l’occhio
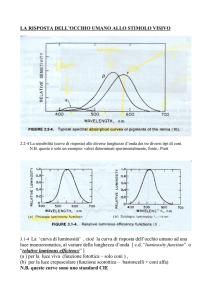
le proprietà della luce
la luce è il mezzo attraverso il quale il sistema visivo svolge la sua funzione
!
la luce è una radiazione magnetica visibile ai nostri occhi e viene descritta
utilizzando parametri quali lunghezza d’onda, frequenza e ampiezza d’onda.
!
una radiazione ad alta frequenza possiede molta energia come ad esempio i
raggi x
!
una radiazione a bassa frequenza possiede meno energia come le onde radio
l’occhio
le proprietà della luce
il nostro sistema visivo coglie solo una piccola parte dello spettro elettromagnetico
compresa tra i 400 e i 700 nanometri
!
un colore “caldo” come il rosso deriva da radiazioni ad onda lunga
!
un colore “freddo” come il blu deriva da radiazioni ad onda corta
!
la combinazione della gamma visibile emessa dal sole appare bianca
!
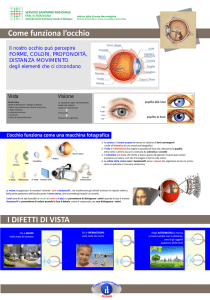
l’occhio
la struttura dell’occhio
!
la pupilla permette ai raggi di raggiungere la
retina, i suoi pigmenti neri assorbono luce e la
sua dimensione è controllata da un muscolo
chiamato iride i cui pigmenti sono il nostro
“colore” degli occhi. !
pupilla e iride sono rivestiti dalla cornea. !
questa struttura è priva di vasi e viene rifornita di
fluido dall’umor acqueo. la parte “bianca”
dell’occhio è la sclera alla quale sono collegati
tre muscoli extraoculari che permettono lo
spostamento dell’occhio.
l’occhio
la struttura dell’occhio
la retina è ricca di vasi sanguigni che si
originano dal disco ottico una regione
completamente priva di recettori e perciò
“cieca”.
!
al centro della retina troviamo una regione
più scura chiamata macula mentre il
centro esatto della retina è la fovea.
!
la parte di retina che va dalla fovea al naso è
detta nasale mentre la parte di retina
opposta è detta temporale
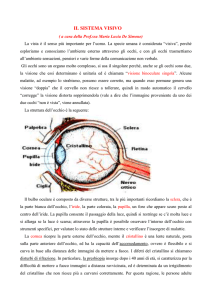
l’occhio
la struttura dell’occhio
andiamo ad analizzare l’occhio più da vicino:
!
sezionando l’occhio troviamo il
cristallino. localizzato dietro l’iride è
controllato da due muscoli ciliari.
!
gli aggiustamenti del cristallino ci
permettono di “mettere a fuoco”.
!
lo spazio tra cristallino e retina è riempito
dall’umor vitreo, una sostanza più densa
dell’umor acqueo.
l’occhio
la formazione dell’immagine nell’occhio
quando la luce colpisce l’occhio i suoi raggi
vengono deviati attraverso la rifrazione
convergendo verso la parte posteriore
dell’occhio
!
il punto in cui i raggi deviati convergono è
detto distanza focale e questa distanza
dipende dalla cornea; più stretta è la
curvatura minore è la distanza focale.
!
il reciproco in metri della distanza focale è
la diottria
l’occhio
la formazione dell’immagine nell’occhio
il cristallino contribuisce alla rifrazione della luce. il suo elevato potere di
messa a fuoco, specie con gli oggetti posti vicino all’occhio avviene modificando
la sua forma attraverso un processo chiamato accomodamento. quando la
luce colpisce l’occhio i suoi raggi vengono deviati attraverso la rifrazione
convergendo verso la parte posteriore dell’occhio
l’occhio
la formazione dell’immagine nell’occhio
anche la pupilla cambia in relazione alla luce. quando passiamo da un ambiente
scuro ad uno più illuminato le pupille si restringono. !
questo riflesso pupillare alla luce è dato dal collegamento tra retina e
neuroni del tronco. si tratta di un riflesso consensuale; stimolando un solo occhio
anche l’altro reagisce
l’occhio
la formazione dell’immagine
nell’occhio
quanto spazio visivo catturano i nostri
occhi? possiamo rispondere a questa
domanda analizzando il campo visivo.
!
se fissiamo un oggetto davanti ai nostri
occhi e mentre li teniamo fissi spostiamo
questo oggetto in diverse direzioni, ci
accorgiamo che a un certo punto
l’oggetto esce dal campo visivo.
l’occhio
la formazione dell’immagine nell’occhio
la capacità dell’occhio nel distinguere due punti
vicini è detta acuità ed è determinata dalla
spaziatura tra i recettori della retina e dalla
precisione della rifrazione
!
le distanze sulla retina vengono descritte in termini
di angolo visivo
l’occhio
!
!
nella tabella di Snellen i caratteri vengono distinti
uno dall’altro in base ai diversi angoli visivi.
!
la nostra vista è 20/20 se si riconoscono lettere che
sottendono un angolo di 0,083°
l’occhio
anatomia microscopica della retina
il sistema base per l’elaborazione dell’immagine
retinica prevede fotorecettori, cellule bipolari
e gangliari.
!
!
inoltre le cellule orizzontali ricevono i segnali dai
fotorecettori e comunicano con le cellule bipolari
mentre le cellule amacrine ricevono i segnali dalle
cellule bipolari e comunicano con le gangliari
!
IMPORTANTE!
✓SOLO i fotorecettori sono fotosensibili
✓SOLO le cellule gangliari lasciano la retina
l’occhio
anatomia microscopica della retina
la retina ha un’organizzazione laminare e le sue
cellule sono organizzate in strati.
!
i diversi strati sono denominati in questo modo:
!
1.strato di cellule gangliari (contiene le gangliari)
!
2.strato plessiforme interno (contiene le sinapsi
tra bipolari, amacrine e gangliari)
!
3.strato nucleare interno (contiene i corpi cellulari
di bipolari, amacrine e orizzontali)
l’occhio
4.strato plessiforme esterno (contiene le sinapsi
tra bipolari, orizzontali e fotorecettori)
5.strato nucleare esterno (contiene i corpi
cellulari dei fotorecettori)
6.strato dei segmenti esterni l’occhio
anatomia microscopica della retina
milioni di fotorecettori si occupano di tradurre
la luce in segnale nervoso
!
ogni fotorecettore può essere diviso in quattro
settori:
un segmento esterno, uno interno, un corpo
cellulare e una terminazione sinaptica.
!
a livello del segmento esterno sono presenti
molti dischi membranosi
!
i bastoncelli hanno un segmento esterno
grande e ricco di dischi mentre i coni hanno
un segmento esterno breve e povero di dischi
l’occhio
anatomia microscopica della retina
in condizioni di visione scotopica (bassa luminosità)
solo i bastoncelli contribuiscono alla visione in
quanto ricchi di fotopigmenti
!
al contrario in condizioni di visione fotopica (alta
luminosità) i coni svolgono un ruolo determinante
!
i bastoncelli possiedono un solo fotopigmento
mentre i coni esistono in tre varianti ognuna con un
proprio fotopigmento
!
l’occhio
anatomia microscopica della retina
la struttura della retina varia tra fovea e
periferia; !
la retina nelle porzioni periferiche presenta
bastoncelli sensibili a basse luminosità e quindi
questa porzione di retina è più sensibile alla
luce !
ogni cellula gangliare riceve input da diversi
fotorecettori
!
l’occhio
anatomia microscopica della retina
la visione diurna richiede l’intervento dei coni e un basso rapporto tra fotorecettori
e cellule gangliari in quanto l’immagine dev’essere dettagliata
!
la zona della retina in cui la visione è maggiormente dettagliata è la fovea
!
IMPORTANTE: in corrispondenza della fovea la retina si “infossa” permettendo alla
luce di colpire i fotorecettori senza attraversare gli altri strati cellulari
!
nella fovea NON ci sono bastoncelli ma SOLO coni
l’occhio
la fototrasduzione
per capire come avviene la conversione da luce
a potenziali d’azione seguiremo come esempio i
bastoncelli che sono molto più numerosi dei
coni nella nostra retina
!
la fototrasduzione ricorda molto da vicino
quanto succede in una comune trasmissione
sinaptica
!
la stimolazione luminosa attiva la
proteina G la quale attiva un enzima che va a
modificare la concentrazione citoplasmatica del
secondo messaggero.
!
tutto questo porta alla chiusura dei canali ionici
con conseguente modifica del potenziale di
membrana
!
l’occhio
la fototrasduzione
al buio il potenziale a riposo del segmento
esterno dei bastoncelli è -30mV (nelle
cellule nervose normalmente a -65mV) per via
del continuo ingresso di Na in appositi canali
!
questo movimento di Na è detto corrente
al buio
!
un particolare secondo messaggero chiamato
GMPciclico (guanosinmonofosfato ciclico)
viene continuamente prodotto dai
fotorecettori e stimola l’apertura dei canali
Na
!
capitoli
8-9-10-11-12-13-14-18-20-21-24
l’occhio
la fototrasduzione
in condizioni di buona luminosità, la luce riduce il
GMPc provocando la chiusura dei canali Na, il
potenziale si abbassa ulteriormente con
conseguente iperpolarizzazione dei bastoncelli
alla luce
!
l’occhio
la fototrasduzione
l’iperpolarizzazione è determinata
dall’assorbimento della luce da parte di un
fotopigmento dei bastoncelli chiamato
rodopsina.
!
la rodopsina è una proteina di membrana
composta da una parte proteica (l’opsina) e da
un agonista chimico (il retinale)
!
l’assorbimento della luce modifica la
conformazione del retinale che a sua volta attiva
l’opsina. questo processo è detto
“sbiancamento”
l’occhio
la fototrasduzione
lo sbiancamento stimola una proteina G chiamata
transducina ad attivare l’enzima fosfodiesterasi
(PDE) il quale riduce la concentrazione di GMPc e la
conseguente chiusura dei canali per Na dei bastoncelli
!
l’occhio
la fototrasduzione
ma cosa succede nei coni? nei coni la
fototrasduzione è simile a quella appena
descritta con una sola grande differenza data
dal diverso tipo di opsina presente nei
dischi del segmento esterno.
!
troviamo infatti tre opsine con sensibilità
sprettrali diverse:
coni “blu”
coni “verdi”
e coni “rossi”
!
l’occhio
la fototrasduzione
questi tre coni sono alla base della percezione
del colore. in ogni punto della retina esiste un gruppo di
tre tipi di coni (teoria tricromatica) per
cui in base alla risposta di questi tre coni
percepiamo i diversi colori.
!
oggi sappiamo che almeno in parte anche i
bastoncelli partecipano alla rilevazione del
colore, infatti siamo in grado di distinguere il
colore anche in condizioni di bassa luminosità.
!
l’occhio
la fototrasduzione
sappiamo bene che passare dalla luce al buio non è rapido. occorrono
diversi minuti per un buon adattamento al buio.
!
per adattarsi al buio è necessario dilatare le pupille, rigenerare rodopsina
oltre ad un generale riassestamento retinico. !
passare dal buio alla luce (specie se intensa) richiede molto meno tempo.
per un buon adattamento alla luce seguiamo un percorso opposto
rispetto a quello appena descritto.
!
l’occhio
l’elaborazione retinica
!
la verità è che lo stimolo preferito di un
fotorecettore è il buio, quindi in condizioni
di buio i fotorecettori liberano il loro
neurotrasmettitore; il glutammato.
!
ricordiamo che nello strato plessiforme esterno
le cellule bipolari e orizzontali entrano in
sinapsi con tutti i fotorecettori quindi in questo
strato l’input visivo viene raccolto e smistato
verso le cellule gangliari
!
Membrane potential (mV)
Light flash
Least intense flash response
−40
−45
−50
−55
−60
Most intense flash response
−65
0
100
200
300
Time (ms)
400
500
600
Figure 10.5
gle cone sti
(the cone h
which acco
of the respo
to a brief fla
highest ligh
rates (at abo
response is
tors; interes
tors depola
and Baylor,
nella retina
i fotorecettori NON presentano dei veri e propri PdA. la
much of the processing within the retina is mediated by graded potentials,
luce produce
dei cambiamenti
nel potenziale
membrana
e questo
largely because
action potentials
are not di
required
to transmit
information
influisce
sulla
nt liberato
over
the quantità
relativelydishort
distances involved.
Perhaps even more surprising is that shining light on a photoreceptor,
either a rod or a cone, leads to membrane hyperpolarization rather than depolarization (Figure 10.5). In the dark, the receptor is in a depolarized state,
Dark
e
Rod
Light
Rod
outer
segment
Ca
2+
Na
e
-
r
r
s
Rod
outer
segment
Na+
Ca2+ Na+
cGMP
cGMP
cGMP
cGMP
Inside
s
-
+
Outside
0
−
0
+
Rod
inner
segment
−
+
Rod
inner
segment
la visione si basa su un continuo aggiustamento tra le condizioni di luminosità e la
quantità di ntinliberato
the photopigment
rods and cones that contributes to the functional specialization
of these two receptor types. Most of what is known about the
!
molecular
events of phototransduction
hasconcentrazione
been gleaned from
la liberazione
di nt dipende dalla
di experiments
Ca.
in rods, in which the photopigment is rhodopsin (Figure 10.7A). When the
!
retinal moiety in the rhodopsin molecule absorbs a photon, its configuration
al buio
numero
di canali
aperti retinal;
per il Ca
alto mentre
si abbassa
changes
fromilthe
11-cis isomer
to all-trans
thisèchange
then triggers
a
di in
alta
seriescondizioni
of alterations
theluminosità
protein component of the molecule (Figure 10.7B).
The changes lead, in turn, to the activation of an intracellular messenger
called transducin, which activates a phosphodiesterase that hydrolyzes
drasticamente in
sodium channels in the outer segment
membrane.
(A)
(B)
Outer segment
membrane
C
3 PDE hydrolyzes cGMP,
reducing its concentration
Opsin
GMP
GMP
GMP
1 Light stimulation
of rhodopsin leads
to activation of a
G-protein, transducin
2 Activated G-protein
activates cGMP
phosphodiesterase
(PDE)
Light
α
11-cis retinal
GTP
Transducin
βγ
GDP
Na+
Na+
Closed Na+
channel
Disk
Rhodopsin
N
cGMP
cGMP
cGMP
Open Na+
channel
GTP
4 This leads
to closure of
Na+ channels
Disk
membrane
α
PDE
Inside of cell
Outside of cell
stimolazione luminosa - attivazione proteina G (transducina) attivazione fosfodiesterasi (PDE) - riduzione della concentrazione di
GMPc - chiusura canali Na.
rves10
5/14/04
10:45 AM
Page 242
242 Chapter Ten
−6
−4
−2
Starlight
Luminance (log cd/m−2)
2
0
Moonlight
4
Indoor lighting
6
8
Sunlight
Luminance of
white paper in:
Good color vision
Best acuity
No color vision
Poor acuity
Visual function
Scotopic
Absolute
threshold
Mesopic
Cone
threshold
Figure 10.9 The range of luminance
values over which the visual system
operates. At the lowest levels of illumination, only rods are activated. Cones
begin to contribute to perception at
about the level of starlight and are the
only receptors that function under relatively bright conditions.
Photopic
Rod
saturation
begins
50% bleach
Best
acuity
Indirect
ophthalmoscope
Damage
possible
elderly individuals suffering from macular degeneration (Box C). People
who have lost cone function are legally blind, whereas those who have lost
rod function only experience difficulty seeing at low levels of illumination
(night blindness; see Box B).
Differences in the transduction mechanisms utilized by the two receptor
types is a major factor in the ability of rods and cones to respond to different
ranges of light intensity. For example, rods produce a reliable response to a
single photon of light, whereas more than 100 photons are required to produce a comparable response in a cone. It is not, however, that cones fail to
effectively capture photons. Rather, the change in current produced by single
l’occhio
l’elaborazione retinica
comunemente classifichiamo le cellule bipolari in OFF e ON. !
bipolari off si depolarizzanno in risposta al glutammato !
bipolari on si iperpolarizzano in risposta al glutammato
bipolari off: depolarizzazione se la luce si spegne
bipolari on: depolarizzazione se la luce si accende
l’occhio
l’elaborazione retinica
!
ogni bipolare riceve afferenze da un numero variabile di fotorecettori oltre ad
essere connessa con le cellule orizzontali
!
questa rete di connessioni determina il campo recettivo della cellula
per campo recettivo si intende quell’area della retina che stimolata dalla luce
produce una modificazione del potenziale di membrana.
il campo recettivo di una bipolare si divide in due: un’area circolare che riceve afferenze direttamente dai fotorecettori chiamata
centro del campo recettivo mentre la parte adiacente che riceve
afferenze dalle cellule orizzontali è detta periferia del campo recettivo.
!
l’occhio
l’elaborazione retinica
la risposta di una bipolare alla luce verso
il suo centro è opposta alla risposta alla
luce diretta verso la sua periferia.
!
se la luce al centro depolarizza la cellula
(centro ON) la luce diretta alla periferia
la iperpolarizza (risposta OFF). per
questo si parla di campi recettivi
centro-periferia antagonisti.
!
l’occhio
le efferenze retiniche
la retina invia al cervello segnali che provengono unicamente dalle cellule gangliari
e la maggior parte di queste cellule possiede un campo recettivo centro-periferia
come quello appena descritto.
!
avremo quindi gangliari centroON e centroOFF in cui la risposta al centro è
contrastata dalla risposta alla periferia
!
of ganglion cells, “on”-center and “off”-center (Figure 10.14).
Turning on a spot of light in the receptive field center of an on-center ganglion cell produces a burst of action potentials. The same stimulus applied
to the receptive field center of an off-center ganglion cell reduces the rate of
(B)
(A)
(C)
Dark spot
in center
Light spot
in center
changes. (A) Effects of light spot in the
receptive field center. (B) Effects of dark
spot in the receptive field center. (C)
Effects of light spot in the center followed by the addition of light in the
surround.
Center plus
surround
Center
only
t3
t2
t2
t2
t1
t1
t1
t0
t0
t0
++
+++
++ + +
+++
++
On-center
ganglion cell
+++++
++++++++
+++++++++
++++
++++
+++
+++
+++
+++
+++
+++
++++
++++
+ + + + + + + ++
++++++
+++
Off-center
ganglion cell
t0
t1
t2
Time
t0
t1
t2
Time
t0
t1 t2
t3
Time
gangliari centro ON e OFF sono presenti in numero pressoché uguale. la
parziale sovrapposizione dei campi recettivi comporta che ciascun punto
sulla retina venga analizzato da diverse cellule gangliari on e off
!
le informazioni sull’aumento/riduzione della luce vengono portate
separatamente al cervello dalle gangliari on e off
!
in questo modo ogni piccola variazione nell’intensità luminosa viene
comunicata al cervello. Se questa informazione arrivasse lungo una sola via
sarebbe meno efficace in quanto un aumento (più luce) rispetto al livello
basso di frequenza di scarica verrebbe colto molto prontamente mentre una
riduzione (meno luce) verrebbe colta più lentamente.
dark spot in the center of the ganglion
cell receptive field.
intracellular cascade that closes cGMP-gated Na channels, reducing inward
(A)
(B)
Surround
Center
(C)
Light spot
in center
Surround
Dark spot
in center
t2
t2
t1
t1
t0
Center
cone
Glutamate
mGluR6
–
AMPA
kainate
+
t0
Center cone
Center cone
la depolarizzazione
graduale delle
cellule bipolari
aumenta il rilascio di glutammato e provoca la
depolarizzazione delle gangliari associate attraverso i
recettori
Cainato
e NMDA
t0 t1
t0 t1
t2
t2
!
Time
Time
Off-center
la differenzaOff-center
tracellle bipolariOn-center
on cell
e off è data
dai diversi
bipolar
bipolar
bipolar cell
recettori per il glutammato presenti sulla loro
membrana:
On-center
bipolar cell
On-center
bipolar cell
Off-center
bipolar cell
Glutamate
AMPA,
kainate,
NMDA
!
+
t1
t2
t1
t2
t1
t2
t1
t2
t2
t1
t2
t1
t2
t1
t2
ON - mGluR6
(iperpolarizzazione)
Off-center
On-center
Off-center
On-center
ganglion cell
ganglion
cell
ganglion
cell
ganglion
cell
OFF - AMPA e Cainato (depolarizzazione)
+
!
On-center
ganglion cell
Off-center
ganglion cell
t1
dark spot in the center of the ganglion
cell receptive field.
(A)
(B)
Surround
Center
(C)
Light spot
in center
Surround
t2
sinapsi tra fotorecettori e
bipolariOFF sono chiamate signconservative in quanto la
modificazione del potenziale della
membrana viene mantenuto
!
Glutamate
AMPA,
kainate,
NMDA
On-center
ganglion cell
t2
t1
t1
t0
t0
Center cone
Center cone
Center
cone
Glutamate
sinapsi tra fotorecettori e
AMPA
mGluR6
kainate
–
+
bipolariON sono chiamate
signinverting in quanto la
modificazioneOn-center
del potenziale dellaOff-center
bipolar cell
bipolar cell
membrana è opposta
!
!
!
!
!
!
Dark spot
in center
t0 t1
Time
t2
t0 t1
Time
t2
On-center
bipolar cell
Off-center
bipolar cell
On-center
bipolar cell
Off-center
bipolar cell
t1
t1
t1
t1
+
+
t2
t2
t2
t2
On-center
ganglion cell
Off-center
ganglion cell
On-center
ganglion cell
Off-center
ganglion cell
t1
t1
t1
t1
Off-center
ganglion cell
t2
t2
t2
t2
l’occhio
le efferenze retiniche
le gangliari rispondano a differenze di luminosità all’interno del loro campo recettivo
!
prendiamo una cellula centroOFF;
in caso di luminosità uniforme centro e periferia si annullano a vicenda (a)
!
quando l’ombra entra nel campo periferico la cellula si iperpolarizza (b)
!
al contrario quando l’ombra entra nel centro del campo la cellula si depolarizza (c)
!
infine quando l’ombra è uniforme centro e periferia tornano ad annullarsi (d)
!
l’occhio
le efferenze retiniche
quando consideriamo l’output di tutte le cellule centroOFF ognuna di esse
avrà uno dei quattro possibili output appena elencati
!
in questo modo possiamo concludere che questa organizzazione centroperiferia dei campi recettivi amplifica il contrasto tra i bordi. infatti il nostro
sistema visivo è tarato più sulla detezione di variazioni locali piuttosto che
sulla grandezza assoluta della luce
!
receptive field of an on-center cell toward its periphery, the response of the
cell to the spot of light decreases (Figure 10.16). When the spot falls completely outside the center (that is, in the surround), the response of the cell
falls below its resting level; the cell is effectively inhibited until the distance
from the center is so great that the spot no longer falls on the receptive field
at all, in which case the cell returns to its resting level of firing. Off-center
la stessa cosa riguarda una gangliare centro ON
Light
++
+++
++ ++
+++
++
++
+++
++ + +
+++
++
++
+++
++ + +
+++
++
++
+++
++ + +
+++
++
++
+++
++ + +
+++
++
Response rate (impulses/s)
100
80
60
40
20
Spontaneous level
of activity
0
1
2
3
4
Distance (degrees) from center of receptive field
5
Figure 10.16 Rat
on-center ganglion
as a function of the
from the receptive
the x axis correspo
distance of 5°, the
receptive field.
Dark
A
++
+++
++ + +
+++
++
Light
Edge
B
C
++
+++
++ + +
+++
++
D
++
+++
++ + +
+++
++
++
+++
++ + +
+++
++
On-center
ganglion cells
E
++
+++
++ + +
+++
++
D
Response rate
a hypothetganglion
A–E) are
k edge.
most
that lie
E
C
A
Spontaneous
level of activity
B
Position
cells exhibit a similar surround antagonism. Stimulation of the surround by
l’occhio
le efferenze retiniche
un esempio può chiarire quanto detto:
fissate la figura e rispondete se i due quadrati grigi al centro sono dello stesso grigio
la maggior parte di noi vede il grigio a sinistra come più scuro di quello a destra
nonostante siano perfettamente identici
l’occhio
vi sembrano ancora diversi?
ce
-
h
s
ve
en
xt
to
s
h
.
ir
d.
s
(B)
(C)
ce
-
h
s
ve
en
xt
to
s
h
.
ir
d.
s
(B)
(C)
l’occhio
i tipi di cellule gangliari
oltre alla distinzione centroON e OFF è possibile distinguere le gangliari anche
in base alla loro morfologia, fisiologia ed alle loro proprietà elettrofisiologiche.
!
le gangliari di grandi dimensioni sono dette di tipo M (circa il 5% delle cellule
gangliari) mentre quelle piccole sono dette di tipo P (circa il 90%).
il restante 5% di cellule sono classificate come nonM-nonP.
l’occhio
i tipi di cellule gangliari
le cellule M hanno campi recettivi più ampi e sono più sensibili a stimoli con
basso contrasto e inoltre rispondono ad uno stimolo al centro del campo con
una risposta transiente mentre le P rispondono con una scarica continua.
!
per tali ragioni si pensa che le M siano importanti per la detezione del
movimento mentre le P per l’analisi di dettagli fini
l’occhio
le cellule P sono sensibili a differenze nella lunghezza d’onda della luce, per questo
vengono dette anche cellule opponenti ai colori.
!
come funzionano queste cellule?
consideriamo una cellula P con centro ON per il rosso e periferia OFF per il verde.
la luce rossa verso il centro depolarizza la cellula mentre la luce verde diretta alla
periferia annulla la risposta del centro alla luce rossa
queste cellule vengono dette R+V- l’occhio
i tipi di cellule gangliari
!
l’opponenza tra blu e giallo (a carico delle cellule nonM-nonP) funziona allo
stesso modo: prendiamo una cellula centroON per il blu e periferiaOFF per il
giallo.
!
una luce blu al centro attiva i coni blu mentre una luce gialla in periferia attiva
sia quelli verdi che quelli rossi.
ng, Helmholtz, Maxwell,
, and Mach, to name only a
omas Young first proposed
must have three different
rticles”—i.e., the three cone
ental problem has been
h the relative activities of
pes can more or less explain
ceived in color-matching
performed in the laboratory,
n of color is strongly influtext. For example, a patch
exact same spectrum of
to the eye can appear quite
ending on its surround, a
called color contrast (Figure
, test patches returning difa to the eye can appear to be
r, an effect called color cone B). Although these phee well known in the niney, they were not accorded a
in color vision theory until
s work in the 1950s. In his
demonstration, Land (who
achievements founded the
pany and became a billionollage of colored papers that
erred to as “the Land Monuse of their similarity to the
Dutch artist Piet Mondrian.
lemetric photometer and
ble illuminators generating
, and long wavelength light,
d that two patches that in
modern debate about how color percepts
are generated that now spans several
decades. For Land, the answer lay in a
series of ratiometric equations that could
integrate the spectral returns of different
regions over the entire scene. It was rec-
LAND, E. (1986) Recent advances in Retinex
theory. Vis. Res. 26: 7–21.
PURVES, D. AND R. B. LOTTO (2003) Why We See
What We Do: An Empirical Theory of Vision,
Chapters 5 and 6. Sunderland MA: Sinauer
Associates, pp. 89–138.
(A)
(B)
The genesis of contrast and constancy effects by exactly the same context. The two panels
demonstrate the effects on apparent color when two similarly reflective target surfaces (A) or
two differently reflective target surfaces (B) are presented in the same context in which all the
information provided is consistent with illumination that differs only in intensity. The appearances of the relevant target surfaces in a neutral context are shown in the insets below. (From
Purves and Lotto, 2003)
l’occhio
i tipi di cellule gangliari
le cellule M non sono opponenti
al colore in quanto ricevono input
da tutti i diversi tipi di coni per
cui la percezione del colore è
data dall’attività relativa delle
cellule gangliari che ricevono
input da tutti e tre i tipi di coni.
!
fissate per circa 1 minuto la croce
al centro del quadrato rosso
!
poi passate a quello bianco vicino
l’occhio
i tipi di cellule gangliari
l’affaticamento dei coni rossi ci fa
vedere come verde il quadrato
bianco
l’occhio
i tipi di cellule gangliari
la stessa cosa accade fissando il
quadrato blu e passando poi a
quello bianco.
!
l’affaticamento dei coni blu ci farà
percepire il quadrato bianco
come se fosse giallo