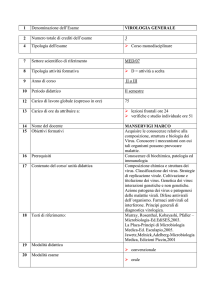
29
BOVINI
Large Animals Review, Anno 10, n. 2, Aprile 2004
MALATTIA DELLE MUCOSE
DA BVDV TIPO 2:
DESCRIZIONE DI UN FOCOLAIO IN PUGLIA
NICOLA DECARO1, MICHELE CAMERO1, GABRIELLA ELIA1, VITO MARTELLA1,
ANNAMARIA PRATELLI1, PIETRO GARGANO2, SEBASTIANO TINELLI3,
GIOVANNI LEOGRANDE3, CANIO BUONAVOGLIA1
1
Dipartimento di Sanità e Benessere Animale - Facoltà di Medicina Veterinaria di Bari
Strada per Casamassima Km 3 - 70010 Valenzano (BA)
2
Istituto Zooprofilattico Sperimentale del Mezzogiorno - Sezione di Cosenza
3
Veterinario Libero Professionista
Riassunto
Gli autori descrivono un focolaio di infezione acuta sostenuta da uno stipite del virus della diarrea virale bovina tipo 2 (BVDV-2) in un allevamento di bovine da latte della Puglia. Quattro vitelloni di 10 mesi di età hanno sviluppato
una grave forma morbosa, caratterizzata da febbre elevata, diarrea emorragica e fenomeni emorragico-necrotici a livello della mucosa della cavità orale e nasale, e sono morti a distanza di 6-7 giorni dalla comparsa delle manifestazioni cliniche.
L’isolamento virale, effettuato a partire da campioni d’organo prelevati da uno dei 4 animali morti, ha evidenziato la
presenza di uno stipite BVDV citopatogeno, che è stato caratterizzato come BVDV tipo 2 in base ai risultati della tipizzazione mediante nested-PCR e dell’analisi di sequenza. Anticorpi specifici per BVDV sono stati messi in evidenza in
51 dei 55 bovini allevati: dei 4 animali sieronegativi, 2 sono stati successivamente identificati come persistentemente
infetti in base ai risultati degli esami virologici effettuati sul sangue. Da questi soggetti, un vitello di 6 mesi ed una
bovina di 1 anno, è stato isolato uno stipite BVDV-2 non citopatogeno, con identità nucleotidica, a livello del gene
Erns, del 100% rispetto allo stipite citopatogeno isolato dal vitellone morto. Le caratteristiche cliniche ed epidemiologiche del focolaio descritto farebbero pensare ad una infezione da BVDV-2 trombocitopenico. Tuttavia, l’isolamento di
uno stipite citopatogeno BVDV tipo 2, mai descritto in precedenza come responsabile di sindrome emorragica, e l’identificazione in allevamento di soggetti con infezione persistente depongono per un episodio atipico di malattia delle
mucose.
Summary
A severe outbreak of bovine viral diarrhoea virus type 2 (BVDV-2) infection in a dairy cattle herd of Apulia (Italy) is
reported. Four calves, 10 months of age, developed a severe illness with fever, haemorrhagic diarrhoea, haemorrhages and necrosis of the oro-nasal mucosa, and died within 6-7 days from the onset of the clinical signs.
A BVDV cytopathogenic strain was isolated from tissue samples collected from a dead calf and was characterized
as BVDV type 2 by both nested-PCR and sequence analysis. BVDV antibodies were detected in 51 out of 55 bovines:
2 (a 6-month-old calf and a 12-month-old heifer) of the 4 seronegative animals were found BVDV persistently infected by virological investigations carried out on blood samples. The BVDV-2 non cytopathogenic strain isolated from
these animals showed a 100% nucleotide identity (E rns gene) to the cytopathogenic strain isolated from the dead calf.
Clinical and epidemiological pattern of the outbreak might indicate an acute infection by a BVDV-2 thrombocytopenic
strain. However, the isolation of a BVDV-2 cytopathogenic strain, which has never been associated to the haemorrhagic syndrome, and the detection of persistently infected animals in the same herd lead to the diagnosis of a severe
outbreak of mucosal disease.
30
Malattia delle mucose da BVDV tipo 2: descrizione di un focolaio in Puglia
INTRODUZIONE
Il virus della diarrea virale bovina (BVDV), isolato per
la prima volta nel 1946 da bovini con diarrea, febbre e leucopenia (Olafson et al., 1946; Olafson e Rickard, 1947), è
associato a varie forme morbose che comprendono infezioni subcliniche, immunodepressione, forme respiratorie,
forme intestinali e disturbi della sfera riproduttiva (ritorni
in calore, aborti, mummificazione fetale) (Baker, 1995).
BVDV appartiene alla famiglia Flaviviridae, genere Pestivirus, che comprende il virus della peste suina classica
(CSFV), il virus della border disease (BDV) e due distinti
genotipi di BVDV, BVDV tipo 1 e BVDV tipo 2 (Ridpath
et al., 1994; Heinz et al., 2000). Si tratta di virus ad RNA
monocatenario, a polarità positiva, codificante per una poliproteina, che è scissa, ad opera di proteasi cellulari e virali, in proteine strutturali (C, Erns, E1, E2) e non strutturali
(Npro, NS2-3, NS4A, NS4B, NS5A, NS5B) (Thiel et al.,
1996; Heinz et al., 2000). Entrambi i genotipi, BVDV 1 e
BVDV 2, possono esistere in due diversi biotipi, citopatogeno (cp) e non citopatogeno (ncp). Stipiti non citopatogeni particolarmente virulenti di BVDV 2, detti trombocitopenici, sono associati ad una patologia grave, chiamata
in passato sindrome emorragica (HS) e, più recentemente,
diarrea virale bovina acuta grave (SA-BVD), la quale è caratterizzata da febbre elevata, trombocitopenia, diatesi
emorragica, leucopenia, diarrea emorragica e morte
(Rebhun et al., 1989; Corapi et al., 1990; Pellerin et al.,
1994; Ridpath, 2003). In Italia, la circolazione di BVDV tipo 2 è documentata fin dal 1990 (Buonavoglia et al., 1991)
ed interessa sia la popolazione ovina (Pratelli et al., 2001)
che bovina (Giangaspero et al., 2001; Luzzago et al.,
2001). Tuttavia, è stato descritto un solo episodio di infezione da virus trombocitopenico, in bovini vaccinati con
un vaccino contaminato da uno stipite BVDV tipo 2 altamente virulento (Falcone et al., 2000).
L’infezione di una bovina gravida con uno stipite BVDV
ncp ha, generalmente, esiti diversi a seconda dell’epoca di
gestazione. Nelle prime settimane di gravidanza si possono
verificare riassorbimento embrionale, con conseguente ritorno in calore, aborto o mummificazione fetale, mentre
l’infezione nell’ultimo stadio di gestazione può esitare in
malformazioni fetali o nella nascita di vitelli perfettamente
sani e sieropositivi (Baker, 1995).
Tuttavia, ai fini epidemiologici risulta particolarmente importante l’infezione tra il terzo ed il quarto mese di gravidanza, quando il feto non ha ancora raggiunto l’immunocompetenza. In queste condizioni, il virus diffonde liberamente nei tessuti fetali e, quando il sistema immunitario
raggiunge la piena maturazione, riconosce come self il virus
e non produce anticorpi specifici. Di conseguenza, si assiste
alla nascita di vitelli immunotolleranti nei confronti di
BVDV, perché sieronegativi, e persistentemente infetti (PI).
Questi soggetti rappresentano il serbatoio dell’infezione ed
eliminano costantemente BVDV attraverso secreti ed escreti
(Brownlie, 1990; Baker, 1995; Moenning e Liess, 1995).
I vitelli PI, in particolari condizioni, possono sviluppare
la malattia delle mucose (MD), una patologia sporadica ad
esito letale, che si manifesta per lo più in animali di età
compresa tra 6 mesi e 2 anni. La MD è caratterizzata da
febbre elevata, diarrea, a volte emorragica, disturbi respiratori ed ulcerazioni estese su mucosa orale, esofagea e ga-
strointestinale, con morte entro 1-2 settimane dall’esordio
della sintomatologia (Baker, 1987; Moenning e Plagemann, 1992). L’insorgenza della MD in bovini PI è legata
all’acquisizione della citopatogenicità da parte dello stipite
ncp responsabile dell’infezione persistente. L’attitudine citopatogena è correlata all’espressione della proteina virale
NS3, che rappresenta un segnale apoptotico per le cellule
infette. Nel biotipo ncp questa proteina non strutturale è
parte integrante del complesso NS2-3 e non esplica attività
citolitica, mentre nel biotipo cp la proteina NS3, grazie a
meccanismi biomolecolari alquanto complessi, è libera e
determina apoptosi cellulare (Meyers e Thiel, 1996).
Nella presente nota si descrive un focolaio di infezione
acuta sostenuta da uno stipite BVDV-2 in un allevamento
di bovine da latte della Puglia.
MATERIALI E METODI
Animali
I casi clinici sono stati osservati in un allevamento di bovine da latte della provincia di Taranto (Puglia). All’epoca
dell’insorgenza del focolaio l’effettivo comprendeva 59 bovini: 32 vacche in produzione (25 brune, 6 frisone ed 1 jersey), 9 manze (6 brune e 3 frisone), 7 vitelli di 2 mesi (4
bruni, 1 frisone, 1 jersey e 1 incrocio), 4 vitelli bruni di circa 6 mesi di età, 6 vitelloni all’ingrasso di 10 mesi (2 bruni,
2 frisoni e 2 incroci), 1 bruno di 18 mesi destinato alla fecondazione delle bovine presenti in azienda. Le diverse categorie di animali erano allevate in locali separati.
In Giugno 2003, a distanza di pochi giorni l’uno dall’altro, 4 dei 6 vitelloni all’ingrasso hanno manifestato una
grave sintomatologia, caratterizzata inizialmente da febbre
(39,5°C) e diarrea emorragica; l’ampolla rettale si presentava ripiena di feci miste a sangue e materiale necrotico. A
distanza di 2-3 giorni dalla comparsa dei primi segni clinici, si potevano apprezzare, sulla mucosa orale, emorragie
diffuse ed erosioni ricoperte da materiale fibrinoso, con
prevalente localizzazione a livello delle gengive (Fig. 1),
palato duro (Fig. 2), superficie interna delle guance. Successivamente le emorragie comparivano anche sulla mucosa delle cavità nasali e sul musello (Fig. 3), con presenza di
scolo nasale mucoso (Fig. 4). A livello oculare si evidenziavano congiuntivite e scolo siero-mucoso. Gli animali colpiti sono stati trattati con antibiotici e soluzioni reidratanti,
ma, nonostante la terapia, non hanno presentato alcun miglioramento e sono morti a distanza di 6-7 giorni dall’esordio della sintomatologia.
Poco prima della morte, il veterinario aziendale ha effettuato un prelievo di sangue per la raccolta del siero da un
vitellone con sintomatologia in atto, mentre, dopo la morte, dallo stesso animale sono stati prelevati campioni di
polmone, fegato e milza per gli accertamenti di laboratorio. Non è invece stato possibile disporre di un campione
di sangue intero per gli esami ematologici.
L’indagine epidemiologica ha accertato che nell’estate
del 2002 in allevamento erano state introdotte 4 bovine di
razza frisona, di cui 2 erano gravide ed hanno partorito vitelli sani. Inoltre, circa un anno prima erano state acquistate 2 frisone gravide, una delle quali è stata identificata come madre di uno degli animali malati.
31
BOVINI
Large Animals Review, Anno 10, n. 2, Aprile 2004
FIGURA 1 - Emorragie ed erosioni sulla mucosa gengivale.
FIGURA 3 - Emorragie su musello e cavità nasali.
FIGURA 2 - Emorragie sul palato duro.
FIGURA 4 - Scolo nasale mucoso.
Isolamento virale
Frammenti di ciascun organo da esaminare sono stati
omogenati in terreno minimo essenziale di Dulbecco (DMEM) (10% p/v); l’omogenato è stato centrifugato a 4.000
x g per 15 min ed il surnatante di ciascun campione, opportunamente trattato con antibiotici, è stato inoculato su monostrati di cellule di polmone embrionale bovino (PEB). Le
colture cellulari sono state osservate quotidianamente al microscopio rovesciato per la evidenziazione dell’eventuale effetto citopatico; dopo 5 giorni di incubazione, monostrati infetti sviluppati su vetrini sono stati sottoposti a test di immunofluorescenza indiretta (IFI), utilizzando anticorpi monoclonali panpestivirus ed un siero anti-IgG di topo marcato
con fluoresceina (Sigma-Aldrich, Milano).
Caratterizzazione dello stipite pestivirus
Nested-PCR
Dal criolisato delle cellule infette e da ciascun organo è
stato estratto l’RNA virale mediante RNeasy Total RNA kit
(Qiagen GmbH, Hilden, Germania). Su ciascun estratto è
stato effettuato un test RT-PCR panpestivirus, utilizzando
il kit GeneAmp® RNA PCR (Applied Biosystems, Applera
Italia, Monza) e la coppia di primers P1/P2, in grado di
amplificare un frammento del gene della proteina strutturale Erns di 826 basi appaiate (bp) di BVDV tipo 1, BVDV
tipo 2 e BDV (Sullivan e Akkina, 1995). Sono state utilizzate le seguenti condizioni termiche: trascrizione inversa
(RT) a 42°C per 30 min, inattivazione della MuLV Reverse
Transcriptase a 99°C per 5 min, attivazione della Amplitaq
Gold DNA polymerase a 94°C per 10 min, 40 cicli di 94°C
per 1 min (denaturazione), 55°C per 1 min (annealing),
72°C per 1 min (polimerizzazione), seguiti da una estensione finale a 72°C per 10 min.
Un’aliquota (2 µl) della diluizione 1:500 del prodotto PCR
è stata sottoposta ad un ulteriore processo di amplificazione
(nested-PCR), utilizzando il primer antisenso P2 ed un set di
primers senso (TS1, TS2, TS3) tipo specifici, in grado cioè di
legarsi in maniera selettiva a BVDV tipo 1, BVDV tipo 2 o
BDV e di produrre amplificati di ampiezza diversa a seconda
del genotipo virale (Sullivan e Akkina, 1995). Le condizioni
termiche erano le seguenti: 94°C per 10 min, 30 cicli di 94°C
per 1 min, 50°C per 45 sec, 72°C per 45 sec, seguiti da una
fase di polimerizzazione finale a 72°C per 10 min.
Aliquote (8 µl) dei prodotti RT-PCR e nested sono state
analizzate mediante corsa elettroforetica a 60 V per 90 min
e visualizzazione al transilluminatore UV dopo colorazione con etidio bromuro.
Le sequenze dei primers utilizzati e le loro specificità
sono riportate nella Tabella 1.
32
Malattia delle mucose da BVDV tipo 2: descrizione di un focolaio in Puglia
Tabella 1
Primers utlizzati nei test RT-PCR e nested-PCR
a
b
Primer
Sequenza (5’→ 3’)
Senso
Specificità
Ampiezza amplificato
P1 a
P2 a,b
AACAAACATGGTTGGTGCAACTGGT
CTTACACAGACATATTTGCCTAGGTTCCA
+
-
BVDV-1, BVDV-2, BDV
826 bp
TS1 b
TS2 b
TS3 b
TATATTATTTGGAGACAGTGAATGTAGTAGCT
TGGTTAGGGAAGCAATTAGG
GGGGGTCACTTGTCGGAGG
+
+
+
BDV
BVDV-2
BVDV-1
566 bp
448 bp
223 bp
RT-PCR
nested-PCR.
Analisi di sequenza
Gli amplificati RT-PCR sono stati purificati su colonnine
Ultrafree-DA (Amicon, Millipore, Bedford, USA) ed inviati
alla Genome Express (Labo Grenoble, Francia), per il sequenziamento diretto con entrambi i primers. Le sequenze
ottenute sono state assemblate utilizzando il software
BIOEDIT (Hall, 1999) ed il consensus è stato confrontato
con le sequenze presenti nelle banche dati (EMBL/GenBank) mediante BLAST (NCBI, www.ncbi.nih.gov).
Esami sierologici
I sieri del vitellone morto e degli altri bovini presenti in
allevamento sono stati sottoposti al test di virusneutralizzazione (VN) per BVDV. La diluizione 1:5 di ciascun siero
da testare è stata messa a contatto per 45 min a 37°C con
100 TCID50/50 µl dello stipite BVDV tipo 2 omologo isolato dall’animale morto. Le miscele siero-virus sono state
inoculate in piastre microtiter a 96 pozzetti contenenti cellule di polmone embrionale bovino (PEB) e la lettura è
stata effettuata dopo 5 giorni di incubazione a 37°C.
Identificazione degli animali immunotolleranti e
persistentemente infetti (PI)
Allo scopo di identificare eventuali soggetti PI presenti in
allevamento, dai 4 bovini sieronegativi per BVDV sono stati
effettuati, a distanza di 30 giorni, due prelievi di sangue intero da sottoporre agli esami virologici. Da ciascun campione di sangue è stato raccolto il buffy coat mediante stratificazione su Lympholyte®-H (Cedarlane, Hornby, Ontario, Canada), seguendo le istruzioni della casa produttrice. I buffy
coats così ottenuti sono stati utilizzati in prove di isolamento
virale e nested-PCR per BVDV. Gli amplificati PCR sono
stati sequenziati e confrontati con i dati di sequenza dello
stipite BVDV-2 isolato dagli organi del vitellone morto.
Diagnosi differenziale
In sede di diagnosi differenziale sono state prese in considerazione alcune malattie infettive ad eziologia virale, in
grado di determinare quadri clinici anche solo in parte sovrapponibili alla sintomatologia osservata. In particolare,
sono stati effettuati test PCR per ovine herpesvirus-2
(OvHV-2), responsabile della febbre catarrale maligna
(Baxter et al., 1993), e per le infezioni sostenute da herpesvirus bovino tipo 1 (BoHV-1) (Vilcek, 1993) e tipo 4
(BoHV-4) (Boerner et al., 1999). Inoltre, è stato effettuato
un test RT-PCR per la ricerca dell’acido nucleico dei virus
della bluetongue (BTV) (Katz et al., 1993).
RISULTATI
Gli omogenati d’organo non hanno determinato la comparsa di effetto citopatico al primo passaggio su cellule, mentre, al secondo passaggio, è stato evidenziato un effetto citopatico tipico di BVDV, caratterizzato da vacuolizzazione del
citoplasma, aumento della rifrangenza e lisi cellulare. Il test
IFI per pestivirus ha fornito esito positivo, evidenziando una
caratteristica fluorescenza citoplasmatica (Fig. 5).
Il test RT-PCR per pestivirus ha prodotto un amplificato delle dimensioni attese (826 bp) in tutti i campioni esaminati (Fig. 6A). Con la nested-PCR sono stati ottenuti
amplificati di 448 bp, che hanno permesso di caratterizzare il virus isolato come BVDV tipo 2 (Fig. 6B).
L’analisi di sequenza mediante BLAST ha evidenziato
per lo stipite pestivirus isolato una elevata omologia con
sequenze BVDV tipo 2 presenti nella GenBank. In particolare è stata dimostrata una identità nucleotidica del
99% con lo stipite BVDV-2 Parker (numero di accesso
AF145971), isolato negli Stati Uniti, e del 98% con lo stipite BVDV-2 Giessen-1 (numero di accesso AF104030,
Becher et al., 1999), isolato in Germania.
Il test VN per BVDV ha evidenziato la presenza di anticorpi in 51/55 bovini dell’allevamento, con una sieroprevalenza pari al 93%, mentre il vitello morto, come atteso,
è risultato sieronegativo.
Gli esami virologici sui buffy coat dei 4 bovini sieronegativi presenti in allevamento hanno permesso di individuare
due animali con infezione persistente da BVDV-2: una
manza di 1 anno risultava positiva in RT-PCR e in n-PCR
ed un vitello di 6 mesi positivo solo in n-PCR. Da entrambi i soggetti è stato isolato uno stipite BVDV non citopatogeno caratterizzato come BVDV-2. A distanza di 30 giorni
tali risultati sono stati confermati.
L’analisi di sequenza del frammento amplificato ha dimostrato una perfetta identità nucleotidica rispetto allo
stipite BVDV-2 isolato dal vitellone.
I test PCR per BoHV-1, BoHV-4, OvHV-2 e BTV hanno fornito esito costantemente negativo.
33
DISCUSSIONE
FIGURA 5 - Test di immuonofluorescenza su PEB infette con lo stipite
BVDV isolato: fluorescenza citoplasmatica.
La diagnosi clinica di MD nei bovini non è particolarmente difficoltosa in quanto i sintomi, le lesioni e l’evoluzione sono tipici e, molto spesso, patognomonici.
Nel focolaio descritto nella presente nota è stato difficoltoso effettuare una diagnosi clinica in quanto, in base
ai sintomi e alle lesioni, si poteva anche sospettare una
infezione acuta grave da BVDV-2 trombocitopenico (SABVD). La mancanza di esami ematologici (conta piastrinica, ecc.) e i risultati preliminari di biologia molecolare
effettuati sugli organi dell’animale morto (stipite BVDV2) hanno reso ancora più complicata la formulazione di
una diagnosi precisa. Tuttavia l’isolamento dall’animale
morto di uno stipite BVDV-2 cp ha permesso di sospettare un focolaio di MD piuttosto che di SA-BVD, in
quanto in tutti i focolai finora descritti è stato sempre
isolato uno stipite BVDV-2 ncp (Corapi et al., 1990; Ridpath, 2003).
L’individuazione nell’allevamento di 2 soggetti PI, dai
quali è stato isolato uno stipite BVDV-2 ncp identico allo
stipite cp isolato dall’animale morto, ha permesso di
chiarire in modo definitivo che il focolaio osservato era
un grave episodio di MD.
BVDV-2 è stato più volte segnalato in Italia sia nei bovini (Giangaspero et al., 2001; Luzzago et al., 2001) che negli ovini (Pratelli et al., 2001). In particolare negli ovini è
stato isolato in allevamenti con gravi forme di clostridiosi
neonatali. Il grave episodio di MD descritto nella presente
nota pone in evidenza la necessità e l’urgenza di effettuare
piani di profilassi verso l’infezione da BVDV negli allevamenti bovini ed ovini prevedendo l’impiego di vaccini calibrati sulle nuove “realtà” epidemiologiche, allestiti cioè
con BVDV-1 e BVDV-2.
Figura 6A
Parole chiave
Bovino, pestivirus, malattia delle mucose.
Key words
Bovine, pestivirus, mucosal disease.
Bibliografia
1.
2.
3.
Figura 6B
FIGURA 6 - PCR e nested-PCR per pestivirus. (A) PCR panpestivirus
(826 bp): linea 1, marker GeneRulerTM 100bp DNA Ladder, MBI Fermentas GMBH, Germania; linea 2, controllo positivo BVDV-1; linea 3, controllo positivo BVDV-2; linee 4-6, organi del vitello morto (polmone, fegato, milza); linea 7, controllo negativo. (B) Tipizzazione mediante nested-PCR: linea 1, marker GeneRulerTM 100bp DNA Ladder, MBI Fermentas GMBH, Germania; linea 2, controllo positivo BVDV-1 (223 bp);
linea 3, controllo positivo BVDV-2 (448 bp); linee 4-6, organi del vitello
morto (polmone, fegato, milza); linea 7, controllo negativo.
4.
5.
6.
Baker JC. Bovine viral diarrhea virus: a review. J. Am. Vet. Med. Assoc. 1987, 190: 1449-1458.
Baker JC. The clinical manifestations of bovine viral diarrhea virus
infection. Vet. Clin. North Am. Food Anim. Pract. 1995, 11: 425445.
Baxter SIF, Pow A, Bridgen A, Reid HW. PCR detection of the sheepassociated agent of malignant catarrhal fever. Arch. Virol. 1993, 132:
145-159.
Becher P, Orlich M, König M, Thiel H-J. Nonhomologous RNA recombination in bovine viral diarrhea virus: molecular characterization of a
variety of subgenomic RNAs isolated during an outbreak of fatal mucosal disease. J. Virol. 1999, 73: 5646-5653.
Boerner B, Weigelt W, Buhk H-J, Castrucci G, Ludwig H. A sensitive
and specific PCR/Southern blot assay for detection of bovine herpesvirus 4 in calves infected experimentally. J. Virol. Methods 1999, 83:
169-180.
Brownlie J. The pathogenesis of bovine viral diarrhea virus infections.
Rev. Sci. Technical. 1990, 9: 43-59.
BOVINI
Large Animals Review, Anno 10, n. 2, Aprile 2004
34
7.
8.
9.
10.
11.
12.
13.
14.
15.
Malattia delle mucose da BVDV tipo 2: descrizione di un focolaio in Puglia
Buonavoglia C, Tempesta M, Marsilio F, Buonavoglia D, Gatti A, Sands
JJ, Compagnucci M. Border disease degli ovi-caprini. Nota sull’isolamento e la caratterizzazione del virus in Italia. O.&D.V. 1991, 12: 47-49.
Corapi WV, Elliott RD, French TW, Arthur DG, Bezek DM, Dubovi EJ.
Thrombocytopenia and hemorrhages in veal calves infected with bovine viral diarrhea virus. J. Am. Vet. Med. Assoc. 1990, 196: 590-596.
Falcone E, Tollis M, Conti G. Bovine viral diarrhea disease associated
with a contaminated vaccine. Vaccine 2000, 18: 387-388.
Giangaspero M, Harasawa R, Zecconi A, Luzzago C. Genotypic characteristic of bovine viral diarrhea virus 2 strains isolated in Northern
Italy. J. Vet. Med. Sci. 2001, 63: 1045-1049.
Hall TA. BioEdit: a user-friendly biological sequence alignment and
analysis program for Windows 95/98/NT. Nucl. Acids Symp. Ser.
1999, 41, 95-98.
Heinz FX, Collett MS, Purcell RH, Gould EA, Howard CR, Houghton M,
Moormann RJM, Rice CM, Thiel H-J. Family Flaviviridae. In: van Regenmortel MHV, Fauquet CM., Bishop DHL, Carstens EB, Estes MK,
Lemon SM, Maniloff J, Mayo MA, McGeoch DJ, Pringle CR, Wickner
RB (eds). Virus Taxonomy. Seventh Report of the International Committee on Taxonomy of Viruses. Academic Press, New York, 2000, pp.
859-878.
Katz JB, Gustafson GA, Alstad AD, Adler KA, Moser KM. Colorimetric
diagnosis of prolonged bluetongue viremia in sheep, using an enzyme-linked oligonucleotide sorbent assay of amplified viral nucleic
acids. Am. J. Vet. Res. 1993, 54: 2021-2026.
Luzzago C, Bandi C, Bronzo V, Ruffo G, Zecconi A. Distribution pattern
of bovine viral diarrhoea virus strains in intensive cattle herds in Italy.
Vet. Microbiol. 2001, 83: 265-274.
Meyers G, Thiel H-J. Molecular characterization of pestiviruses. Adv.
Virus Res. 1996, 47: 53-118.
16.
17.
18.
19.
20.
21.
22.
23.
24.
25.
26.
27.
Moenning V e Plagemann GW. The pestiviruses. Adv. Virus Res. 1992,
41: 53-91.
Moenning V, Liess B. Pathogenesis of intrauterine infections with bovine viral diarrhea virus. Vet. Clin. North Am. Food Anim. Pract. 1995,
11: 477-487.
Olafson P, MacCallum AD, Fox FH. An apparently new transmissible
disease of cattle. Cornell Vet. 1946, 36: 205-210.
Olafson P, Rickard CG. Further observations on the virus diarrhea
(new transmissible disease) of cattle. Cornell Vet. 1947, 37: 104-106.
Pellerin C, van den Hurk J, Lecomte J, Tussen P. Identification of a
new group of bovine viral diarrhea virus strains associated with
severe outbreaks and high mortalities. Virology 1994, 203: 260268.
Pratelli A, Martella V, Cirone F, Buonavoglia D, Elia G, Tempesta M,
Buonavoglia C. Genomic characterizion of pestiviruses isolated from
lambs and kids in Southern Italy. J. Virol. Methods 2001, 94: 81-85.
Rebhun WC, French TW, Perdrizet JA, Dubovi EJ, Dill SG, Karcher LF.
Thrombocytopenia associated with acute bovine virus diarrhea infection in cattle. J. Vet. Intern. Med. 1989, 3: 42-46.
Ridpath JF, Bolin SR, Dibovi EJ. Segregation of bovine viral diarrhea
virus into genotipes. Virology 1994, 205: 66-74.
Ridpath JF. BVDV genotypes and biotypes: practical implications for
diagnosis and control. Biologicals 2003, 31: 127-131.
Sullivan DG, Akkina RK. A nested polymerase chain reaction assay to
differentiate pestiviruses. Virus Res. 1995, 38: 231-239.
Thiel H-J, Plagemann PGW, Moenning V. Pestiviruses. In: Fields BN,
Knipe DM, Howley PM (eds). Fields Virology, 3rd edition., Vol 1. Lippincott-Raven, Philadelphia, 1996, pp. 1059-1073.
Vilcek S. Detection of the bovine herpesvirus-1 (BHV-1) genome by
PCR. J. Virol. Methods 1993, 41: 245-247.