sistema + ambiente = universo
La Bioenergetica è lo studio quantitativo delle
trasformazioni energetiche associate alle reazioni chimiche.
Queste trasformazioni obbediscono alle leggi della
Termodinamica.
Leggi della Termodinamica
1a Legge (principio di conservazione dell’energia)
L’energia totale dell’universo è costante
2a Legge
Un processo può avvenire spontaneamente solo
se l’entropia dell’universo aumenta
Ad una certa T, lo stato energetico di un sistema chimico
può essere descritto in termini di:
H = entalpia (contenuto termico)
S = entropia (grado di disordine)
G = energia libera di Gibbs
(energia utile a compiere un lavoro)
Le variazioni di G, H, S sono tra loro correlate
dall’equazione ΔG = ΔH - TΔS
ΔG<0
Né ΔH né Δs, ma solo ΔG può permettere di predire
la spontaneità di una reazione.
ΔG è espresso in kcal/mole
o kJoule/mole (1 kcal=4.2 kJ)
ΔG: standard chimico
pH=0
ΔG’: standard biochimico pH=7
Il ΔG’ dipende solo dallo
stato iniziale e da quello finale
e non dalla via seguita dalla
trasformazione.
ΔG’ può predire la direzione
di una reazione,
ma non la velocità che dipende
dall’enzima che catalizza
quella reazione.
ΔG’0 = variazione di energia libera standard
- è correlata alla k’eq
- è una costante della reazione
- considera [S]=[P]=1 M
Nella realtà le concentrazioni di S e di P sono ben lontane
dall’essere 1M, quindi se consideriamo le reali concentrazioni:
ΔG’ = variazione di energia libera reale
- è una variabile
- dipende da ΔG’0 e dalle reali [S] e [P]
ΔG’0 = variazione di energia libera standard
- è correlata alla k’eq
- è una costante della reazione
- considera [S]=[P]=1 M
ΔG’ = variazione di energia libera reale
- è una variabile
- dipende da ΔG’0 e dalle reali [S] e [P]
Una reazione termodinamicamente non favorevole può essere
trascinata da una reazione favorevole ad essa accoppiata
Le variazioni di energia libera
sono additive.
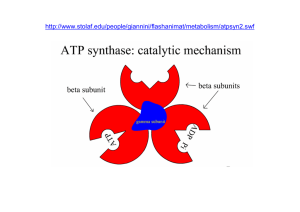
L’ATP è il principale trasportatore di energia ed è il punto di
unione tra i processi catabolici e quelli che richiedono energia.
La contrazione muscolare rappresenta uno dei pochi casi in cui
l’idrolisi dell’ATP è di per sé la fonte della energia chimica.
In tutti gli altri casi l’ATP interagisce covalentemente
con un substrato attraverso il trasferimento di
un gruppo (Pi, PPi, AMP).
Basi strutturali dell’elevato ΔG’0 dell’ATP
-
Repulsione elettrostatica
-
Stabilizzazione per risonanza
-
“
“
ionizzazione
-
“
“
idratazione
La forte tensione dovuta alla repulsione elettrostatica
diminuisce quando l’ATP è idrolizzato
I prodotti dell’idrolisi hanno una maggiore stabilità
rispetto alla molecola intatta dell’ATP.
SCISSIONE ORTOFOSFORICA
Benché i ΔG’0 siano fortemente negativi, l’ATP è
stabile e idrolizzabile solo dallo specifico enzima.
Composti fosforilati quali PEP,1,3-BPG, CP hanno un ΔG’0
di idrolisi superiore a quello dell’ATP.
È questa posizione intermedia che consente all’ATP di funzionare
in modo efficace come trasportatore di gruppi fosforici.
L’ATP e l’ADP sono sempre presenti come complessi con
il Mg2+, che maschera in parte le loro cariche negative.
Il ΔG’ dell’idrolisi dell’ATP dipende da:
[reali reagenti], pH, [Mg2+]
ΔG’0 = - 7.3 kcal/mole
ΔG’ = - 12 kcal/mole
L’ATP viene continuamente consumato e rigenerato:
il turnover è molto rapido, una molecola viene
consumata circa 1 minuto dopo la sua formazione.
Nel rene e nel cervello più dei 2/3 dell’ATP prodotto
vengono utilizzati per pompare Na+ e K+ attraverso
la membrana plasmatica (pompa Na+K+ ATPasi).
Anche i tioesteri possiedono un’elevata energia
libera di idrolisi negativa
ΔG’0 = - 7.5 kcal/mole