specie
Durata del ciclo sessuale Durata dell’estro
Vacca
Pecora
Capra
Scrofa
Giovenca
Cagna
Gatta
Coniglia
Cavia
Donna
21 (16-30)
17 (14-20)
21 (15-24)
21 (18-24)
21 (16-30)
2-3 volte l’anno
3-4 volte l’anno
28
16
28
1-2
1-2
1-2
2 (1-3)
6 (2-13)
6-14
2-14
*
10 ore
*
Ovulazione
Durata della gestazione
14 ore dopo l’estro
280 (270-295)
verso la fine dell’estro
150 (144-156)
verso la fine dell’estro
150 (144-156)
24-36 ore dall’estro
115 (110-118)
1-2 giorni dopo l’estro
336 (320-355)
dal 3° al 21° giorno dall’estro
63 (60-66)
27 ore dopo il coito
58 (56-60)
10 ore dopo il coito
31 (30-33)
10 ore dall’estro
68 (62-72)
14±1 gg. dalla fase mestruale 266 (255-275)
* la durata dell’estro non è quantificabile in quanto i sintomi esterni sono poco marcati.
Parametri relativi al ciclo sessuale in alcuni Mammiferi (i dati sono espressi in giorni salvo diversa indicazione)
FECONDAZIONE
La fecondazione è il processo attraverso cui lo spermatozoo e l’ovocita si fondono
per formare lo zigote, che rappresenta il nuovo individuo.
La fecondazione consente il ripristino del numero diploide di cromosomi e la
determinazione del sesso dell’individuo; inoltre è fondamentale per la variabilità
biologica grazie all’integrazione delle caratteristiche ereditarie materne e paterne.
L’integrazione del materiale genetico paterno e materno dei due pronuclei viene
indicata come singamia.
La fecondazione può essere esterna o interna. Nei Mammiferi è interna e nonostante
durante il coito vengano deposti milioni di spermatozoi nelle vie genitali femminili,
solo alcune centinaia raggiungono la sede della fecondazione ed uno solo partecipa
alla singamia. Il coinvolgimento di più spermatozoi nella fecondazione, definito
polispermia, è tollerato in Pesci, Anfibi e Uccelli, anche se gli spermatozoi in eccesso
comunque degenerano. Invece nei Mammiferi, la presenza di spermatozoi
soprannumerari è un evento anomalo che porta alla morte dell’embrione.
Il trasporto degli spermatozoi lungo le vie genitali femminili avviene in due fasi:
una fase rapida, associata alle contrazioni muscolari delle vie genitali dopo il coito,
permette agli spermatozoi di arrivare nell’ampolla entro 15 minuti dall’accoppiamento, tali
spermi sono però a bassa vitalità.
una fase lenta gli spermatozoi si muovono dalla vagina o dall’utero verso l’istmo dove
vengono stoccati fino al momento dell’ovulazione, quando vengono rilasciati dall’epitelio
istmico da fattori specifici e riprendono un’intensa attività che contribuisce al loro
trasferimento nell’ampolla e alla penetrazione degli strati che circondano l’ovocita.
Nella vacca e nella pecora sono necessarie dalle 6 alle 12 ore perché un adeguato numero di spermatozoi
raggiunga l’istmo. La vitalità degli spermatozoi nell’istmo è di circa 20 ore nella vacca e nella pecora, e
fino a 36 ore nella scrofa. Anche nella donna gli spermatozooi rimangono vitali più giorni.
Riconoscimento specie-specifico e Capacitazione
Durante il loro soggiorno nelle vie genitali femminili gli spermatozoi subiscono modificazioni
biochimiche e fisiologiche per cui acquisiscono la capacità di fecondare. Questo processo,
definito capacitazione, comprende la rimozione di fattori inibitori derivati dal liquido seminale
che rivestono gli spermatozoi nell’epididimo, oltre alla rimozione di colesterolo dalla membrana
plasmatica in modo da renderla meno stabile e più idonea alla fusione con altre membrane. E’
stata osservata anche una perdita di glicoproteine di superficie che impediscono l’interazione
tra lo spermatozoo e la zona pellucida dell’ovocita.
La capacitazione, iniziata nell’utero e conclusa nell’istmo, permette la reazione acrosomiale che
consiste nella liberazione degli enzimi litici presenti nell’acrosoma, attraverso la fusione in più
punti tra la membrana plasmatica dello spermatozoo e la membrana acrosomiale esterna.
Negli organismi a fecondazione esterna più semplici esiste una sorta di meccanismo a tempo
per cui, subito dopo l’emissione, gli spermatozoi subiscono la reazione acrosomiale,
indipendentemente dalla vicinanza agli ovociti, questo meccanismo è conservato anche nella
cavia.
In molti altri organismi gli spermatozoi sono attratti chemiotatticamente dall’emissione di
sostanze da parte degli ovociti e subiscono la reazione acrosomiale una volta giunti a ridosso
dell’ovocita, ma questo avviene anche nel coniglio.
Negli organismi a fecondazione interna, in particolare nei Mammiferi, gli spermatozoi, grazie
alla loro intensa motilità e all’espansione del cumulo ooforo che si è verificata poco prima
dell’ovulazione, si infilano tra le maglie della matrice della corona radiata; in questi animali
infatti, la reazione acrosomiale si verifica dopo il riconoscimento specie-specifico. Lo
spermatozoo, raggiunta la zona pellucida si lega ad un recettore glicoproteico specifico
presente nella zona pellucida, la molecola ZP3; tale legame innesca la reazione acrosomiale
consentendo l’esposizione dell’acrosina per la digestione della zona pellucida.
Fusione dei Gameti
Avviene attraverso la fusione delle membrane
plasmatiche di ovocita e spermatozoo. L’ingresso
del pronucleo maschile provoca lo sblocco della
meiosi dell’ovocita che emette i globuli polari
(cane, cavallo e volpe hanno il blocco meiotico in
metafase I, gli altri Mammiferi in metafase II). Lo
spermatozoo porta all’interno dell’ooplasma
mitocondri ed un centriolo; i primi degenerano
mentre il secondo diventa il centro organizzatore
dell’aster; nel citoplasma dello zigote saranno
quindi presenti solo i mitocondri dell’ovocita.
Una volta formato il pronucleo femminile il
materiale genetico si porta in piastra equatoriale e
si fonde. Si forma così lo zigote, il nuovo
individuo.
Globuli polari
Pronucleo maschile
Pronucleo femminile
Coda in degenerazione
Membrana di
fecondazione
Stadio dei Pronuclei
Fusione dei Pronuclei
Zigote
Il blocco della polispermia si distingue in due fasi denominate
blocco rapido e blocco lento.
Blocco della Polispermia
Il blocco rapido è determinato da un’onda di
depolarizzazione della membrana dell’ovocita che, dal
punto di ingresso del primo spermatozoo, si propaga su
tutta la superficie. L’ingresso del primo spermatozoo
favorisce l’ingresso di ioni, in particolare Na+. Questi
innalzano il potenziale della membrana (intorno a -70mV in
fase di riposo) fino a 20mV, determinando un blocco
nell’ingresso degli spermatozoi che non possono fondersi
con membrane che abbiano un potenziale superiore a 10mV. Questo blocco è istantaneo ma temporaneo poiché
le pompe ioniche presenti sulla membrana in breve
ripristinano il potenziale di riposo.
Il blocco lento è legato all’ingresso di ioni Ca++, che
induce il rilascio del contenuto dei granuli corticali. Questi
granuli, derivati dall’apparato di Golgi, sono posti sotto la
membrana
dell’ovocita,
contengono
materiale
mucopolisaccaridico ed enzimi proteolitici, e sono
considerati omologhi dell’acrosoma. Il rilascio dei granuli
corticali consiste nella fusione della loro membrana con la
membrana plasmatica e nella conseguente esocitosi del
loro contenuto. Gli enzimi dei granuli corticali distruggono i
recettori di membrana specie-specifici per gli spermatozoi,
mentre
i
mucopolisaccaridi,
richiamando
acqua,
induriscono ed ispessiscono la zona pellucida che viene
indicata come membrana di fecondazione. Questi
cambiamenti costituiscono il blocco lento della polispermia,
con carattere permanente. Esso inizia dopo 20 secondi e
termina entro 1 minuto dall’ingresso del primo
spermatozoo.
Zona pellucida
Scoppio dei granuli corticali
Membrana di fecondazione
Attivazione del metabolismo dell’uovo
Si distinguono
reazioni precoci (rappresentate dagli eventi che portano al blocco della polispermia e
alla rimozione del blocco della meiosi)
reazioni tardive (che comprendono l’attivazione della sintesi di DNA, la prima divisione
mitotica, l’attivazione degli mRNA materni e la conseguente sintesi proteica)
Ripresa della meiosi
Corona radiata
Zona pellucida
Granuli corticali
Reazione acrosomiale
Membrana ooplasmatica
Scoppio dei granuli corticali
Ingresso dello spermatozoo
SEGMENTAZIONE
La segmentazione è l’evento che porta il nuovo individuo dalla condizione di
unicellularità alla condizione di pluricellularità, attraverso una serie di divisioni
mitotiche che si interrompono in genere quando viene ripristinato il rapporto
nucleo/citoplasmatico tipico della specie.
L’embrione entra in segmentazione allo stadio di zigote e al termine del
processo è divenuto una blastula formata da cellule chiamate blastomeri, che
nel caso dei Mammiferi sarà una blastocisti, mentre nel caso degli Uccelli sarà
una discoblastula.
La modalità attraverso cui avviene la segmentazione è legata alla quantità di
vitello presente nell’uovo. Uova con scarsa o moderata quantità di vitello
segmentano completamente (segmentazione oloblastica), mentre uova con
grande quantità di vitello segmentano parzialmente (segmentazione
meroblastica).
Nei Vertebrati si descrive uno sviluppo di tipo regolativo:
la perdita di alcuni blastomeri, durante la segmentazione, non altera lo
sviluppo embrionale poiché le restanti cellule possono compensare tale
perdita. Infatti, i primi blastomeri di un embrione hanno la capacità di formare
una blastula completa se opportunamente separati. Questa capacità
differenziativa, definita totipotenza, nei Mammiferi si estende allo stadio di
due, quattro od otto blastomeri e successivamente viene persa. Tuttavia
blastomeri isolati in una fase successiva e reintrodotti in blastocisti ospiti
possono dare origine a tutti i tessuti embrionali, ma non a quelli
extraembrionali, indicando una riduzione della capacità differenziativa che
viene ora definita pluripotenza.
Introduciamo a questo punto il concetto di “potenza” di una cellula per indicare l’intera gamma di
strutture o tipi cellulari a cui essa può dare origine se esposta a differenti ambienti (in vivo o in vitro) ed il
concetto di staminalità o di cellula staminale per indicare cellule che:
si possono dividere illimitatamente,
sono morfologicamente indifferenziate,
la cui progenie comprende sia cellule identiche sia una discendenza cellulare destinata a differenziarsi.
Naturalmente questa definizione di staminalità deve tener conto della potenzialità differenziativa di una
cellula che è diversa tra una cellula staminale embrionale ed una cellula staminale adulta presente in un
tessuto a rinnovamento continuo (es. epidermide).
Secondo una recente classificazione possiamo definire almeno quattro classi di potenza in vivo:
a) totipotenza: indica le cellule capaci di originare sia l’embrione sia l’extraembrione (nei Mammiferi i
blastomeri sono totipotenti fino allo stadio di otto cellule);
b) pluripotenza: indica le cellule capaci di originare tutti e tre i foglietti embrionali (es. le cellule della
massa cellulare interna);
c) multipotenza: indica le cellule che possono originare solo una certa classe o categoria di cellule
correlate (es. cellule embrionali già determinate come appartenenti ad un foglietto che originano tutti i tipi
derivati del foglietto stesso; o nell’adulto le cellule emopoietiche del midollo osseo, capaci di originare
diversi tipi cellulari del sangue o le cellule adipose che originano fibro- e condro- blasti);
d) unipotenza: indica le cellule già determinate in un’unica direzione cellulare (un esempio di questo tipo
sono le cellule satelliti del muscolo striato che rimangono quiescenti per lungo tempo e sono riattivate,
dopo un evento traumatico o degenerativo, per fornire unicamente nuovi mioblasti che si fonderanno per
dare miotubi; la staminalità di queste cellule sta nel fatto che sono capaci di auto-rinnovarsi oltre che di
fornire mioblasti).
La segmentazione è quindi un processo che conduce alla formazione di cellule con un elevato grado di
potenza (toti- e pluripotenti) che, se opportunamente separate, sono in grado di fornire cellule staminali
embrionali (il loro utilizzo futuro, in campo umano come in campo veterinario, dipenderà dalle commissioni
bioetiche di ogni nazione).
Segmentazione
TOTALE
(uova oloblastiche)
Simmetria della
Segmentazione
Quantità e
posizione del tuorlo
Animali
Mammiferi
Rotazionale
Alecitiche
Radiale
Oligolecitiche,
Isolecitiche
Anfiosso
Radiale
Mesolecitiche,
Telolecitiche
Anfibi
Discoidale
Macrolecitiche,
Telolecitiche
Pesci, Rettili, Uccelli
e Mammiferi
Macrolecitiche,
Centrolecitiche
Insetti
PARZIALE
(Marsupiali e Placentati)
(Monotremi)
(uova meroblastiche)
Superficiale
Classificazione dei tipi di segmentazione in diverse specie animali
Il tuorlo è un adattamento evolutivo dell’embrione che pertanto può svilupparsi
senza assunzione esterna di cibo.
Gli animali che hanno uova con poco tuorlo (oligolecitiche), come il riccio di mare
(Echinoderma) o l’Anfiosso (Cordato), hanno uno sviluppo embrionale molto breve
con la formazione di una larva in grado di muoversi e nutrirsi.
Passando ai Vertebrati, dagli Anfibi ai Pesci e poi ai Rettili, si osserva un
progressivo aumento di tuorlo che si accompagna alla riduzione e poi alla
scomparsa dello stadio larvale. Infatti, alcuni Anfibi con uova contenenti una
discreta quantità di tuorlo (mesolecitiche), presentano segmentazione totale e
formano un girino come stadio larvale; altri Anfibi adattati alle zone desertiche, con
uova contenenti granda quantità di tuorlo (macrolecitiche) come quelle dei Pesci e
dei Sauropsidi, hanno segmentazione parziale e l’embrione si può sviluppare
interamente a spese del tuorlo. Per contro i Mammiferi Placentati, con uova prive
di tuorlo (alecitiche), hanno segmentazione totale ma sviluppano, accanto
all’embrione, la placenta, un annesso che, attraverso scambi materno-fetali,
provvede al nutrimento dello stesso.
L’ornitorinco depone le uova (1-3) in una tana
nella quale dopo la schiusa i piccoli
permangono per circa due settimane allattati
dalla madre
L’echidna trattiene l’uovo nell’utero fino
allo stadio di morula e successivamente
viene deposto e sistemato in una tasca
cutanea dove dopo la schiusa, il piccolo
viene allattato
I Mammiferi sono considerati filogeneticamente derivati da antenati correlati ai
Rettili primitivi che avevano probabilmente uova macrolecitiche simili a quelle
degli attuali Sauropsidi. Durante la filogenesi i Mammiferi sono diventati vivipari
in quanto la madre forniva nutrimento all’embrione attraverso la placenta. Non
essendoci necessità di tuorlo ai fini nutrizionali, questo è progressivamente
scomparso e le uova sono divenute più piccole. Questo aspetto dello sviluppo
evolutivo può facilmente essere evidenziato nelle tre sottoclassi dei Mammiferi:
Prototeri o Monotremi
Metateri o Marsupiali
Euteri o Placentati
Infatti i Prototeri depongono uova macrolecitiche ed il tuorlo costituisce il
nutrimento dell’embrione. I Metateri sviluppano uova con moderato contenuto di
tuorlo; quest’ultimo viene però espulso per cui il nutrimento dell’embrione è
sostenuto, in utero, da una pseudo-placenta. Infine gli Euteri hanno uova prive di
tuorlo ed il nutrimento dell’embrione in utero è sostenuto esclusivamente dalla
placenta.
In parallelo alla diminuzione del tuorlo, si assiste al passaggio da una
segmentazione parziale ad una segmentazione totale.
Segmentazione meroblastica discoidale
blastoderma
Cavità
sottogerminale
tuorlo
La segmentazione parziale (meroblastica) discoidale è caratteristica della
maggior parte dei Pesci, Rettili, Uccelli e Mammiferi Monotremi. Nelle
uova di queste specie il citoplasma è limitato al disco germinativo, La
segmentazione è definita parziale discoidale in quanto interessa solo il
disco germinativo, una sottile area al polo animale, a forma di calotta
sferica, in diretto contatto con la massa del tuorlo che occupa il resto
della cellula (uova macrolecitiche, telolecitiche) e che nel pollo ha un
diametro di circa 3mm.
La prima divisione è appena una fessura perpendicolare al centro del
disco germinativo; la seconda, perpendicolare alla prima, è ancora
limitata al centro del disco e porta alla formazione di quattro cellule
continue col tuorlo sia nella parte marginale che nella parte inferiore,
dette merociti.
Col progredire della segmentazione, sempre secondo piani meridiani, si
formano cellule centrali separate lateralmente dalle cellule marginali ma
tutte ancora in continuità col tuorlo sottostante. I primi veri blastomeri,
completamente separati dal tuorlo, si formano o per convergenza e
fusione dei solchi di segmentazione nella porzione citoplasmatica
sottonucleare dei merociti o per divisione cellulare con solchi di
segmentazione tangenziali. In tal caso le cellule più superficiali formano i
veri blastomeri, mentre le cellule inferiori mantengono la continuità col
tuorlo. Inoltre le cellule marginali, anch’esse in connessione col tuorlo,
formano una struttura sinciziale detta periblasto che ha la funzione di
demolire il tuorlo. Alla fine della segmentazione si ottiene pertanto una
blastula discoidale detta discoblastula in cui il blastoderma superficiale
è separato dal tuorlo da una cavità detta cavità sottogerminale. La
presenza di tale cavità permette di identificare nel blastodisco un’area
centrale più chiara detta area pellucida, composta da blastomeri ben
definiti posti sulla cavità sottogerminale e un’area marginale più scura
detta area opaca, composta da merociti in continuità col tuorlo. Al
momento della deposizione gruppi di cellule del blastoderma possono
diffondere nella cavità sottogerminale dove si riuniscono a formare un
foglietto continuo detto ipoblasto, mentre le cellule rimaste in superficie
formano l’epiblasto. Lo spazio delimitato da questi due foglietti
corrisponde al blastocele.
Segmentazione oloblastica rotazionale
Nei Mammiferi (Placentati e Marsupiali) la segmentazione è oloblastica (totale) rotazionale in quanto i piani di
segmentazione della seconda divisione sono ortogonali tra loro. A questa particolarità si unisce il fatto che le divisioni
sono molto lente ed asincrone cosicché l’embrione può risultare formato da un numero dispari di blastomeri. Inoltre,
si ha la formazione di uno stadio intermedio nella segmentazione dei Mammiferi, in cui l’embrione è chiamato
morula, in quanto i suoi blastomeri si compattano grazie allo stabilirsi di contatti intercellulari mediati da una
molecola di adesione calcio-dipendente chiamata E-caderina (detta anche ovomorulina), ciò conferisce all’embrione
l’aspetto di una mora.
I blastomeri secernono un liquido che si raccoglie in una cavità detta blastocele la cui formazione provoca lo
spostamento di una parte delle cellule da un lato dell’embrione che è ora detto blastocisti. A questo stadio
l’embrione perde la membrana di fecondazione (schiusa) e si assiste ad un notevole mutamento di forma e
dimensioni della blastocisti.
Nella blastocisti le cellule non sono tutte uguali, quelle spostate da un lato sono più grandi ed unite da giunzioni
comunicanti, formano il disco embrionale (massa cellulare interna, nodo embrionale) dal quale deriverà l’embrione;
quelle alla periferia sono più piccole, numerose e unite da giunzioni occludenti, formano il trofoblasto, mediano gli
scambi trofici ed in seguito entreranno nella formazione della placenta.
Annidamento della blastocisti
Mentre subisce il processo di segmentazione, l’embrione viene trasportato lungo le tube uterine mediante il movimento
delle ciglia dell’epitelio tubarico e la contrazione della componente muscolare della parete tubarica. In genere, esso impiega
un tempo varabile nelle diverse specie per raggiungere l’utero.
Durante il percorso tubarico la membrana di fecondazione derivata dalla zona pellucida, priva di cariche elettriche ed
immunologicamente inerte, protegge l’embrione da un’eventuale risposta immunitaria materna.
La membrana di fecondazione, inoltre, impedisce l’impianto della blastocisti nelle tube uterine.
Una volta raggiunto l’utero, la blastocisti sguscia dalla membrana di fecondazione e resta libera per un breve periodo nel
lume dell’utero. In questo periodo essa assorbe liquidi dalla cavità uterina ed aumenta di volume. Successivamente le
cellule del trofoblasto, poste sul polo in cui ha sede l’embrioblasto, prendono rapporto con la mucosa uterina ed incomincia
l’impianto o annidamento della blastocisti in utero.
Uomo
Cavallo
Asino
Tempo in g impiegato Bovino
dall’embrione per giungere Bufalo
in utero
Ovino
Caprino
Suino
Cane
3
8-10
8-10
3
3
3
3
3-4
8-10
L’impianto o annidamento può essere di vario tipo:
-impianto centrale in cui la blastocisti rimane al centro della
cavità uterina,
-impianto eccentrico, in cui la blastocisti viene accolta in un
diverticolo dell’endometrio ed è quasi totalmente avvolta dalla
mucosa uterina,
-impianto interstiziale in cui la blastocisti penetra nello
spessore dell’endometrio che viene eroso dagli enzimi delle
cellule trofoblastiche.
GASTRULAZIONE
In tutti i Vertebrati al termine della segmentazione si forma una blastula, più o meno cava e colma di liquido circondata
da cellule organizzate in un foglietto detto blastoderma. Tali cellule vanno incontro ad una serie di spostamenti, correlati
a modificazioni morfologiche, che comportano anche un profondo cambiamento della forma della blastula.
La gastrulazione è il processo che porta al riordino delle cellule del disco embrionale prima in due
(stadio didermico) e poi in tre (stadio tridermico) strati concentrici denominati foglietti germinativi che
dall’esterno all’interno sono rappresentati da ectoderma, mesoderma ed endoderma, dai quali
deriveranno tutti i tessuti e gli organi dell’individuo.
Il riordino delle cellule avviene attraverso movimenti morfogenetici irreversibili ed il processo non comporta in genere
aumento volumetrico dell’embrione, anche se l’attività mitotica è comunque intensa. Si assiste però a modificazioni
metaboliche in relazione all’iniziale differenziamento.
Sebbene i tipi di gastrulazione varino notevolmente nelle diverse specie animali, anche in relazione al tipo di blastula
che si è formata alla fine del processo di segmentazione, i meccanismi primari di tale processo sono conservati
filogeneticamente e sono ripetutamente utilizzati in varie tappe dello sviluppo.
La gastrulazione è il risultato di una combinazione dei seguenti meccanismi di movimento:
1. espansione di cellule in superficie in modo da ricoprire e circondare quelle più interne o epibolia;
2. movimento di divisione di uno strato di cellule in due lamine parallele o delaminazione;
3. movimenti di aggregazione cellulare o di addensamento conseguenti ad aumento di adesione tra le
cellule o ad aumento locale delle divisioni mitotiche.
4. spostamento di cellule dalla superficie verso l’interno.
Questo spostamento di cellule verso l’interno può avvenire per:
• invaginazione, ripiegamento di una porzione del blastoderma o di una lamina cellulare verso
l’interno, simile all’infossamento di una palla di gomma quando viene schiacciata in un punto
circoscritto o embolia;
• involuzione, ripiegamento e scorrimento di cellule in modo da accollarsi alla superficie interna dello
strato superficiale;
• ingressione o immigrazione, migrazione di singole cellule del blastoderma all’interno dell’embrione.
Anche se l’uovo degli Uccelli e quello dei Mammiferi sono molto diversi tra loro, di fatto le due blastule che derivano dalle
rispettive segmentazioni si assomigliano abbastanza.
Infatti in entrambi i casi abbiamo un embrione biconvesso che sovrasta una cavità, nel caso degli uccelli piena di tuorlo
chiamata cavità vitellina, nel caso dei mammiferi piena di liquido chiamata blastocele o cavità della blastocisti.
In realtà le due gastrulazioni sono quasi uguali per cui descriveremo solo quella dei Mammiferi.
Il passaggio allo stadio didermico consiste in una delaminazione delle cellule del disco embrionale verso
la cavità della blastocisti dove va a costituire il foglietto endodermico (endoblasto o ipoblasto). La massa
cellulare compresa tra trofoblasto ed endoderma diviene l’ectoderma (ectoblasto o epiblasto).
In seguito il foglietto endodermico si espande verso il trofoblasto rivestendone la superficie interna, in
questo modo il blastocele diviene cavità vitellina.
Prima della formazione del terzo foglietto si ha la formazione della cavità amniotica attraverso modalità
diverse in relazione alla specie.
La “ex-blastocisti” a questo stadio presenta un’area embrionale didermica interposta tra cavità amniotica
e cavità vitellina; parliamo ora di gastrula iniziale.
La gastrulazione continua con la comparsa di
un ispessimento di cellule che in posizione
dorso-caudale danno luogo ad una struttura
chiamata linea primitiva, la quale termina con
un infossamento arrotondato detto nodo di
Hensen. Le cellule che si ammassano a livello
della linea primitiva migrano attraverso il nodo
di Hensen e si insinuano tra ectoderma ed
endoderma andando a formare il mesoderma.
Le future cellule mesodermiche migrano in direzione laterale,
cranio-laterale, caudale e cefalica. La migrazione in direzione
cefalica si evidenzia con la comparsa di un ispessimento,
davanti al nodo di Hensen e sotto l’ectoderma, chiamato
processo cefalico o notocordale. Il mesoderma non si
frappone tra ectoderma ed endoderma a livello delle future
membrane bucco-faringea e cloacale.
Entro 2 settimane la migrazione mesodermica ha interessato tutta l’area
embrionale e comincia ad invadere quella extraembrionale con le due
lamine di mesoderma laterale.
La linea primitiva ad un certo punto regredisce ed il processo
cefalico viene esteso in direzione cefalo-caudale ed evolve in
notocorda definitiva solida, posta dorso-medialmente tra
ectoderma ed endoderma con a lato le due bande longitudinali
di mesoderma. Il nodo di Hensen indietreggia e si localizza,
nei deuterostomi, a livello della regione anale.
Negli spazi tra i tre foglietti si insinua un tessuto interstiziale di origine
mesodermica chiamato mesenchima, il quale rappresenta la matrice per
tutti i tessuti connettivi.
Prescindendo da situazioni particolari, nei Mammiferi
Placentati si possono riconoscere diverse modalità di
amniogenesi che sono da ricondurre al diverso rapporto tra
trofoblasto e MCI:
– nei Carnivori, nel maiale e nel coniglio il trofoblasto al di
sopra della MCI si atrofizza, le cellule dell’epiblasto si
appiattiscono al di sopra dell’ipoblasto allineandosi col
trofoblasto marginale e quindi si sollevano le pieghe
amniotiche (amniogenesi per pieghe).
– nei Primati, nel riccio e nel pipistrello la MCI rimane
circondata superiormente dal trofoblasto che continua a
proliferare con un serie di divisioni nucleari non seguite da
citodieresi. Questo processo comporta un ispessimento ed
un differenziamento del trofoblasto in citotrofoblasto e
sinciziotrofoblasto. Tale ispessimento impedisce l’apertura in
superficie della cavità che si viene a costituire per deiscenza
tra le cellule dell’epiblasto. Questa cavità diventa quindi
cavità amniotica definitiva (amniogenesi per cavitazione o
schizocelia). Le cellule che ne delimitano la parte profonda
costituiscono l’epiblasto embrionale mentre quelle della
porzione laterale e superficiale costituiscono il foglietto
epiblastico dell’amnios.
– negli Ungulati (cavallo, Ruminanti), tra le cellule
dell’epiblasto sormontate dal trofoblasto, si formano delle
piccole fessure che poi confluiscono in un’unica cavità.
Questa è detta cavità amniotica primitiva in quanto è
delimitata da cellule dell’epiblasto. Successivamente la
parete superiore di questa cavità, insieme al trofoblasto, va in
lisi ed i margini liberi della porzione rimasta si appiattiscono
in modo da formare il disco epiblastico allineato col
trofoblasto. A questo punto al limite tra trofoblasto ed
epiblasto si sollevano le pieghe amniotiche (amniogenesi di
transizione).
MAPPE PROSPETTICHE O DEI TERRITORI PRESUNTIVI
L’osservazione del succedersi degli eventi formativi negli embrioni ha permesso agli scienziati, che si occupavano di
embriologia descrittiva, di tracciare il destino di linee cellulari a partire dalle prime fasi di segmentazione.
Inoltre, l’embriologia sperimentale, mediante raffinate metodiche di marcatura cellulare, ha consentito di seguire il
destino delle cellule a mano a mano che l’embrione si differenzia nei vari tessuti.
Più recentemente, la biologia dello sviluppo ha studiato l’espressione di geni regolatori del differenziamento e
caratterizzato cellule, ancora prima che il loro fenotipo potesse essere osservato al microscopio.
Tutti questi studi hanno permesso di disegnare, per ogni specie studiata, delle vere e proprie mappe
dei territori embrionali in via di sviluppo.
Le rappresentazioni schematiche di territori embrionali precedenti la gastrulazione, che origineranno i
tre foglietti primordiali con le strutture embrionali annesse vengono indicate, con il termine di mappe
prospettiche o presuntive. Il confronto tra le mappe permette di evidenziare che, anche in animali
profondamente diversi, nell’embrione il rapporto tra i territori che daranno origine ai tre foglietti è
spesso molto simile. Per esempio, la notocorda occupa sempre una posizione medio-dorsale ed il
territorio che originerà il sistema nervoso (neuroectoderma) è posto sempre anteriormente ad essa. Il
territorio che originerà l’ectoderma di superficie (epidermide) circonda il neuroectoderma sempre nella
stessa posizione.
Ectoderma di
superficie
ANFIOSSO
Neuroectoderma
RANA
Notocorda
PESCE ZEBRA
Endoderma
Mesoderma
POLLO
Mesoderma
extraembrionale
TOPO
Nello schema sono rappresentate le mappe presuntive di diversi Cordati, indicanti il destino delle diverse regioni
dell’embrione all’inizio della gastrulazione.
Differenziamento dei foglietti embrionali
Ectoderma
E’ il foglietto embrionale più
esterno, ha la forma di un disco
epiteliale in continuità con
l’ectoderma amniotico e si divide
in tre territori principali:
l’ectoderma di rivestimento
il tubo neurale
la cresta neurale
La notocorda induce l’ectoderma soprastante (neuroectoderma) ad
ispessirsi per formare il disco neurale la cui evoluzione
(neurulazione) porta alla formazione del sistema nervoso.
Il disco neurale si in vagina formando la doccia neurale i cui cercini
chiudendosi formano il tubo neurale definitivo che darà origine al
sistema nervoso centrale (encefalo e midollo spinale). Inizialmente il
tubo neurale è in rapporto col sacco amniotico attraverso i neuropori
anteriore e posteriore, che si chiuderanno in seguito.
Dal punto di saldatura dei cercini neurali si forma la cresta neurale,
struttura transitoria da cui derivano il sistema nervoso periferico, la
midollare del surrene, i melanociti, le componenti scheletricoconnettivali della testa.
Dal restante ectoderma (di rivestimento), derivano l’epidermide e i
derivati cutanei (peli, piume, unghie, artigli, ecc.), le creste dentarie,
l’ipofisi anteriore, i placodi. Infatti alla chiusura completa del tubo
neurale, da esso originano paia di vescicole (otiche, ottiche,
olfattorie) che si portano verso l’ectoderma epidermico, inducendo la
formazione dei:
placodi otici, dai quali originano i labirinti membranosi dell’orecchio
interno,
placodi del cristallino, che daranno i cristallini degli occhi,
placodi olfattori, che daranno le fosse nasali e la mucosa olfattoria,
placodi epibranchiali, che daranno neuroni sensitivi.
Con il termine di induzione si intende il fenomeno per cui un particolare
agente (induttore) fa nascere in un altro corpo prossimo (indotto) delle
proprietà che altrimenti non si realizzerebbero.
Un
induttore
embrionale
esiste
in
quanto
esistono
contemporaneamente uno o più sistemi reagenti o bersaglio suscettibili
o competenti a reagire all’induttore stesso.
La competenza al fattore induttore è in genere limitata ad uno stadio
specifico dello sviluppo del sistema competente.
Il cordomesoderma induce il neuroectoderma a formare il tubo neurale,
questo una volta chiuso origina le vescicole ottiche che inducono la
formazione dei placodi del cristallino.
Una struttura indotta (disco neurale) diventa quindi in questo caso un
induttore.
Il cordomesoderma viene definito induttore primario.
La vescicola ottica è un induttore secondario.
Derivati
dell’Ectoderma
Mesoderma
E’
il
foglietto
embrionale
intermedio e si divide in cinque
territori principali:
Placca precordale
Cordomesoderma
Mesoderma parassiale
Mesoderma intermedio
Mesoderma laterale
Cavità amniotica
Ectoderma
Mesoderma
Endoderma
Placca notocordale
Ectoderma
Placca notocordale
Endoderma
Mesoderma parassiale
Mesoderma
somatopleurico
Mesoderma intermedio
Mesoderma
splancnopleurico
Mesoderma laterale
Processo
notocordale
Mesoderma parassiale
Mesoderma intermedio
Mesoderma laterale
Immediatamente a lato della notocorda si formano, per proliferazione cellulare, 2 bande longitudinali di
mesoderma parassiale in direzione cefalo-caudale. Lateralmente a questo il mesoderma rimane più
sottile e viene diviso in intermedio e laterale.
Con l’inizio della regressione della linea primitiva il
mesoderma parassiale si metamerizza formando paia di
somiti.
Le cellule della parte ventro-mediale dei somiti,
inizialmente epitelioidi, entrano in mitosi e diventano
mesenchimatiche, migrano lateralmente alla notocorda a
formare lo sclerotomo le cui cellule si differenzieranno in
condroblasti ed osteoblasti.
La parte dorso-laterale del somite si divide in:
-miotomo dal quale originerà la muscolatura scheletrica,
Il mesoderma intermedio è metamerico
nelle parti cervicale e toracica
(nefrotomi) mentre più caudalmente è
compatto (blastema metanefrogeno).
Dal mesoderma intermedio derivano
pronefro, mesonefro e metanefro, dotti
genitali, tessuto interstiziale delle
gonadi (cellule della teca, cellule del
Leydig), corticale del surrene.
-dermatomo che formerà derma e tessuto sottocutaneo,
-Sindetomo che darà tendini e legamenti.
Il mesoderma laterale si delamina in due foglietti:
-mesoderma viscerale o splancnopleura
-mesoderma parietale o somatopleura.
Nell’area extraembrionale questi due foglietti forniscono la componente mesodermica rispettivamente al
sacco vitellino e all’allantoide e ad amnios e corion. Tra i due foglietti si trova invece delimitato il celoma;
la parte intraembrionale di questo forma le cavità peritoneale, pleuriche e pericardica.
Dal mesoderma laterale deriva anche il sistema cardio-circolatorio.
Derivati del
Mesoderma
Endoderma
L’endoderma
comincia
il
suo
differenziamento formando il tubo
intestinale primitivo collegato nella
sua parte media con il sacco vitellino
attraverso il dotto onfalomesenterico.
Anteriormente l’intestino è delimitato
dalla membrana bucco-faringea e
posteriormente da quella cloacale.
La porzione posteriore dell’intestino
è in comunicazione con l’ allantoide.
Oltre al rivestimento epiteliale ed alle
ghiandole del tubo intestinale, dal
foglietto endotermico derivano anche
l’epitelio e le ghiandole delle vie
aeree inferiori e del polmone, il
rivestimento epiteliale della vescica
e dell’uretra, della cavità timpanica e
della tromba di Eustachio.
Di origine endotermica sono anche
tonsille, tiroide, paratiroidi e timo.
Endoderma
L’endoderma comincia
il suo differenziamento
formando
il
tubo
intestinale primitivo.
(fegato, pancreas)
Durante lo sviluppo dei Mammiferi si succedono due
periodi indicati come periodo embrionale e periodo
fetale.
Il periodo embrionale va dalla fecondazione al
momento in cui il nuovo organismo è nettamente
definito nel suo aspetto esteriore e nelle sue parti
costitutive.
A questo segue il periodo fetale, che si estende fino
al termine della gravidanza, nel quale si perfeziona e
si completa l’organogenesi.
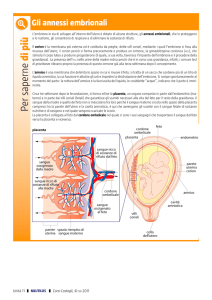
ANNESSI EMBRIONALI: aspetti evolutivi
Gli annessi embrionali sono strutture che si formano da estensioni dei foglietti embrionali nella
cosiddetta area extraembrionale e permettono lo sviluppo dell’embrione e del feto.
Nei Vertebrati acquatici a fecondazione esterna, come la maggior parte dei Pesci Teleostei e degli
Anfibi, lo sviluppo embrionale avviene all’interno della membrana dell’uovo circondata da un esile
strato gelatinoso, cosicchè gli scambi gassosi e l’allontanamento dei cataboliti avvengono facilmente
tra l’embrione e l’acqua circostante. Il tuorlo, necessario alla nutrizione dell’embrione, viene
inglobato nel citoplasma delle cellule dell’endoderma (Anfibi) o avvolto dai tre foglietti endoderma,
mesoderma e ectoderma (Teleostei) che insieme formano la parete del cosiddetto sacco del tuorlo o
vitellino. Questo costituisce l’unico annesso presente nei Pesci.
L’espansione dei Vertebrati sulle terre emerse ha richiesto, tra i tanti adattamenti, che gli embrioni
fossero particolarmente protetti dalla disidratazione e dalla forza di gravità. E’ stato quindi ricreato
un ambiente acquatico dove gli embrioni potessero svilupparsi: l’amnios con il liquido amniotico in
esso contenuto. Pertanto i Rettili, gli Uccelli e i Mammiferi possiedono questo nuovo annesso e
sono definiti amnioti.
Inoltre i Vertebrati terrestri ovipari (la maggior parte dei Sauropsidi e i Mammiferi Monotremi) hanno
sviluppato degli involucri a funzione protettiva e nutritizia attorno alle uova, quali l’albume, la
membrana testacea e il guscio calcareo. Tali involucri, rendendo difficoltosa l’eliminazione delle
sostanze di scarto e la respirazione, hanno determinato la necessità della formazione di altri due
annessi embrionali: il corion e l’allantoide.
La viviparità, che comporta l’instaurarsi di un rapporto tra l’ovidutto o l’utero materno e l’embrione, è
sporadica nei Vertebrati acquatici (alcuni squali) e nei Rettili (camaleonti), ma raggiunge il successo
evolutivo nei Mammiferi Metateri e Euteri, con rapporti sempre più stretti tra madre ed embrione. Tali
rapporti sono resi possibili da profonde modificazioni del corion e della mucosa uterina che insieme
costituiscono la placenta, organo che attua i necessari scambi metabolici tra madre ed embrione
rendendo pertanto inutile la presenza di tuorlo nelle uova di questi animali.
Formazione degli annessi embrionali negli Uccelli
I foglietti embrionali che si sono differenziati nell’embrioblasto si espandono a circondare il tuorlo, che però al polo
distale rimane in contatto con l’albume.
Il mesoderma laterale si delamina in
-splancnopleura che aderisce all’endoderma
-somatopleura che aderisce all’ectoderma.
Il sacco vitellino è l’avvolgimento extraembrionale più interno, è a diretto contatto con il tuorlo ed è formato da
endoderma e splancnopleura.
La delimitazione del corpo dell’embrione porta alla separazione dell’intestino primitivo dal sacco vitellino che
rimangono però in comunicazione attraverso il dotto onfalomesenterico o canale vitellino.
L’endoderma del sacco vitellino si solleva in pieghe in modo da ampliare la superficie di contatto con il
deutoplasma che viene digerito da enzimi secreti dalle cellule endodermiche, gli intermedi vengono riassorbiti e
trasferiti all’embrione attraverso i vasi vitellini che passano nel dotto onfalomesenterico. Anche l’albume viene
riassorbito in questo modo.
A livello del sacco vitellino si hanno i primi fenomeni emopoietici. Man mano che il tuorlo viene utilizzato il sacco
vitellino si riduce a nel pollo al 19° giorno di incubazione è completamente riassorbito.
Con l’approfondarsi dell’embrione si sollevano, nell’area pellucida extraembrionale, 4 pieghe amniotiche (craniale,
caudale e laterali) che si chiudono sopra l’embrione costituendogli attorno 2 involucri: corion più esterno ed amnios
più interno.
L’amnios racchiude la cavità amniotica ripiena di liquido amniotico prodotto in parte dalle cellule del sacco stesso
ed in parte proveniente dall’albume. Il liquido amniotico viene utilizzato dall’embrione.
Il corion o sierosa di Von Baer si estende
distalmente verso il sacco vitellino, al polo
acuto dell’uovo forma una espansione che
circonda l’albume (sacco dell’albume).
Tra il mesoderma che tappezza internamente il
corion e quello che tappezza esternamente
l’amnios e il sacco vitellino è compreso il
celoma extraembrionale.
L’allantoide è una estroflessione caudoventrale
dell’intestino primitivo che nel pollo compare
alla 60a ora di incubazione. Presenta la stessa
costituzione del sacco vitellino.
Espandendosi l’allantoide entra in contatto con
il corion formando l’allanto-corion all’interno del
quale si sviluppano i vasi allantoidei che
entrano in comunicazione con i vasi
intraembrionali.
L’allantoide attraverso il corion è in contatto con
il guscio calcareo dal quale preleva ioni Ca++
utili all’ossificazione dell’embrione, presiede gli
scambi gassosi e raccoglie i cataboliti. Negli
ultimi due giorni di incubazione regredisce e si
essicca e alla schiusa rimane aderente al
guscio.
Formazione degli annessi embrionali nei Mammiferi
Il sacco vitellino nei Mammiferi placentati va incontro
a rapida involuzione a meno che non vada a formare
l’onfalo-corion. Riveste comunque notevole importanza
in quanto costituisce la sede iniziale dell’emopoiesi ed
è a suo carico che si forma la linea germinale. A
differenza degli Uccelli il sacco è completamente
chiuso e deriva dalla cavità della blastocisti che
rivestita da endoderma diviene la cavità vitellina.
La parte distale della blastocisti non viene raggiunta
dal mesoderma extraembrionale in tutte le specie per
cui il sacco vitellino nella sua parte distale può o meno
contrarre rapporti con il corion.
Indipendentemente dalla modalità di formazione o dal
grado di sviluppo o di involuzione, con il sollevamento
dell’embrione, il sacco vitellino si delimita dall’intestino
primitivo a cui resta legato tramite il peduncolo
vitellino.
Nei Primati il sacco vitellino è poco sviluppato ed
occupa solo in parte la cavità della blastocisti che
viene invasa da cellule mesenchimali. Il mesenchima
si addensa poi contro la parete degli annessi
(endoderma
vitellino,
ectoderma
amniotico,
citotrofoblasto) e si delimita così il celoma
extraembrionale.
L’amnios è un sacco completamente chiuso che
circonda l’embrione. Contiene il liquido amniotico
derivato da accumulo di urina fetale, da secrezioni
ghiandolari, da interscambi tra circolazione materna e
fetale. Il liquido amniotico è continuamente rinnovato
e contiene
-alti livelli di Na+, Cl-, fruttosio
-bassi livelli di K+, Mg++, glucosio, creatinina, acido
urico, urea
-cellule desquamate, ferro, enzimi.
Il liquido amniotico ha diverse funzioni
-idrata, protegge e facilita i movimenti del feto
-agevola la progressione del feto durante il parto
-i suoi enzimi proteolitici e lipolitici digeriscono
embrioni eventualmente morti nelle prime fasi della
gravidanza.
Nei Mammiferi l’allantoide compare più precocemente
che negli Uccelli, in forma di estroflessione caudale del
sacco vitellino. Il peduncolo allantoideo o uraco collega
l’allantoide all’intestino posteriore.
Questo annesso perde nei Mammiferi l’originaria
funzione di raccolta dei cataboliti embrionali, ma
assume importanza nelle specie in cui entra in contatto
con il corion formando l’allanto-corion che partecipa
alla formazione della placenta alla quale fornisce la
vascolarizzazione tramite le arterie e le vene
allantoidee o ombelicali. Negli Equidi si forma anche
l’allanto-amnios.
Il liquido allantoideo si forma per accumulo di urina
fetale e per secrezione delle cellule allantoidee. E’ un
ultrafiltrato contenente fruttosio, urea, acido urico,
creatinina, ioni.
L’allantoide stocca i cataboliti che non vengono
immediatamente eliminati dalla placenta, il suo grado
di sviluppo è in stretta relazione con il tipo di placenta
proprio di una determinata specie.
Il corion è l’annesso embrionale più esterno. La sua superficie sviluppa villosità che in alcune zone regrediscono
(chorion laeve) mentre permangono (chorion frondosum) in altre destinate a contrarre rapporti con la mucosa
uterina per la formazione della placenta.
I villi possono essere digitiformi, semplici o ramificati, o avere struttura laminare come nel suino. Nelle placente per
apposizione il villo è dato da epitelio cubico con asse mesenchimale vascolarizzato da vasi allantoidei. Nelle
placente endotelio-coriali, le cellule del trofoblasto si organizzano a formare il sinciziotrofoblasto con asse
mesenchimale, che forma lamelle anastomizzate. Nelle placente emo-coriali si ha la maggior complessità
strutturale dei villi. Nell’uomo i villi primari sono costituiti inizialmente da travate sinciziotrofoblastiche con all’interno
un asse citotrofoblastico. Questo villo diviene secondario quando (18° giorno) al suo interno compare un asse
mesenchimale ed il villo si ramifica. Dalla 4a settimana nel mesenchima compaiono i capillari che entrano in
rapporto con i vasi allantoidei ed i villi si dicono terziari.
Placenta
La placenta è una struttura complessa formata da una componente fetale (corion) ed
una componente materna (mucosa uterina). Ha il compito di provvedere agli scambi
materno-fetali per la nutrizione, la respirazione e l’escrezione dei cataboliti prima
dell’embrione e poi del feto, oltre ad avere un’importante funzione endocrina ed
immunologica.
Nella più semplice delle placente il corion si presenta liscio ed è semplicemente
poggiato sull’epitelio della mucosa uterina. Questa pseudo-placenta è propria dei
Metateri che sono detti aplacentati.
Nei Mammiferi Placentati le placente presentano notevoli differenze.
Per una loro descrizione e classificazione si possono seguire differenti criteri che
tengono conto:
1. dell’origine della vascolarizzazione del corion,
2. della distribuzione dei villi sul corion,
3. del rapporto tra corion e mucosa uterina,
4. del comportamento della mucosa uterina al momento del parto.
Origine della vascolarizzazione del corion
Prima della formazione della placenta l’embrione dei Mammiferi si nutre per istiotrofo o latte uterino, cioè
utilizzando i metaboliti contenuti nel liquido prodotto dalle ghiandole uterine.
Con la formazione della placenta si ha una nutrizione per emotrofo e l’assunzione dei metaboliti dalla
madre attraverso la circolazione placentare.
Quando, per l’assenza del celoma extraembrionale, la somatopleura del corion è fusa con la
splancnopleura del sacco vitellino, il corion viene vascolarizzato dalla splancnopleura del sacco vitellino.
In tal caso la placenta è definita vitellina o onfaloide. Questo tipo di placenta è presente oltre che nei
Metateri, nei primi stadi della gravidanza degli Equidi e per tutta la gravidanza dei Carnivori, nei quali è
coadiuvata da una placenta allantoidea.
Nelle specie in cui l’allantoide si espande nel celoma extraembrionale e si pone in contatto con la
somatopleura del corion, questo viene vascolarizzato dalla splancnopleura dell’allantoide per cui la
placenta è definita allantoidea. Questo tipo di placenta è presente in alcune specie di Metateri e nella
maggior parte degli Euteri.
Infine, in alcune specie (Primati e Roditori) l’allantoide è una struttura poco sviluppata, anche se i vasi
allantoidei continuano ad essere ben sviluppati ed a vascolarizzare il corion. In tal caso la placenta viene
definita anallantoidea.
Distribuzione dei villi sul corion
Sulla base della distribuzione dei villi sulla superficie
del corion la placenta può essere definita: diffusa,
multipla o cotiledonare, zonata e discoidale.
Nella placenta diffusa (A) (suino ed Equidi) i villi sono
situati su tutta la superficie del corion e affondano in
piccole nicchie della mucosa uterina.
Nella placenta multipla o cotiledonare (B) (Ruminanti)
i villi, raggruppati a ciuffi in piccole zone, costituiscono
i cotiledoni. Questi entrano in rapporto con la mucosa
uterina a livello di aree ben distinte che si chiamano
caruncole. Ogni cotiledone con la rispettiva caruncola
costituisce un placentoma. Il complesso dei cotiledoni
costituisce il chorion frondosum mentre le aree
intercotiledonari costituiscono il chorion laeve.
Nella placenta zonata (C) (Carnivori)i villi del chorion
frondosum sono disposti a formare un anello che
avvolge la fascia equatoriale del corion mentre le
estremità costituiscono il chorion laeve. In alcune
specie tale anello può essere incompleto.
Nella placenta discoidale o bidiscoidale (D) (Primati) il
chorion frondosum costituisce una zona circolare o
ovoidale che interessa un tratto relativamente ristretto
del corion.
A
B
A
C
D
La barriera materno-fetale inizialmente è costituita da tre strati
extraembrionali fetali (l’endotelio dei vasi, il connettivo
mesodermico e l’epitelio del corion) e da tre strati della
mucosa uterina (endotelio vasale, connettivo ed epitelio di
rivestimento).
Tale situazione si riscontra anche in stadi avanzati della
gravidanza solo nel suino e negli equini. In queste specie la
placenta viene definita epitelio-coriale (A) in quanto l’epitelio
del corion è a contatto con l’epitelio della mucosa uterina per
cui la barriera placentare è completa.
In altre specie, con l’avanzamento della gravidanza ed il
completamento dello sviluppo della placenta, si assiste ad una
erosione della mucosa uterina da parte dell’epitelio dei villi, in
modo da ridurre il numero degli strati della componente
materna.
Nella placenta dei Ruminanti i villi coriali erodono parzialmente
l’epitelio della mucosa uterina per cui l’epitelio del corion in
alcuni punti è in contatto col connettivo della mucosa uterina
(sindesmo- coriale). In tal caso la barriera placentare si riduce
dell’epitelio della componente materna. Secondo alcuni autori,
in alcuni Ruminanti cellule binucleate dell’epitelio dei villi
coriali si fondono con cellule dell’epitelio endometriale per cui
definiscono tale tipo di placenta sinepitelio-coriale (B).
Nei Carnivori i villi del corion erodono sia l’epitelio che il
connettivo della mucosa uterina, prendendo rapporto con
l’endotelio dei vasi materni. Tale tipo di placenta è definita
endotelio-coriale (C).
In Insettivori, Roditori, Lagomorfi e Primati i villi coriali hanno
eroso completamente gli strati materni per cui pescano in
lacune sanguigne localizzate nella mucosa uterina. Tale tipo di
placenta è definito emo-coriale (D).
Rapporto tra corion e mucosa uterina
Comportamento della mucosa uterina al momento del parto
Come è stato precedentemente descritto, la mucosa uterina (decidua) può
subire delle modificazioni per consentire l’impianto della blastocisti.
L’entità e l’ampiezza di tali modificazioni sono diverse nelle varie specie.
La placenta degli Equidi (epitelio-coriale) e quella dei Ruminanti (sindesmocoriale o sinepitelio-coriale) sono dette adeciduate in quanto la mucosa uterina
non ha subìto erosione da parte dei villi del corion per cui il distacco di questi
con il secondamento avviene senza perdite di sangue.
Nelle placente in cui è presente erosione (endotelio-coriale e emocoriale), il
distacco del corion con il secondamento può determinare perdite ematiche più
o meno imponenti seguite dall’espulsione della decidua. Tali placente in cui una
parte dell’endometrio viene eliminata al momento del parto, vengono definite
deciduate.
Tra le placente deciduate, l’erosione è meno profonda ma più ampia nella
placenta endotelio-coriale (Carnivori), mentre è più profonda e meno ampia
nella placenta emo-coriale (Primati).
Funzioni della Placenta
La placenta assicura gli scambi respiratori e metabolici tra madre e feto; essa inoltre ha funzione
di protezione impedendo, tramite la barriera placentare, il passaggio di sostanze o microrganismi
nocivi nella circolazione fetale.
Poiché in nessun caso il sangue fetale può mescolarsi al sangue materno, gli scambi sono resi possibili dalla
permeabilità selettiva del filtro placentare. Tale permeabilità dipende, oltre che dalla barriera placentare dalla
superficie totale di contatto tra le componenti materna e fetale e dalle caratteristiche della vascolarizzazione delle
due componenti.
In direzione materno-fetale attraversano la barriera placentare: ossigeno, acqua, elettroliti, sali minerali, protidi,
glucidi, lipidi, vitamine, ormoni. Talora possono passare anticorpi, farmaci, virus, batteri, parassiti.
In direzione fetale-materna attraversano la barriera placentare: anidride carbonica, acqua, cataboliti quali urea e
acido urico, elettroliti ed ormoni. Mentre il passaggio di sostanze gassose e di sali minerali avviene per diffusione, il
passaggio di macromolecole (proteine, lipidi, glucidi) richiede una scissione di queste ad opera dell’epitelio coriale
ed un successivo passaggio degli elementi di base nel sangue fetale.
Le placente deciduate (emo- ed endotelio-coriale) permettono inoltre il passaggio degli anticorpi in direzione
materno-fetale contribuendo ad una certa immunità dei neonati. Nelle placente adeciduate (epitelio- e sinepiteliocoriale) gli anticorpi non superano la barriera placentare ed i neonati possono acquisire anticorpi con l’assunzione
del colostro all’inizio dell’allattamento.
La placenta svolge inoltre un’importante funzione endocrina in quanto produce diversi ormoni:
-Gonadotropine corioniche, che stimolano la sintesi di progesterone da parte del corpo luteo,
-Progesterone,
-Estrogeni,
-CGP o HPL (chorionic growth hormone-prolactin o human placental lactogen), che favorisce la secrezione lattea e
stimola lo sviluppo dell’apparato scheletrico,
-Relaxina, che in prossimità del parto determina un rilassamento della sinfisi pubica e dei legamenti del bacino, in
modo tale da rendere più agevole il transito del feto lungo il canale genitale.
Ricapitolando….
….la placenta definitiva degli Equini è una placenta adeciduata,
vascolarizzata dall’allantoide, con microcotiledoni distribuiti su tutta la
superficie del corion (diffusa completa) e con un rapporto tra villi coriali
e mucosa uterina del tipo epitelio-coriale
….la placenta dei Suini è una placenta adeciduata, vascolarizzata
dall’allantoide, con villi distribuiti su tutta la superficie del corion ad
eccezione delle estremità (placenta diffusa incompleta). Il rapporto tra
villi coriali e mucosa uterina è di tipo epitelio- coriale
…. la placenta dei Ruminanti è adeciduata,
dall’allantoide, cotiledonare e sinepitelio-coriale
vascolarizzata
…. la placenta definitiva dei Carnivori è allantoidea, zonata, deciduata e
endotelio-coriale,mentre la placenta vitellina è funzionante solo per
breve tempo
…. la placenta dei Primati è anallantoidea, discoidale nell’uomo
(bidiscoidale nelle scimmie), deciduata e emo-coriale
Malformazioni congenite
Sono denominate malformazioni congenite tutte le anomalie morfologiche e funzionali presenti
alla nascita e derivanti da errori insorti durante lo sviluppo.
Le malformazioni congenite sono indotte da fattori teratogeni.
Tali fattori possono essere distinti in fattori endogeni, correlati ad alterazioni geniche o
cromosomiche, e fattori esogeni determinati dalle condizioni in cui viene a trovarsi l’embrione nel
corso dello sviluppo.
I fattori endogeni vanno distinti in cromosomici e genici.
Le alterazioni cromosomiche comprendono la delezione, la duplicazione, la traslocazione e
l’inversione di parti di cromosomi, ed inoltre la perdita (monosomia) o l’aggiunta (trisomia) di
cromosomi rispetto al corredo tipico della specie.
Le alterazioni geniche sono legate a cambiamenti della successione nucleotidica e spesso sono
mutazioni puntiformi di singoli geni.
I fattori esogeni possono essere classificati come naturali, o più precisamente casuali, quando
dipendono dall’ambiente o dalle condizioni di vita della madre, terapeutici, se sono correlati
all’introduzione di farmaci utilizzati a scopo terapeutico, e sperimentali quando sono impiegati per
lo studio dell’azione teratogena.
L’azione di un fattore esogeno viene definita diretta se agisce direttamente sul prodotto del
concepimento (ad esempio inoculazione di una sostanza nell’embrione o intervento chirurgico su
di un suo organo), o indiretta, quando agisce sulla madre o sulle strutture deputate agli scambi
materno-fetali
Le differenze tra specie, nella funzione di trasporto placentare, sono molto importanti e vanno tenute in
forte considerazione soprattutto quando dati ottenuti su animali da laboratorio vengono estrapolati in
riferimento all’uomo o ad animali domestici.
Ad esempio il Talidomide che non è teratogeno nei Roditori, è teratogeno a dosi elevate nel coniglio e
nella maggior parte dei Mammiferi, ed è altamente teratogeno anche a piccole dosi nei Primati. Al
contrario l’Acido Acetilsalicilico che è un potente teratogeno nei Roditori, non è teratogeno nei Primati.
L’effetto teratogeno può dipendere da vari fattori quali:
• lo stadio del processo riproduttivo e la sensibilità dell’embrione;
• la sensibilità della madre;
• le caratteristiche del fattore esogeno.
Lo stadio del processo riproduttivo è un
elemento fondamentale nella determinazione
delle malformazioni.
Si definisce periodo critico per una struttura
il momento in cui essa è in via di formazione.
Agenti che alterano la blastula, la gastrula, o
interferiscono con l’apposizione alla mucosa
uterina, provocano in genere aborto precoce.
Dallo stadio di neurula è più probabile che uno
o pochi apparati siano interessati all’azione di
un agente teratogeno, anche se spesso la
lesione indotta in un sistema può provocare
malformazioni secondarie in altre strutture.
Infine alcuni agenti teratogeni possono
interessare lo sviluppo del sistema nervoso
centrale di molte specie anche nei primi stadi
postnatali.
E’ da rilevare, però, che esiste una differente
sensibilità non solo tra le varie specie animali,
ma anche tra le razze e tra i singoli individui
nell’ambito della medesima specie e razza.
La sensibilità della madre.
La non idonea alimentazione materna, specialmente
se carente di alcune vitamine, aminoacidi essenziali e
sali minerali, nonchè l’anomalo equilibrio ormonale
materno possono essere causa di malformazioni.
Anche gli alcaloidi contenuti in alcuni vegetali
consumati dagli erbivori possono essere teratogeni ed
ancora, piante altamente tossiche per la madre
risultano anche embriotossiche.
Altro fattore importante è l’età della madre: è stato
dimostrato che sono più facilmente riscontrabili
malformazioni nei soggetti nati da madri molto giovani
o relativamente vecchie.
Le caratteristiche del fattore esogeno.
L’attività teratogena può essere legata a vari fattori tra cui
l’intensità e la durata dell’azione del fattore teratogeno sul
prodotto del concepimento.
Nel caso si tratti di un farmaco, al fine dell’azione teratogena,
hanno influenza rilevante: la dose, la modalità, la durata ed il
periodo di sviluppo durante il quale avviene la
somministrazione del farmaco. Inoltre l’azione teratogena
potrebbe non essere riferita direttamente alla molecola
introdotta in quanto il farmaco può essere trasformato
(metabolizzato) ed il metabolita potrebbe essere il vero
responsabile dell’effetto teratogeno.
Infine in alcuni casi l’azione teratogena può dipendere dal
potenziamento dell’effetto di due farmaci, di per sé inattivi,
che possono determinare malformazioni, se vengono usati
contemporaneamente.