CAPITOLO 1
Principi di endocrinologia
SEU
Sistema endocrino .......................................................................................................... 1
Classificazione degli ormoni .......................................................................................... 2
Ormoni peptidici .......................................................................................................... 4
Ormoni steroidei ......................................................................................................... 5
Ormoni derivati dagli aminoacidi ................................................................................ 6
Recettori ormonali ........................................................................................................... 7
Recettori di membrana ............................................................................................... 8
Recettori nucleari ..................................................................................................... 13
Effetti fisiologici degli ormoni......................................................................................... 14
Sistemi di retroregolazione o di regolazione ormonale a feedback .............................. 14
Ritmi endocrini .............................................................................................................. 16
Ormoni e sistema immune ........................................................................................... 17
Malattie endocrine .......................................................................................................... 18
Prevalenza .................................................................................................................... 18
Classificazione .............................................................................................................. 18
Endocrinopatie da insufficienza ormonale ........................................................................... 18
Endocrinopatie da eccesso ormonale ...................................................................... 19
Diagnosi ........................................................................................................................ 20
Anamnesi ed esame obiettivo ................................................................................... 20
Indagini di laboratorio ................................................................................................ 20
Indagini per immagini ................................................................................................ 22
Terapia .......................................................................................................................... 22
Terapia ormonale in malattie non endocrine e nello sport ......................................... 23
Eritropoietina: impiego clinico e nel doping....................................................................... 23
ROMA
Sistema endocrino
Il sistema endocrino è un sistema di comunicazione tra cellule deputato al controllo della crescita, della riproduzione e delle funzioni essenziali per la sopravvivenza. Il
termine endocrino, dal greco ενδο (interno) e κρινω (secerno), indica la secrezione
di molecole biologicamente attive, i cosiddetti ormoni, all’interno del corpo; il termine
esocrino indica, invece, il processo di secrezione di specifiche sostanze all’esterno
oppure all’interno di un lume attraverso appositi dotti escretori. Le cellule secernenti
ormoni sono localizzate in organi specifici, le ghiandole endocrine, oppure sono disperse nell’ambito di tessuti non endocrini, costituendo il sistema endocrino diffuso.
Il termine ormone deriva dal greco ορµαω (mettere in moto, eccitare) e indica una
sostanza che è prodotta da una cellula endocrina, viene rilasciata nel circolo sanguigno
ed evoca risposte funzionali in cellule distanti dalla sua sede di produzione. Pertanto
2
Endocrinologia
gli ormoni sono considerati molecole informazionali o messaggeri chimici in quanto
responsabili della comunicazione intercellulare. Si sottolinea che gli effetti biologici variano in base al tipo di ormone. Per l’espletamento dell’azione ormonale è necessario
che l’ormone, una volta sintetizzato e secreto, venga trasportato nel circolo sanguigno
fino a raggiungere i tessuti bersaglio, dove sono presenti i recettori ormonali, strutture
specializzate che riconoscono specificamente l’ormone e ne traducono il messaggio
nella cellula bersaglio. Nella definizione di comunicazione endocrina è implicito il
concetto di trasporto dell’ormone dalla sua sede di produzione all’organo bersaglio. La
comunicazione affidata agli ormoni avviene soprattutto attraverso il circolo ematico, ma
anche attraverso altre modalità: alcune molecole agiscono sulle cellule circostanti alla
cellula di origine (comunicazione paracrina), mentre altre interagiscono con la stessa
cellula che li ha prodotti (comunicazione autocrina). Esistono infine sostanze sintetizzate da neuroni che sono responsabili della comunicazione neurocrina, una forma
specializzata di comunicazione intercellulare che si distingue in sinaptica, se la molecola viene secreta attraverso una struttura sinaptica, e non sinaptica o neurosecretoria,
se viene rilasciata nel circolo sanguigno o nello spazio extracellulare, Fig. 1.1.
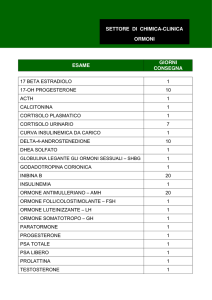
Nella Tab. 1.1 sono schematicamente riportati i principali ormoni attualmente noti.
SEU
Classificazione degli ormoni
In base alla struttura chimica gli ormoni sono distinti in tre classi principali: peptidi, steroidi e derivati dagli aminoacidi, Tab 1.2. La struttura molecolare degli ormoni
ne determina le caratteristiche funzionali. Gli ormoni peptidici sono idrosolubili e
pertanto circolano liberi nel plasma ed esplicano i loro effetti dopo il legame con recettori localizzati sulla membrana delle cellule bersaglio, senza penetrare all’interno
ROMA
Figura 1.1 – Principali meccanismi di secrezione.
Principi di endocrinologia
3
Tabella 1.1 – Ormoni principali.
Sede di produzione
Ipotalamo
Adenoipofisi
Neuroipofisi
Tiroide
Paratiroidi
Cute, intestino
Nome
Somatostatina
Ormone stimolante il rilascio di GH
Dopamina
Ormone stimolante il rilascio di ACTH
Ormone stimolante il rilascio di TSH
Ormone stimolante il rilascio di gonadotropine
SEU
Sigla
SRIH
GHRH
DA
CRH
TRH
GnRH
Ormone della crescita o somatropina
Prolattina
Adrenocorticotropina o corticotropina
Ormone stimolante la tiroide o tireotropina
Ormone luteinizzante
Ormone follicolostimolante
GH
PRL
ACTH
TSH
LH
FSH
Ormone antidiuretico o vasopressina
Ossitocina
ADH
OT
Tiroxina o tetraiodiotironina
Triiodotironina o triiodiotironina
Calcitonina
Ormone paratiroideo
T4
T3
CT
PTH
Vitamina D3 o colecalciferolo
–
Corticale surrenale
Cortisolo
Aldosterone
Deidroepiandrosterone
Androstenedione
17-OH-progesterone
–
–
–
–
–
Midollare surrenale
Adrenalina
Noradrenalina
Sangue
Angiotensina II
Ovaio
Testicolo
Rene
Fegato
17 β−estradiolo
Progesterone
Testosterone
Inibina
Eritropoietina
Diidrossivitamina D o calcitriolo
ROMA
Fattore di crescita insulino-simile 1
Idrossivitamina D o calcidiolo
A
NA
E2
Pg
T
–
EPO
1,25(OH)2D
IGF-1
25(OH)D
Sistema gastroenterico
Secretina
Gastrina
Colecistochinina
Peptide inibente lo stomaco
Peptide glucagone-simile 1
Motilina
Peptide intestinale vasoattivo
Somatostatina
Bombesina
Glucagone
Ghrelina
Polipeptide pancreatico
–
–
CCK
GIP
GLP-1
–
VIP
SRIH
–
–
–
PP
Pancreas
Insulina
Glucagone
Somatostatina
Sostanza P
Ins
Glu
SRIH
Tessuto adiposo
Leptina
Epifisi
Melatonina
Tessuto trofoblastico
Gonadotropina corionica
Lep
–
hCG
4
Endocrinologia
Tabella 1.2 – Classificazione degli ormoni in base alla struttura chimica.
Peptidici
Peptidi e polipeptidi
CRH, GnRH, GHRH, TRH, PRL, ACTH, GH, IGF-1, ADH, ossitocina, angiotensina II,
renina, ormone natriuretico atriale, calcitonina, PTH, colecistochinina, gastrina, ghrelina,
glucagone, GIP, GLP-1, insulina, VIP, polipeptide pancreatico, somatostatina, leptina
Glicoproteine
FSH, LH, TSH, hCG
Peptidi
Steroidi
Tiroidei
pro-ormone
enzimatica
enzimatica
enzimatica
deposito in granuli
passaggio diretto deposito nella
nel plasma
colloide
idrosolubili
liposolubili
Recettore
SEU
Catecolamine
Meccanismo
d’azione
Steroidei
Cortisolo, aldosterone, androstenedione, deidroepiandrosterone, estradiolo, estriolo,
estrone, progesterone, testosterone, diidrotestosterone, calcidiolo, calcitriolo
Derivati da
aminoacidi
Derivati dal triptofano
Melatonina, serotonina
Derivati della tirosina
Adrenalina, noradrenalina, dopamina, tiroxina, triiodiotironina
di queste. Al contrario gli ormoni steroidei sono liposolubili e quindi diffondono liberamente all’interno della cellula esercitando la loro azione dopo legame con recettori
localizzati nel nucleo, Tab. 1.3.
Tabella 1.3 – Caratteristiche generali degli ormoni.
Sintesi
Secrezione
Solubilità
Trasporto
Variazione dei
livelli plasmatici
Emivita
Metabolismo
Effetto biologico
deposito nei granuli
cromaffini
liposolubili
idrosolubili
non legati a proteine legati a proteine
plasmatiche
plasmatiche
legati a proteine
plasmatiche
non legati a proteine plasmatiche
rapida
lenta
lenta
rapida
minuti (peptidi),
ore (glicoproteine)
ore
giorni
secondi/minuti
ROMA
organo bersaglio,
plasma
fegato
fegato
organo bersaglio,
plasma
membrana
nucleare
nucleare
membrana
attivazione di enzimi, sintesi proteica
trasporto ionico
sintesi proteica
attivazione di enzimi, trasporto ionico
rapido
lento
rapido
lento
Ormoni peptidici
Comprendono peptidi, polipeptidi e glicoproteine.
Sintesi. È simile a quella di tutte le altre proteine e si realizza a partire da geni
specifici che vengono trascritti con formazione di RNA, rimodellato nel nucleo attraverso il processo di splicing con produzione di RNA messaggero maturo. Il fenomeno dello splicing varia nei diversi tessuti e determina la formazione di ormoni
differenti a partire dalla trascrizione di un gene comune. Una volta trasferito dal
nucleo al citoplasma l’RNA messaggero viene tradotto in una catena peptidica in
corrispondenza del reticolo endoplasmatico rugoso. La sintesi dell’ormone peptidico
Principi di endocrinologia
5
si completa attraverso modificazioni co-traduzionali e post-traduzionali (rimozione
di alcune sequenze, formazione di ponti disolfuro, iodazione, fosforilazione, solfatazione, acetilazione glicosilazione, assemblaggio di più subunità proteiche, ecc.). Le
modificazioni post-traduzionali si realizzano nelle cisterne dell’apparato di Golgi, nel
quale gli ormoni proteici neosintetizzati vengono trasferiti mediante apposite vescicole di trasporto. L’ormone peptidico finale viene immagazzinato in appositi granuli
di secrezione, che si accumulano all’interno della cellula.
Secrezione. Si realizza attraverso due modalità: la secrezione costitutiva, che
consiste nella rapida esocitosi di ormone neosintetizzato a livello della membrana
apicale della cellula, e la secrezione regolata, che prevede invece la formazione di
scorte di ormone, immagazzinato e concentrato nei granuli, e rilasciato in risposta
ad uno stimolo in corrispondenza della superficie laterobasale. La presenza di depositi intracellulari di ormone consente alla cellula di rispondere agli stimoli secretori
in un tempo più breve rispetto a quello necessario per la sintesi “de novo” delle
molecole ormonali.
Trasporto e metabolismo. Gli ormoni peptidici circolano in forma libera nel plasma, con poche eccezioni. L’emivita dei peptidi è di pochi minuti, mentre gli ormoni
glicoproteici hanno un’emivita più lunga, compresa tra i 50 minuti e le 4 ore e proporzionale ai residui di acido sialico delle catene oligosaccaridiche. Infatti gli ormoni
glicoproteici sono metabolizzati a livello epatico e la presenza di acido sialico ne
previene la degradazione. Gli ormoni peptidici non glicoproteici sono invece degradati dalle cellule bersaglio e solo in minima parte vengono distrutti dalle proteasi
plasmatiche; una piccola quota è escreta direttamente con le urine.
SEU
Ormoni steroidei
Gli ormoni steroidei sono accomunati dalla presenza del nucleo steroideo del
ciclopentanoperidrofenantrene derivato dal colesterolo. Si distinguono ormoni con
nucleo steroideo integro, che comprendono gli steroidi gonadici e surrenali, e ormoni con nucleo steroideo aperto, rappresentati dalla Vitamina D e dai suoi metaboliti.
Steroidi gonadici e surrenali. Si dividono in sottogruppi in base al numero di
atomi di carbonio del nucleo steroideo: il progesterone, i glucocorticoidi e i mineralocorticoidi derivano dal nucleo del pregnano che contiene 21 atomi di carbonio
mentre gli estrogeni derivano dal nucleo dell’estrano con 18 atomi di carbonio e gli
androgeni dal nucleo dell’androstano con 19 atomi di carbonio, Fig. 1.2. La sintesi
degli ormoni steroidei inizia da un precursore comune, il colesterolo, e procede
attraverso tappe biosintetiche che sono identiche nel surrene, nell’ovaio e nel testicolo. La distribuzione tessuto-specifica degli enzimi coinvolti nella steroidogenesi
consente la produzione differenziale degli steroidi nelle tre ghiandole: i glucocorticoidi e i mineralcorticoidi sono sintetizzati dal surrene, gli androgeni sono prodotti
dalle cellule di Leydig del testicolo, dal surrene e dalle cellule della teca dell’ovaio,
ed infine gli estrogeni e il progesterone sono i principali ormoni steroidei sintetizzati dall’ovaio; gli estrogeni sono prodotti anche per conversione periferica dagli
androgeni, che avviene soprattutto a livello del tessuto adiposo. Contrariamente
agli ormoni peptidici, gli steroidi non sono immagazzinati all’interno delle cellule,
ma vengono secreti subito dopo essere stati sintetizzati. Inoltre gli ormoni steroidei
circolano nel plasma legati a specifiche proteine di trasporto; solo l’ormone libero è
in grado di interagire con i recettori e quindi di esprimere le sue azioni biologiche.
ROMA
6
Endocrinologia
SEU
Figura 1.2 – Nucleo dei principali ormoni steroidei.
Il legame con le proteine di trasporto controlla la clearance dell’ormone proteggendolo dal catabolismo, garantisce una riserva ormonale circolante e controlla la
disponibilità dell’ormone per le cellule bersaglio, senza influenzarne l’attività biologica. Il metabolismo degli steroidi consiste in modificazioni dei gruppi funzionali
con formazione di composti idrosolubili e quindi eliminabili attraverso il rene. Per
maggiori informazioni si rimanda ai capitoli Corticosurrene, Ipertensioni endocrine,
Sistema riproduttivo femminile e maschile.
Vitamina D. È uno steroide di cui esistono due forme naturali, l’ergocalciferolo
(vitamina D2) di derivazione vegetale e il colecalciferolo (vitamina D3) di derivazione animale, prodotto per irradiazione ultravioletta del 7-deidrocolesterolo contenuto
nella cute. Sia la vitamina D2 che la vitamina D3, per poter diventare biologicamente
attive, necessitano di modificazioni metaboliche: l’idrossilazione epatica determina
la formazione di calcidiolo o 25(OH)D, che è poi trasformato a livello renale in calcitriolo o 1,25(OH)2D, dotato della massima attività biologica. Tutte le forme di vitamina
D circolano nel sangue legate a specifiche proteine di trasporto e la principale via di
eliminazione è il rene. Per maggiori informazioni si rimanda al capitolo Paratiroidi.
ROMA
Ormoni derivati dagli aminoacidi
Comprendono i derivati del triptofano (serotonina e melatonina) e i derivati della
tirosina (ormoni tiroidei e catecolamine). Gli ormoni tiroidei, prodotti dalle cellule
follicolari della tiroide, sono delle iodotironine (T3 e T4). Per maggiori informazioni si
rimanda al capitolo Tiroide. Le catecolamine dotate di effetti ormonali sono l’adrenalina e la noradrenalina sintetizzate, oltre che dai neuroni del sistema nervoso
simpatico, anche a livello della midollare del surrene. Per maggiori informazioni si
rimanda al capitolo Ipertensioni endocrine.
Principi di endocrinologia
7
Recettori ormonali
Gli ormoni circolano nel plasma a concentrazioni bassissime (10-15-10-9 M) e pertanto le cellule bersaglio devono riconoscerli selettivamente in presenza di un eccesso di molecole con struttura chimica simile. La capacità di riconoscimento selettivo è
garantita dalla presenza di strutture di ricezione specializzate, i recettori, che legano
gli ormoni e ne mediano le azioni. I recettori sono localizzati sulla superficie plasmatica o all’interno della cellula bersaglio in base alla struttura chimica dell’ormone: gli
ormoni che non possono attraversare la membrana (peptidi, catecolamine) si legano a recettori localizzati sulla membrana plasmatica, mentre quelli che diffondono
attraverso la membrana plasmatica all’interno della cellula (steroidi, iodotironine)
riconoscono recettori intracellulari, Fig. 1.3.
I recettori hanno caratteristiche strutturali comuni: tutti presentano una regione
o “dominio” che riconosce e lega l’ormone e una regione deputata alla generazione
di un segnale intracellulare che traduce il messaggio ormonale in risposte funzionali
della cellula bersaglio. Anche le proprietà che regolano il legame ormone-recettore
sono le stesse in quanto tutti i recettori sono caratterizzati da specificità, elevata affinità, saturabilità e proprietà di trasduzione del segnale. La specificità è la capacità di
riconoscere un solo ormone o più ormoni con funzioni analoghe. L’affinità definisce
il legame preferenziale con uno specifico ormone in maniera stabile e reversibile,
rappresenta cioè il grado di complementarietà strutturale che determina la forza
dell’associazione. La saturabilità indica la capacità di legare l’ormone fino ad un
massimo che corrisponde all’occupazione di tutti i recettori. La capacità di trasduzione rappresenta la proprietà di evocare risposte specifiche a livello delle cellule
bersaglio. Queste caratteristiche distinguono i recettori da altre strutture in grado
SEU
ROMA
Figura 1.3 – Meccanismo d’azione degli ormoni.
8
Endocrinologia
di legare l’ormone quali, ad esempio, le proteine di trasporto presenti nel plasma,
che riconoscono e legano l’ormone ma che hanno una bassa affinità, una bassa
specificità di legame, un’elevata capacità e non sono in grado di trasdurre il segnale
veicolato dall’ormone.
Agonisti, superagonisti, antagonisti. Sono molecole che presentano caratteristiche strutturali simili ad un determinato ormone e che pertanto sono in grado di
interagire con il suo recettore evocando o meno una risposta ormonale. Gli agonisti
si legano al recettore con la stessa affinità dell’ormone naturale, attivando il meccanismo di trasduzione del segnale ormonale e provocando la medesima risposta cellulare. I superagonisti interagiscono con il recettore con un’affinità maggiore rispetto
all’ormone naturale evocando una risposta più intensa e duratura, come accade per
esempio nel caso del desametasone, un glucocorticoide di sintesi utilizzato a fini
terapeutici per le sue proprietà antinfiammatorie. Invece, gli antagonisti non evocano
risposte biologiche perché incapaci di attivare il meccanismo di trasduzione; pertanto competono con l’ormone naturale per il legame al recettore inibendo la risposta
delle cellule bersaglio. Tra gli antagonisti di sintesi si ricorda il tamoxifene, un antagonista degli estrogeni impiegato nel trattamento del carcinoma della mammella.
Alcuni composti si comportano come agonisti-antagonisti parziali in quanto, dopo il
legame al recettore, evocano risposte funzionali inferiori rispetto a quelle prodotte
dall’ormone naturale e quindi esercitano anche una parziale inibizione.
Modificazioni quantitative dei recettori. L’espressione di specifici recettori è
determinata geneticamente nei diversi tessuti. Tuttavia, in condizioni fisiologiche,
sia il numero di recettori che la loro affinità sono regolati da stimoli ormonali. I meccanismi molecolari che intervengono in questi fenomeni sono molteplici e possono
coinvolgere la modulazione dell’intensità di sintesi e di degradazione dei recettori,
il loro sequestro in comparti subcellulari inaccessibili all’ormone e anche il cambiamento delle loro proprietà funzionali. Il numero di recettori nelle cellule bersaglio è
regolato dagli stessi ormoni attraverso tre modalità: la down-regulation consiste nella riduzione del numero dei recettori a causa di un’eccessiva stimolazione ormonale,
con desensibilizzazione delle cellule bersaglio che mostrano una ridotta capacità
di rispondere all’ormone; la up-regulation rappresenta il meccanismo mediante il
quale alcuni ormoni (angiotensina II, prolattina) inducono l’espressione dei propri
recettori nelle cellule bersaglio, un fenomeno dovuto alla sintesi di nuove molecole di
recettore oppure alla “ricomparsa”, a livello della membrana plasmatica, di recettori
precedentemente internalizzati; il priming indica il fenomeno per cui l’espressione
del recettore di un ormone è controllata da un diverso ormone (ad es. l’FSH induce
la sintesi di recettori per l’LH nelle cellule di Leydig).
SEU
ROMA
Recettori di membrana
Sono strutture localizzate sulla membrana cellulare deputate a trasmettere il
messaggio di ormoni che non sono in grado di diffondere all’interno della cellula perché non liposolubili. Sono distinti, in base al meccanismo d’azione, in 4 sottoclassi:
1) recettori tirosino-chinasici; 2) recettori guanilato-ciclasici; 3) recettori accoppiati
alle proteine G; 4) recettori citochinici. I recettori appartenenti alle prime 2 sottoclassi
sono dotati di funzione effettrice in quanto svolgono attività chinasica o consentono
l’afflusso di ioni, mentre quelli delle sottoclassi 3 e 4 necessitano dell’intervento di un
secondo messaggero (proteina G o tirosino-chinasi intracitoplasmatiche) per adempiere alla propria funzione.
Principi di endocrinologia
9
Recettori tirosino-chinasici. I recettori ad attività tirosino-chinasica, il cui prototipo è rappresentato dal recettore per l’insulina, sono in grado di attivare direttamente le risposte cellulari senza la mediazione di sistemi di trasduzione del segnale
e di messaggeri intracellulari. Sono circa 100 i recettori tirosino-chinasici noti, che si
caratterizzano per un’elevata omologia di sequenza nella regione chinasica, mentre
differiscono sostanzialmente nella porzione extracellulare così da poter riconoscere
ligandi diversi. Strutturalmente sono costituiti da tre regioni, una extracellulare, una
intracellulare e una trasmembrana che unisce le due porzioni intra- ed extracitoplasmatica. Il requisito iniziale per la traduzione del segnale è la dimerizzazione del
recettore indotta dal legame con l’ormone. Dopo la dimerizzazione del recettore i
residui tirosinici presenti nella porzione intracitoplasmatica vengono fosforilati. Questo processo comporta cambiamenti conformazionali del recettore che acquisisce
la capacità di legare sia l’ATP che altre proteine definite “accessorie”, che vengono
fosforilate con conseguente attivazione di altre vie di trasduzione del segnale (ad es.
la via della fosfoinositolo 3 chinasi).
Recettori guanilato-ciclasici. La guanilato-ciclasi regola la via del GMP-ciclico,
un nucleotide ciclico che media gli effetti di numerosi peptidi e, tra gli ormoni, dell’ormone natriuretico atriale. Il recettore di questo ormone è costituito da una singola regione transmembrana con una porzione extracitoplasmatica che riconosce l’ormone
e da un’ampia regione intracellulare che presenta un sito ad attività simil-chinasica.
Si ritiene che l’interazione ormone-recettore determini dei mutamenti conformazionali che interrompono l’azione inibitoria sul sito simil-chinasico consentendo l’attivazione della guanilato-ciclasi.
Recettori accoppiati alle proteine G. La maggior parte dei recettori di membrana è accoppiata a sistemi di trasduzione del segnale rappresentati dalle proteine G.
Il meccanismo d’azione consiste nel legame dell’ormone al recettore con successivo
intervento di un sistema di trasduzione del messaggio che, agendo su un sistema
effettore, porta alla formazione di un secondo messaggero; quest’ultimo regola la
risposta funzionale delle cellule bersaglio allo stimolo ormonale, Fig. 1.4.
SEU
ROMA
Figura 1.4 – Rappresentazione schematica di un recettore di membrana accoppiato alle proteine G.
10
Endocrinologia
I recettori accoppiati alle proteine G, pur essendo eterogenei dal punto di vista
strutturale, hanno caratteristiche comuni. Sono proteine integrali di membrana costituite nella maggior parte dei casi da sette segmenti peptidici trasmembrana e dotate
di un dominio extracellulare deputato al legame con l’ormone e di una porzione intracellulare che interagisce con le proteine G. Le differenze nella sequenza aminoacidica che caratterizza i molteplici recettori accoppiati alle proteine G, permettono di
individuare tre famiglie recettoriali, a loro volta suddivise in base alla lunghezza della
porzione aminoterminale.
Le proteine G sono membri di una superfamiglia di proteine leganti il guanosin-trifosfato (GTP), che comprende proteine del citoscheletro come la tubulina, proteine
che regolano la sintesi proteica e i prodotti del protoncogene ras. Sono associate al
versante citoplasmatico della membrana cellulare e presentano una struttura eterotrimerica essendo costituite da tre subunità, α, β e γ, ciascuna codificata da un gene
diverso. Quando la proteina è in condizioni di riposo le subunità sono associate
mentre durante l’attivazione che segue il legame ormone-recettore si dissociano
in α e β-γ. In assenza di stimolazione ormonale la proteina G in forma inattiva (αβ-γ) è legata al GDP, Fig. 1.5. L’attivazione del recettore, determinata dal legame
dell’ormone, catalizza lo scambio GDP-GTP attraverso un cambiamento conformazionale della subunità α che ne induce il legame al GTP. Il legame del GTP causa la
dissociazione di α dal recettore e dal complesso β-γ e il complesso subunità α-GTP
interagisce con l’effettore. La subunità α è dotata di attività GTP-asica che idrolizza
il GTP e riporta il meccanismo allo stato di riposo.
I sistemi effettori regolati dalle proteine G sono numerosi; quelli di maggiore interesse endocrinologico sono l’adenilato ciclasi, attivata dalla proteina Gs, e la fosfolipasi C, attivata dalla proteina Gq.
Adenilato-ciclasi. È una glicoproteina localizzata sul versante citoplasmatico della membrana cellulare, che converte l’ATP in AMP-ciclico (cAMP). Il cAMP, che funge da messaggero intracellulare, si lega alla proteino-chinasi A, proteina tetramerica
costituita da due subunità regolatorie e due subunità catalitiche. Il cAMP interagisce
SEU
ROMA
Figura 1.5 – Meccanismo d’azione delle proteine G.
Principi di endocrinologia
11
con le subunità regolatrici permettendo il distacco delle subunità catalitiche e trasformando quindi la chinasi nella sua forma attiva, Fig. 1.6. La proteino-chinasi attivata
catalizza reazioni di fosforilazione trasferendo il fosfato dall’ATP ai residui di serina e
treonina delle proteine, la cui funzione viene così attivata o inibita. Nel caso in cui le
proteine fosforilate siano enzimi, la loro attivazione si traduce in risposte funzionali
metaboliche che regolano le attività controllate dall’ormone. Se invece si tratta di
fattori di trascrizione, questi una volta attivati agiscono a livello nucleare modulando
la trascrizione genica: è stato infatti identificato un fattore di trascrizione (c-AMP
response element-binding protein, CREB) in grado di legarsi a sequenze geniche
responsive all’AMP-ciclico (cAMP response element, CRE).
Il meccanismo mediato dall’AMP-ciclico permette un’amplificazione a cascata del
segnale. Infatti ciascun complesso ormone-recettore può attivare circa 10 molecole
di proteina Gs, ognuna delle quali attiva una molecola di adenilato ciclasi producendo 100-150 molecole di AMP-ciclico. Il segnale ormonale viene quindi amplificato di
circa 1.000 volte. Le catecolamine, il glucagone, gli ormoni glicoproteici ipofisari, il
PTH agiscono aumentando le concentrazioni di AMP-ciclico. Per altri ormoni, come
la somatostatina, il messaggio ormonale non è mediato dall’aumento ma dalla riduzione dell’AMP-ciclico, dovuta ad una diminuzione dell’attività dell’adenilato ciclasi
di membrana mediata da proteine Gi.
Fosfolipasi C. Alcuni ormoni, quali TRH, GnRH, catecolamine e angiotensina II,
attivano una classe di secondi messaggeri rappresentata dai componenti della via
del fosfatidilinositolo 4,5-difosfato. L’interazione ormone-recettore determina l’attivazione del sistema di trasduzione mediato da una classe di proteine G dette Gq.
L’attivazione del trasduttore stimola la fosfolipasi C che scinde il fosfatidilinositolo
SEU
ROMA
Figura 1.6 – Rappresentazione schematica dell’attivazione dell’adenilato-ciclasi ed eventi intracellulari regolati dall’AMP- ciclico.
12
Endocrinologia
4,5-difosfato (PIP2) con formazione di diacilglicerolo (DAG) e inositolo 1,4,5-trifosfato (IP3) che agiscono come secondi messaggeri, Fig. 1.7. L’IP3 induce il rilascio di
Ca++ dal reticolo endoplasmatico, mentre il DAG, in presenza di Ca++ e fosfatidilserina, attiva una specifica proteino-chinasi detta proteino-chinasi C o Ca++/fosfolipide
dipendente. Un ulteriore ruolo del Ca++ è rappresentato dal suo legame alla calmodulina, proteina in grado di regolare specifiche proteino-chinasi. La fosforilazione dei
residui di serina e di treonina delle proteine, catalizzata dalla proteino-chinasi C, si
traduce in modificazioni funzionali delle attività enzimatiche e dei processi metabolici
della cellula.
Recettori citochinici. Hanno un meccanismo d’azione simile a quello dei recettori tirosino-chinasici, con la differenza che non possiedono una capacità di fosforilazione intrinseca ma necessitano di una proteina associata. I ligandi per questi recettori comprendono ormoni come il GH, la prolattina, la leptina, l’eritropoietina e gran
parte delle citochine, responsabili della regolazione della risposta immune. Sono
costituiti da un’unica regione transmembrana e presentano un’omologia strutturale
sia nella regione citoplasmatica che in quella extracellulare. Generalmente ciascuna
molecola del ligando si lega a 2 subunità uguali (omodimero) che formano il recettore attivo. In alcuni casi tuttavia il legame può avvenire con subunità diverse tra di
loro (eterodimero); questo fenomeno consente ad uno stesso ligando, in genere una
citochina, di legarsi a differenti coppie di subunità così da ottenere una risposta cellulare più modulata. L’attività tirosino-chinasica è svolta da una famiglia di proteine,
note come JAK (Janus family of tyrosine kinases), le quali dopo la dimerizzazione
del recettore si attivano e fosforilano se stesse attivando a cascata altre proteine tra
le quali i fattori di trascrizione STAT e la via della MAP chinasi.
SEU
ROMA
Figura 1.7 – Rappresentazione schematica dell’attivazione della fosfolipasi C.
Principi di endocrinologia
13
Recettori nucleari
Gli ormoni liposolubili, come gli steroidi e gli ormoni tiroidei, attraversano la membrana cellulare esercitando i loro effetti dopo essersi legati a recettori intracellulari. Il
complesso ormone-recettore riconosce, all’interno del nucleo, specifiche sequenze
di DNA e regola la trascrizione di geni specifici e la loro traduzione in RNA messaggero e proteine. I ligandi che riconoscono i recettori nucleari sono accomunati dal
basso peso molecolare e dal fatto di non essere codificati direttamente dal genoma
in quanto derivano tutti da precursori assunti con la dieta e successivamente modificati nel nostro organismo. Inoltre la liposolubilità che caratterizza questi ormoni
ne garantisce un rapido assorbimento a livello intestinale, permettendo così il loro
impiego a fini terapeutici. Alcuni recettori nucleari sono localizzati nel citoplasma,
mentre altri si trovano solamente all’interno del nucleo. Dal punto di vista funzionale
i recettori nucleari possono essere suddivisi in 2 famiglie: la prima, che include i
recettori per glucocorticoidi, mineralocorticoidi, androgeni e progesterone è detta la
famiglia dei recettori per gli ormoni steroidei; la seconda, comprendente i recettori
per ormoni tiroidei, estrogeni e vitamina D, è indicata come famiglia dei recettori per
gli ormoni tiroidei.
Recettori degli ormoni steroidei. La loro sequenza aminoacidica è organizzata
in porzioni, dette dominii, dotate di proprietà funzionali differenti. Nella parte aminoterminale è presente il dominio di transattivazione che partecipa alla regolazione
genica, mentre nella porzione carbossiterminale è localizzato il dominio deputato
al legame con l’ormone. La parte centrale della sequenza aminoacidica contiene
invece il dominio che si lega al DNA, caratterizzato da una struttura digitiforme determinata da un legame tra un atomo di zinco e quattro residui di cisteina (zinc finger
region). Questa porzione è la più conservata tra i vari recettori della superfamiglia.
In assenza dell’ormone i recettori degli ormoni steroidei sono presenti nel citoplasma
in forma di complessi inattivi con proteine non recettoriali, definite proteine da shock
termico (heat shock proteins = HSP) perché inducibili dopo esposizione delle cellule ad elevate temperature. Il legame con l’ormone induce modificazioni conformazionali del recettore che permettono la dissociazione dalle HSP con formazione di
dimeri del complesso ormone-recettore ed esposizione del dominio che interagisce
con il DNA. Esistono specifiche sequenze geniche ormono-responsive (hormone
response elements = HRE), che sono denominate in base all’ormone a cui si riferiscono (glucocorticoidi, progesterone, androgeni) e che rappresentano gli elementi
di regolazione della trascrizione da parte degli ormoni steroidei, cioè permettono il
controllo dell’espressione del gene da parte dell’ormone specifico.
Recettori degli ormoni tiroidei. I recettori nucleari degli ormoni tiroidei, codificati dal protooncogene c-erbA, hanno una struttura simile a quella dei recettori per gli
steroidi; la regione che si lega al DNA è quella che presenta la maggiore omologia
tra i membri della superfamiglia c-erbA, mentre differiscono la regione C-terminale,
che lega l’ormone, e la regione N-terminale, più corta di quella dei recettori degli
steroidi. I recettori della famiglia degli ormoni tiroidei sono localizzati nel nucleo, legati alla cromatina e non sono associati alle HSP. Il legame dell’ormone al recettore
trasforma quest’ultimo in una forma funzionalmente attiva che interagisce con specifiche sequenze di DNA localizzate nella regione regolatoria del gene.
Recenti studi hanno dimostrato che alcune azioni degli steroidi e degli ormoni
tiroidei possono essere mediate da meccanismi non genomici. Infatti, alcuni recettori per gli estrogeni sono localizzati sulla membrana cellulare; inoltre recettori per
SEU
ROMA
14
Endocrinologia
la T3 sono stati riscontrati sulla superficie interna della membrana mitocondriale nel
fegato, nel rene e nel muscolo, tessuti nei quali gli ormoni tiroidei stimolano la fosforilazione ossidativa. I rapporti di eventuale cooperazione o inibizione tra le azioni
genomiche e non genomiche sono oggetto di approfonditi studi e non sono stati
ancora ben compresi.
Effetti fisiologici degli ormoni
SEU
Gli ormoni, interagendo con recettori localizzati a livello delle cellule bersaglio,
evocano risposte multiple e specifiche, regolando le attività enzimatiche, l’espressione genica e la sintesi proteica. Virtualmente tutti i tessuti dell’organismo sono sensibili agli ormoni e, sebbene ciascun ormone svolga funzioni ben definite, descritte
in dettaglio nei capitoli successivi, nel complesso gli effetti fisiologici degli ormoni
possono essere distinti in quattro categorie.
Sviluppo e crescita. L’influenza degli ormoni sullo sviluppo è evidente fin dalla
vita fetale, come dimostrato per esempio dalle alterazioni del sistema nervoso centrale dovute alla carenza di ormoni tiroidei. Nel periodo postnatale i principali ormoni
deputati al controllo della crescita sono il GH e l’IGF-1; contribuiscono in maniera
determinante anche la vitamina D, necessaria per la maturazione scheletrica, gli
steroidi sessuali, che inducono la saldatura delle epifisi, e gli ormoni tiroidei.
Produzione e conservazione di energia. L’insulina, il glucagone, il GH, le catecolamine, gli ormoni tiroidei e i glucocorticoidi regolano il metabolismo di carboidrati,
lipidi, proteine e acidi nucleici e promuovono la conversione dei composti introdotti con la dieta in energia utilizzabile immediatamente o in riserve. Questi ormoni
agiscono in maniera coordinata in modo da assicurare una risposta adeguata alle
esigenze dell’organismo, esigenze che variano per esempio in condizioni di digiuno
o di stress.
Riproduzione. Gli ormoni controllano lo sviluppo e la funzione delle gonadi e
contribuiscono in modo cruciale alla differenziazione anatomica, funzionale e comportamentale dei due sessi. Gli estrogeni, il progesterone e gli androgeni, regolati
dalle tropine ipofisarie, consentono la maturazione sessuale, l’acquisizione e il mantenimento della capacità riproduttiva, la gravidanza, l’espletamento del parto e la
lattazione.
Mantenimento dell’omeostasi. Gli ormoni sono responsabili del mantenimento
e della regolazione delle condizioni ottimali di tutti gli organi e apparati anche in rapporto alle modificazioni ambientali. Tutti i principali sistemi omeostatici (pressione
arteriosa, frequenza cardiaca, bilancio idroelettrolitico, equilibrio acido-base, temperatura corporea, composizione dei tessuti corporei), tranne la respirazione, sono
sottoposti a controllo ormonale.
ROMA
Sistemi di retroregolazione o di regolazione ormonale a feedback
Il feedback o meccanismo di retroregolazione è una caratteristica fondamentale
dei sistemi endocrini, mediante la quale gli ormoni controllano gli effetti biologici che
essi determinano e questi, a loro volta, regolano la secrezione degli stessi ormoni.
Il sistema di feedback consiste in un flusso bidirezionale continuo di informazioni tra
la sede di produzione dell’ormone e il tessuto bersaglio ed è pertanto fondamentale
per il mantenimento dell’omeostasi. Il feedback è definito positivo o negativo a se-
Principi di endocrinologia
15
conda che provochi una stimolazione o un’inibizione dell’azione ormonale. In base
alla distanza che intercorre tra l’ormone e il tessuto bersaglio, il feedback è distinto
in lungo, corto e ultracorto. Il feedback lungo è quello che si verifica tra ipotalamoipofisi e ghiandola bersaglio, con effetto sia stimolatorio che inibitorio. Un esempio è
il feedback esercitato dagli ormoni tiroidei sulla secrezione di TRH-TSH, Fig. 1.8: la
riduzione degli ormoni tiroidei circolanti induce un rapido incremento delle concentrazioni di TRH e TSH, che stimolano la ghiandola tiroidea a sintetizzare ormoni, con
lo scopo di normalizzare i valori sierici di T3 e T4; al contrario l’elevazione dei livelli
degli ormoni tiroidei sopprime la produzione di TRH e TSH. Il feedback corto si verifica invece tra gli ormoni ipofisari e i loro releasing hormones ipotalamici con effetto
sia stimolatorio che inibitorio; con questo meccanismo il TRH stimola la produzione
di TSH che, a sua volta, inibisce la secrezione di TRH, Fig. 1.8. Infine il feedback
ultracorto è quello per cui l’ormone controlla la sua secrezione inibendo, con meccanismo autocrino e paracrino, la funzione delle cellule che lo producono e di quelle
circostanti. La regolazione a feedback interessa anche sistemi che non coinvolgono
l’ipofisi: ad esempio il calcio con questo meccanismo modula la secrezione di PTH,
mentre la glicemia regola i livelli sierici di insulina.
L’esistenza dei sistemi di feedback riveste una grande importanza diagnostica e
la conoscenza di questi meccanismi di regolazione è fondamentale per la corretta interpretazione dei risultati delle indagini ormonali. Ad esempio, bassi valori di ormoni
tiroidei generalmente si associano ad un’elevazione del TSH (ipotiroidismo primario); quando ciò non accade bisogna considerare la presenza di un deficit tiroideo
secondario ad una patologia ipotalamo-ipofisaria (ipotiroidismo centrale). Inoltre la
maggior parte dei test dinamici (vedi il paragrafo Malattie endocrine) si basa sulla
valutazione dell’integrità dei sistemi di feedback, che risultano alterati in numerose
patologie endocrine.
SEU
ROMA
Figura 1.8 – Feedback lungo (gli ormoni tiroidei inibiscono la secrezione di TSH e TRH) e corto (il
TSH inibisce la secrezione di TRH).
16
Endocrinologia
Ritmi endocrini
Il sistema endocrino è regolato in modo dinamico con oscillazioni temporali variabili (da pochi minuti a 1 anno) delle secrezioni ormonali. Le variazioni regolari nel
tempo sono definite ritmi e possono essere rappresentate con un modello matematico sinusoidale, che consente di quantificare i ritmi biologici utilizzando 5 parametri:
il periodo, l’ampiezza, lo zenith, il nadir e il mesor. Il periodo di un ritmo è l’intervallo
di tempo che intercorre tra 2 episodi identici nel corso della variazione ed è quindi
una misura della frequenza delle variazioni dei livelli ormonali. A seconda della lunghezza del periodo i ritmi si dividono in ultradiani, circadiani e infradiani, Fig. 1.9. Il
ritmo ultradiano ha una durata inferiore alle 20 ore ed è caratteristico degli ormoni
ipofisari, che presentano oscillazioni episodiche della secrezione ad intervalli di 1-4
ore; il ritmo circadiano, con una durata compresa tra 20 e 28 ore, si verifica nella
secrezione della maggior parte degli ormoni; il ritmo infradiano presenta una durata
superiore alle 28 ore ed è tipico della secrezione delle gonadotropine durante il ciclo
mestruale. L’ampiezza del ritmo viene definita come la differenza tra il valore massimo e quello minimo. L’acrofase o zenith di un ritmo corrisponde al massimo dell’oscillazione, cioè al picco di secrezione, mentre il nadir corrisponde al minimo, cioè
al valore più basso della concentrazione dell’ormone. Il mesor (midline estimating
statistic of rhythm) corrisponde alla media aritmetica dei valori dell’ormone.
I ritmi endocrini, e più in generale i ritmi biologici, sono necessari per il mantenimento dell’omeostasi e sono espressione delle capacità adattative dell’organismo
alle variazioni ambientali (ciclo buio-luce) e alle esigenze fisiologiche (ciclo sonnoveglia), rappresentando una sorta di orologio interno che regola la sequenza temporale degli eventi fisiologici. In condizioni patologiche i ritmi endocrini, essenziali per
la normale funzione della maggior parte degli ormoni, sono invece alterati. Queste
alterazioni sono importanti dal punto di vista clinico e diagnostico perché permettono
la corretta interpretazione delle variazioni temporali dei valori ormonali. Ad esempio,
la secrezione di ACTH e di cortisolo è caratterizzata da un ritmo circadiano, con
valori massimi al mattino che si riducono gradualmente durante il resto del giorno raggiungendo livelli minimi nella notte; la perdita della ritmicità circadiana della
SEU
ROMA
Figura 1.9 – Ritmi endocrini.
Principi di endocrinologia
17
secrezione di cortisolo è un indice di ipercortisolismo più importante di un singolo
valore ormonale. La conoscenza del ritmo ormonale è rilevante anche dal punto di
vista terapeutico; infatti, nella terapia di un paziente con insufficienza surrenale, la
dose sostitutiva di cortisone deve essere somministrata rispettando il ritmo fisiologico dell’ormone, cioè 2/3 al mattino e 1/3 al pomeriggio. Al contrario, se si desidera
ottenere la soppressione dell’asse ipofisi-surrene, la dose di corticosteroidi deve
essere somministrata alla sera, prima dell’incremento circadiano dell’ACTH.
SEU
Ormoni e sistema immune
I processi infettivi, infiammatori, autoimmunitari, e più in generale tutti gli eventi
di danno tessutale sono caratterizzati dall’attivazione dell’asse ipotalamo-ipofisi-surrene, da modificazioni dei livelli plasmatici degli ormoni tiroidei, da inibizione della
funzione riproduttiva e da alterazioni del metabolismo dei carboidrati. Queste modificazioni rappresentano delle risposte non specifiche allo stress, all’aumentato catabolismo o alla riduzione dell’apporto calorico; i mediatori dell’infiammazione esercitano peraltro un’azione diretta sul sistema endocrino e viceversa. Infatti il sistema
endocrino e quello immune sono integrati attraverso un circuito bidirezionale mediante il quale gli ormoni e i neuropeptidi influenzano il sistema immune e la risposta
immune, a sua volta, modifica il sistema endocrino.
Effetti del sistema endocrino sul sistema immune. Il sistema endocrino produce ormoni e neuropeptidi che regolano la risposta immune attraverso specifici
recettori presenti sulle cellule del sistema immune. In linea generale i glucocorticoidi,
gli estrogeni, i progestinici e gli androgeni deprimono la risposta immune mentre il
GH, la PRL, il TSH, gli ormoni tiroidei, l’insulina e il CRH la stimolano. Il sistema immune produce peraltro ormoni e neuropeptidi la cui secrezione è controllata dall’ipofisi: peptidi correlati alla proopiomelanocortina, GH, PRL, TSH, LH, FSH e peptidi
derivati dalla proencefalina.
Gli ormoni che influiscono maggiormente sul sistema immunitario sono i glucocorticoidi, che per le loro proprietà antinfiammatorie ed immunosoppressive sono
ampiamente sfruttati in ambito clinico per il trattamento delle patologie autoimmuni e
delle malattie infiammatorie croniche.
Va rilevato che nella risposta immune dopo la pubertà si riscontrano differenze tra
i due sessi: in generale la risposta immune, sia umorale che cellulo-mediata, è più
attiva nei soggetti di sesso femminile rispetto a quelli di sesso maschile, ad eccezione del periodo della gravidanza. Queste differenze sono riconducibili al fatto che gli
estrogeni esercitano effetti sia inibitori che stimolatori sul sistema immune: inducono
un aumento dell’immunità umorale ma deprimono l’immunità cellulo-mediata riducendo l’attività dei linfociti T, l’attività natural-killer e la produzione di fattori timici che
stimolano la proliferazione dei linfociti T suppressor. Questi effetti determinano un
incremento della produzione anticorpale e rivestono pertanto un ruolo rilevante nella
patogenesi delle malattie autoimmuni, la cui prevalenza è infatti significativamente
maggiore nel sesso femminile. Si rileva, inoltre, che gli effetti immunosoppressivi
degli estrogeni e del progesterone sono responsabili della tolleranza immunologica
tipica della gravidanza, che impedisce la reazione immune contro il feto. Nel postpartum la caduta dei livelli di estrogeni e progesterone stimola la reattività immunologica, potendo determinare un’esacerbarsi delle malattie autoimmuni preesistenti
alla gravidanza.
ROMA
18
Endocrinologia
Effetti del sistema immune sul sistema endocrino. Il sistema immune influenza le cellule endocrine attraverso le citochine prodotte dalle cellule immuni attivate.
Le molecole che influiscono maggiormente sulle funzioni endocrine comprendono
l’interleuchina 1 (IL-1), l’interleuchina 2 (IL-2), l’interleuchina 6 (IL-6), il fattore di
necrosi tumorale α (TNFα) e l’interferone γ (IFNγ). Nel complesso il meccanismo
di feedback citochine/asse ipotalamo-ipofisi-surrene stimola la produzione di glucocorticoidi, che a loro volta limitano la risposta infiammatoria. L’asse ipotalamo-ipofisi-tiroide viene invece inibito dalle citochine infiammatorie. Inoltre la produzione di
IFN-γ da parte dei linfociti intratiroidei induce l’espressione aberrante di antigeni del
complesso MHC di classe II innescando il processo autoimmune. L’IL-1 a livello ipotalamico esercita un effetto inibitorio sulla secrezione di GnRH con un meccanismo
mediato dagli oppioidi endogeni, mentre a livello gonadico inibisce l’azione di FSH
ed LH e riduce la sintesi degli steroidi sessuali. Infine le citochine contribuiscono alla
sintesi di GH, di PRL, di vasopressina e di ossitocina.
SEU
Malattie endocrine
Le malattie endocrine, o endocrinopatie, sono caratterizzate da una carenza o da
un eccesso di ormoni, che nella maggior parte dei casi sono riconducibili, rispettivamente, a ipo o iperfunzione di una determinata ghiandola endocrina.
Prevalenza
La prevalenza varia in rapporto a fattori ambientali, genetici e sociali. Nei Paesi
occidentali le malattie endocrine rappresentano la seconda causa di consultazione
del medico; le endocrinopatie più frequenti comprendono le tireopatie, il diabete mellito, l’obesità, le dislipidemie, gli ipogonadismi, le patologie ipofisarie e quelle surrenali. Inoltre, nelle popolazioni ad elevato tenore socio-economico, le endocrinopatie
costituiscono la 7a causa di morte; va peraltro rilevato che la causa più frequente
di morte, cioè la malattia cardiovascolare nelle sue molteplici espressioni cliniche
(cardiopatia ischemia, ictus cerebri, vasculopatia periferica), rappresenta spesso
una complicanza del diabete e delle iperlipidemie ed è quindi significativamente
correlata alle malattie endocrino-metaboliche. Nei Paesi del Terzo Mondo la patologia endocrina più frequente è il gozzo, che in alcune aree colpisce fino all’80% della
popolazione generale, mentre le altre endocrinopatie sono più rare, data l’elevata
prevalenza delle malattie infettive e di quelle da carenza alimentare.
ROMA
Classificazione
Endocrinopatie da insufficienza ormonale
La carenza ormonale si manifesta con una sindrome clinica da insufficienza endocrina della ghiandola corrispondente. Nella maggior parte dei casi l’alterazione
è dovuta ad una ridotta produzione ormonale, mentre più raramente si verifica una
resistenza tessutale all’azione dell’ormone.
1. Distruzione o assenza della ghiandola endocrina. Il deficit ormonale può
essere dovuto ad ipo-agenesia o disgenesia, a distruzione autoimmune (dia-
Principi di endocrinologia
19
bete mellito di tipo 1, ipotiroidismo da tiroidite di Hashimoto, ecc.), a danno iatrogenico (chirurgico, radiante o farmacologico), ad infarto (ad es. apoplessia
ipofisaria), a distruzione flogistica o neoplastica.
2. Deficit della sintesi ormonale. I difetti dell’ormonogenesi possono essere
dovuti ad alterazioni del gene che codifica per l’ormone carente (ad es. deficit
di GH), a deficit di attività enzimatiche (ad es. alterazioni della steroidogenesi
surrenale e dell’ormonogenesi tiroidea), a farmaci che interferiscono con la
biosintesi ormonale o a carenze alimentari (ipotiroidismo da insufficiente apporto di iodio, deficit di vitamina D).
3. Aumento del catabolismo ormonale. L’insufficienza ormonale può essere
indotta da farmaci che stimolano il metabolismo ormonale (ad es. la fenitoina
e gli ormoni tiroidei aumentano il catabolismo dei glucocorticoidi).
4. Resistenza ormonale. Può essere dovuta a deficit recettoriali (ridotto numero di recettori o diminuita affinità del recettore per l’ormone) o postrecettoriali
(alterazioni del sistema di trasduzione del segnale ormonale a valle del recettore). L’insufficiente azione ormonale a livello degli organi bersaglio comporta
un’alterazione del meccanismo di feedback con aumento della produzione
dell’ormone, i cui livelli circolanti sono pertanto normali o elevati. La riduzione
del numero di recettori può essere dovuta anche alla somministrazione di
ormoni esogeni, come si osserva nei pazienti trattati con analoghi del GnRH
a lunga durata d’azione, nei quali si verifica inizialmente un aumento della
produzione di gonadotropine, seguito da una diminuzione della risposta ormonale e castrazione funzionale. L’azione dell’ormone può essere compromessa anche dalla produzione di anticorpi anti-ormone (deficit pre-recettoriale),
come accade in presenza di anticorpi che bloccano il recettore dell’insulina
impedendone l’azione sulle cellule bersaglio. La resistenza ormonale comprende anche i casi di alterata trasformazione dei precursori ormonali nella
forma attiva: ad esempio il deficit della 5α−reduttasi comporta l’incapacità di
trasformare il testosterone in diidrotestosterone, metabolicamente più attivo,
con conseguente ipoandrogenismo tessutale e pseudoermafroditismo maschile. Un altro esempio riguarda il deficit dell’enzima renale che idrossila la
25(OH)D in posizione 1α con conseguente mancata produzione dell’ormone
attivo 1,25(OH)2D.
SEU
ROMA
Endocrinopatie da eccesso ormonale
Le sindromi da eccesso ormonale sono riconducibili ad un’eccessiva presenza di
ormoni a livello delle cellule bersaglio, dovuta a diversi meccanismi.
1. Iperfunzione primaria della ghiandola endocrina. In genere è dovuta ad
un’alterazione primitiva della ghiandola che produce l’ormone. Nella maggior
parte dei casi è causata dall’iperplasia o dalla proliferazione neoplastica, benigna o maligna, di cellule endocrine (adenomi ipofisari, noduli tiroidei autonomi,
iperaldosteronismo, iperparatiroidismo primario, insulinoma, ecc.). Nella malattia di Basedow-Graves l’iperproduzione ormonale è dovuta ad autoanticorpi
diretti contro il recettore del TSH che mimano l’azione dell’ormone tireotropo
stimolando la ghiandola tiroidea.
2. Iperfunzione secondaria da eccesso di tropine ipofisarie. L’iperfunzione
ghiandolare è secondaria ad iperplasia o adenoma ipofisario con iperproduzione di una determinata tropina ipofisaria ed iperstimolazione della ghiandola
20
Endocrinologia
3.
4.
5.
6.
7.
bersaglio (ad es. ipercorticosurrenalismo da adenoma ACTH-secernente o
malattia di Cushing).
Produzione ectopica ormonale. La produzione ectopica è dovuta a tumori
derivanti da cellule che normalmente non producono l’ormone (ad es. sindrome
di Cushing da iperproduzione di ACTH da parte di un microcitoma polmonare).
Ipersensibilità tessutale. È dovuta ad un aumento del numero o dell’affinità
dei recettori per l’ormone. In alcuni casi sono state riscontrate mutazioni che
attivano il recettore o il sistema di trasduzione del segnale indipendentemente
dal legame con l’ormone (ad es. attivazione costitutiva della subunità α della
proteina G nella sindrome di McCune-Albrigth)
Riduzione del catabolismo ormonale. Può essere correlata all’assunzione
di farmaci o a condizioni patologiche intrinseche all’organismo (ad es. iperprolattinemia dovuta a ridotta eliminazione nei pazienti con insufficienza renale).
Iperproduzione periferica dei precursori. L’eccesso ormonale è dovuto all’iperproduzione da eccessiva trasformazione periferica dell’ormone attivo a
partire dai suoi precursori (ad es. iperestrogenismo da trasformazione dell’androstenedione in estrogeni nelle epatopatie).
Eccesso di somministrazione esogena. L’eccesso ormonale è dovuto
all’eccessiva somministrazione di ormoni o sostanze con azione ormonale
(sindrome di Cushing da terapia steroidea, somministrazione di anabolizzanti,
GH o eritropoietina negli atleti).
Diagnosi
SEU
Il quadro clinico associato alle endocrinopatie varia in rapporto alla gravità e alla
durata della disfunzione endocrina. Inoltre talvolta si verificano meccanismi di compensazione per cui la malattia si manifesta soltanto in una fase molto avanzata,
come accade nell’ipotiroidismo da deficit dell’ormonogenesi tiroidea, nel quale la
ridotta produzione di tiroxina determina un’ipersecrezione di TSH con conseguente ipertrofia compensatoria della tiroide e formazione di gozzo. Alcune sindromi da
disfunzione endocrina, ad esempio la tireotossicosi grave, presentano una sintomatologia evidente fin dall’inizio, ma spesso le endocrinopatie esordiscono lentamente
e le prime manifestazioni cliniche sono lievi, sfumate e non facilmente riscontrabili
all’anamnesi e all’esame obiettivo, per cui la malattia può essere svelata soltanto
con opportune indagini di laboratorio.
ROMA
Anamnesi ed esame obiettivo
L’accertamento clinico dello stato endocrino di un paziente si fonda sugli elementi
emersi da un’anamnesi approfondita, familiare e personale, fisiologica e patologica,
e da un esame obiettivo accurato. Bisogna comunque considerare che molti sintomi
e segni associati alla malattie endocrine sono spesso dovuti a cause non endocrine.
Inoltre, dato che gli ormoni influenzano la funzione di tutti i tessuti e di tutti gli organi,
gli aspetti clinici associati alle malattie endocrine sono estremamente vari e possono
essere generalizzati o localizzati.
Indagini di laboratorio
Il sospetto diagnostico deve in ogni caso essere confermato attraverso opportune indagini di laboratorio.
Principi di endocrinologia
21
Determinazione dei valori ormonali basali nel sangue e/o nelle urine. Lo
studio della funzione di una data ghiandola endocrina prevede innanzitutto la determinazione dell’ormone prodotto e/o dei suoi metaboliti nel plasma e/o nelle urine in condizioni basali. Infatti la diagnosi di endocrinopatia è spesso basata sulla
dimostrazione di valori ormonali aumentati o diminuiti, tenendo comunque presente che per la corretta interpretazione dei risultati bisogna considerare anche l’età,
il sesso, l’ora del prelievo, il ritmo circadiano, l’esercizio fisico, lo stato emotivo, la
funzione epatica e renale, la presenza di altre malattie e l’eventuale assunzione
di farmaci.
Le indagini ormonali su sangue permettono di dosare i livelli dell’ormone in un determinato momento, pertanto nel caso degli ormoni con lunga emivita (per esempio
la tiroxina) consentono di accertare lo stato funzionale ghiandola con buona attendibilità, mentre per gli ormoni con breve emivita (come l’adrenalina o l’insulina) l’indagine fornisce informazioni solo sulla quantità di ormone presente al momento del
prelievo. Pertanto un feocromocitoma che rilascia adrenalina in maniera episodica è
associato ad elevati livelli plasmatici dell’ormone solo nel periodo dell’immissione in
circolo dell’adrenalina. La sindrome di Cushing può essere associata ad un’aumentata frequenza dei picchi del cortisolo con livelli plasmatici normali dell’ormone tra i
picchi. Per gli ormoni che vengono secreti e rilasciati in maniera episodica è quindi
necessario determinare più volte l’ormone su prelievi distanziati nel tempo oppure
misurarlo su un pool di campioni prelevati a distanza di 30 minuti l’uno dall’altro.
Molti ormoni, come la tiroxina e gli steroidi, circolano legati a proteine di trasporto
e quindi il dosaggio delle frazioni totali è indicativo solo se non esistono anomalie
quantitative o qualitative delle proteine di trasporto; pertanto il dosaggio delle frazioni libere degli ormoni fornisce risultati più attendibili. Va infine notato che i valori
plasmatici di numerosi ormoni oscillano in uno scarto (range) anche molto ampio,
per cui è bene determinare non solo l’ormone di interesse ma anche quello che lo
regola tramite il meccanismo di feedback perché il loro rapporto è costante e quindi
all’elevazione di uno corrisponde l’abbassamento dell’altro e viceversa, con l’eccezione delle sindromi da resistenza.
La determinazione urinaria di un ormone o dei suoi metaboliti riflette i valori plasmatici dell’ormone e la sua secrezione ghiandolare e in genere si effettua sulle
urine totali delle 24 ore. Il dosaggio urinario in alcuni casi è vantaggioso rispetto a
quello plasmatico in quanto indica la quantità totale di ormone secreto nelle 24 ore,
un dato molto utile per la valutazione degli ormoni secreti in maniera episodica e dotati di una breve emivita. Affinché la determinazione sia valida deve essere misurata
la creatinina urinaria, che conferma l’adeguatezza della raccolta urinaria. Si ricorda
che le alterazioni della funzione renale inoltre possono modificare l’escrezione urinaria dell’ormone e quindi fuorviare la diagnosi.
Indagini dinamiche (di stimolo e di soppressione). Quando la disfunzione
endocrina non è ancora così evidente da modificare i valori ormonali basali è necessario ricorrere alle prove dinamiche di stimolo o di inibizione, basate sulla somministrazione di sostanze che modificano la produzione endogena dell’ormone in studio.
Infatti queste indagini permettono di evidenziare il significato di valori ai limiti della
norma: le prove di stimolo evidenziano condizioni di insufficienza e quelle di inibizione rilevano stati di iperfunzione. I test di stimolo inoltre consentono di distinguere
un’insufficienza endocrina primaria da una secondaria e di determinare la riserva
endocrina della ghiandola bersaglio.
SEU
ROMA
22
Endocrinologia
Determinazione degli autoanticorpi. In alcune endocrinopatie la determinazione di anticorpi diretti verso ormoni o tessuti endocrini è fondamentale per stabilire la
diagnosi. Gli anticorpi possono essere stimolanti, inibenti o bloccanti e nello stesso
individuo possono coesistere più tipi di anticorpi.
Indagini per immagini
In alcuni casi è fondamentale sia per la diagnosi che per la localizzazione dell’endocrinopatia. L’ecografia consente una visualizzazione bidimensionale della struttura esplorata identificando formazioni solide di almeno 3-4 mm e lesioni cistiche di 2
mm. La tomografia computerizzata (TC) e la risonanza magnetica (RM) permettono
di individuare lesioni di 2-3 mm e di distinguere l’iperplasia dal tessuto normale. La
TC è un’indagine basata sull’impiego dei raggi X che consente l’elaborazione elettronica dei dati fornendo immagini degli strati trasversali della struttura esaminata;
il suo potere analitico è cento volte superiore rispetto a quello della radiografia convenzionale. La RM fornisce immagini orientate sui piani assiale, frontale e sagittale.
Si tratta di una tecnica fondata sul principio della risonanza emessa dai nuclei atomici sollecitati da impulsi ad alta energia all’interno di un campo magnetico; la RM si
avvale pertanto dell’interazione di un campo magnetico esterno con quello intrinseco
delle strutture corporee. La scintigrafia è basata sull’assunzione e sulla dismissione
di un radioisotopo (ad es. radioiodio, pertecnetato, MIBG) e fornisce informazioni sia
morfologiche che funzionali della ghiandola in esame.
Terapia
SEU
Il trattamento dei deficit ormonali si avvale della terapia endocrina sostitutiva,
che tuttavia non è sempre di facile attuazione a causa della mancata disponibilità dell’ormone carente o della difficile somministrazione di questo, oppure perché
non è possibile evitare l’instaurarsi di complicanze. Gli ormoni polipeptidici impiegati
per il trattamento sostitutivo sono attualmente sintetizzati in vitro attraverso le tecniche del DNA ricombinante, che permettono di produrre su scala industriale proteine
identiche alla sostanza nativa. Rispetto alle preparazioni estrattive animali (insulina)
o umane (GH) precedentemente impiegate, le molecole sintetizzate in laboratorio
non solo offrono il vantaggio di avere una disponibilità praticamente illimitata e a
basso costo, ma sono anche più sicure e presentano una minore immunogenicità.
L’ingegneria genetica peraltro consente di ottenere proteine parzialmente modificate
rispetto alla molecola nativa, in modo da avere caratteristiche farmacocinetiche differenti mantenendo invariata l’affinità per il recettore; ciò offre notevoli vantaggi clinici, come evidenziato dalla possibilità di impiegare diversi tipi di insulina a seconda
della situazione clinica specifica. Alcuni ormoni sintetizzati mediante le biotecnologie
sono impiegati anche a fini diagnostici.
In caso di iperfunzione la terapia si pone come obiettivo quello di ridurre la sintesi
ormonale o di diminuirne l’azione periferica dell’ormone prodotto in eccesso: a tale
scopo possono essere impiegati appositi presidi farmacologici oppure, in alcuni casi,
si ricorre alla rimozione chirurgica o radiante della causa dell’eccesso ormonale,
rappresentata in genere da un tumore, da un’iperplasia o da una ghiandola intera
affetta da una malattia autoimmune (malattia di Basedow-Graves).
ROMA
Principi di endocrinologia
23
Terapia ormonale in malattie non endocrine e nello sport
Le molteplici azioni degli ormoni hanno permesso il loro impiego anche nel trattamento di malattie non endocrine. Gli ormoni più frequentemente utilizzati sono i
glucocorticoidi, dotati di una potente azione antinfiammatoria. Altri farmaci ampiamente utilizzati nella pratica clinica sono gli antiandrogeni nel trattamento del carcinoma prostatico, i modulatori selettivi del recettore degli androgeni (tamoxifene)
nelle donne affette da carcinoma mammario e gli analoghi del GnRH a lunga durata
d’azione, usati in entrambe queste condizioni per indurre una castrazione funzionale
e minimizzare gli effetti degli steroidi sessuali sulla progressione tumorale.
Va infine ricordato che numerosi ormoni (GH, IGF, eritropoietina, steroidi anabolizzanti, insulina) sono utilizzati dagli atleti per migliorare le prestazioni agonistiche;
purtroppo tale impiego è ingiustificato perché induce dei danni spesso irreversibili
per l’organismo, determinando in alcuni casi complicazioni anche mortali. In questa
sede sono riportati gli aspetti fondamentali riguardanti l’uso dell’eritropoietina, secreta dal rene; per la trattazione degli altri ormoni usati illegalmente nello sport si
rimanda ai capitoli specifici.
SEU
Eritropoietina: impiego clinico e nel doping
L’eritropoietina (EPO) è un ormone glicoproteico di origine renale che promuove
la produzione dei globuli rossi e la cui secrezione è regolata da un sistema di feedback: l’ipossia aumenta la produzione di EPO e di conseguenza anche quella dei
globuli rossi, mentre l’iperossia riduce le concentrazioni dell’ormone. Le principali
indicazioni per l’impiego clinico dell’EPO comprendono l’anemia secondaria ad insufficienza renale e quella che compare in pazienti affetti da cancro o in conseguenza di trattamenti radio- e chemioterapici. Dato che l’EPO naturale non può essere
facilmente purificata in quantità rilevanti, questo ormone viene prodotto con le tecniche dell’ingegneria genetica. L’EPO ricombinante in realtà non è costituita da una
singola proteina, ma da una miscela di sialoproteine con residui zuccherini attaccati
in varia maniera a 4 potenziali siti. In ambito terapeutico viene utilizzata anche la
darbopoietina, una proteina prodotta mediante tecnologia ricombinante che differisce dall’EPO in cinque porzioni; la darbopoietina ricombinante ha due siti aggiuntivi
per l’attacco di due carboidrati e pertanto è composta da un numero maggiore di
entità chimiche. L’EPO è in genere somministrata per via sottocutanea, ma anche
endovenosa, 3 volte a settimana ad una dose iniziale complessiva di 200 UI; la darbopoietina è invece somministrata in un’unica dose settimanale di 1 μg. Il dosaggio
viene modificato in base al valore dell’emoglobina.
Dagli anni ’80 è iniziato l’impiego di EPO anche a scopo di doping, soprattutto
negli atleti sottoposti a sforzi fisici prolungati (ciclisti, maratoneti, ecc.). La finalità
dell’abuso di EPO in ambito sportivo è quella di aumentare la massa circolante degli
eritrociti e quindi la capacità di trasportare l’ossigeno ai tessuti ed in particolare ai
muscoli. Spesso all’EPO viene associato l’impiego di broncodilatatori, che aumentano lo scambio gassoso a livello polmonare. Il Comitato Olimpico Internazionale ha
proibito l’uso di EPO in manifestazioni ufficiali a partire dal 1989; per identificare le
eritropoietine ricombinanti usate dagli atleti vengono impiegate tecniche che sfrutta-
ROMA
24
Endocrinologia
no le differenze nell’attacco dei carboidrati e quindi il diverso numero di molecole di
acido sialico rispetto all’EPO naturale. Ovviamente il limite di queste determinazioni
è correlato all’emivita urinaria dei farmaci impiegati (48 ore per l’EPO e 1 settimana
per la darbopoietina).
Va rilevato che l’impiego dell’EPO, sia a fini terapeutici che nello sport, non è
scevro da rischi e complicanze anche gravi, che comprendono ipertensione, mialgia, trombosi, incidenti cerebrovascolari e infarto miocardio. Una complicanza rara è
l’aplasia specifica della serie rossa, dovuta alla formazione di anticorpi anti-EPO. È
da rilevare che la maggior parte dei dati disponibili è comunque riferita a popolazioni
di pazienti e pertanto non sono direttamente confrontabili con i potenziali rischi dell’impiego dell’EPO negli atleti.
Bibliografia
SEU
Borrione P, Mastrone A, Salvo RA, et al, Oxygen delivery enhancers: past, present, and future, J Endocrinol Invest, 2008, 31(2), 185.
Buonamano A e Romagnani P, Doping con eritropoietina, l’Endocrinologo, 2003, 4, 90.
Federman DD, The endocrine patient, in “Williams Textbook of Endocrinology”, XI Ed, Kronenberg HM, Melmed S, Polonsky KS, Larsen PR Eds, Saunders Elsevier, Philadelphia,
2008, 12.
Gaudard A, Varlet-Marie E, Bressolle F, Audran M, Drugs for increasing oxygen and their potential use in doping: a review, Sports Med, 2003, 33(3), 187.
Kronenberg HM, Melmed S, Larsen PR, Polonsky KS, Principles of endocrinology, in “Williams Textbook of Endocrinology”, XI Ed, Kronenberg HM, Melmed S, Polonsky KS, Larsen
PR Eds, Saunders Elsevier, Philadelphia, 2008, 3.
Lasne F, Martin L, Crepin N, de Ceaurriz J, Detection of isoelectric profiles of erythropoietin
in urine: differentiation of natural and administered recombinant hormones, Anal Biochem,
2002, 311, 119.
Lazar MA, Mechanism of action of hormones that act on nuclear receptor, in “Williams Textbook of Endocrinology”, XI Ed, Kronenberg HM, Melmed S, Polonsky KS, Larsen PR Eds,
Saunders Elsevier, Philadelphia, 2008, 47.
ROMA
Monaco F, Malattie endocrine, in “Endocrinologia Clinica”, IV Ed, Monaco F Ed, SEU, Roma,
2007, 55.
Napolitano G, Bucci I, Consoli A, Monaco F, Principi di endocrinologia, in “Endocrinologia
Clinica”, IV Ed, Monaco F Ed, SEU, Roma, 2007, 1.
Spiegel A, Carter-Su C, Taylor SI, Mechanism of action of hormones that act at the cell surface, in “Williams Textbook of Endocrinology”, XI Ed, Kronenberg HM, Melmed S, Polonsky KS,
Larsen PR Eds, Saunders Elsevier, Philadelphia, 2008, 47.