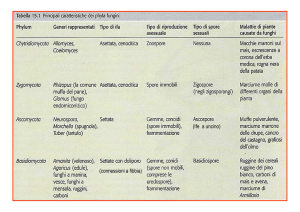
Organismi pionieri e rocce: le patine a ossalati di calcio
di
Marco Del Monte
Università di Bologna, Dipartimento di Scienze della Terra e Geologico-Ambientali
Via Zamboni 67, 40127, Bologna, Italy
E- mail: [email protected]
Gli organismi “pionieri” sono quelli che colonizzano per primi una roccia nuda in
campo: quindi batteri (che in realtà organismi non sono), alghe, funghi e soprattutto
licheni. Noi qui parleremo brevemente solo di questi ultimi rimandando per più
precise e dettagliate informazioni alla letteratura specializzata (Ahmadjian, 1993;
Brown, 1984; Carlile et al., 2001; Dalby et al.,1988; Deacon, 2000; Gilbert, 2000;
Hale, 1983; Hawksworth and Hill, 1984; Laundon, 1986; Nash, 1996; Ozenda et
Clauzade, 1978 ; si veda anche l’elenco dei siti web in calce alla bibliografia).
I licheni sono associazioni mutualistiche, vale a dire simbiosi, tra un fungo e un
partner capace di fotosintetizzare (un’alga o un cianobatterio) che danno vita a un
organismo composito. Il fungo forma un tessuto attorno alle cellule algali, fornendo
loro acqua e sostanze minerali assorbite tramite le ife dal substrato roccioso. A sua
volta il partner fotosintetico fornisce al fungo carbonio organico (amidi e zuccheri) e,
nel caso dei cianobatteri, azoto organico (nitrati e sali d’ammonio) a partire dall’N2
presente in atmosfera. L’alga fornisce inoltre alla simbiosi anche sostanze
auxotrofiche (vitamine). Le “informazioni” e lo scambio dei nutrienti tra i due
partner avvengono attraverso propagoli del fungo, che penetrano la parete algale,
detti austori.
Sotto il profilo nutrizionale dato che l’alga è un organismo autotrofo mentre il fungo
è eterotrofo, il più avantaggiato è il fungo. Sotto il profilo ecologico il fungo,
attraverso il fitto intreccio di ife (che formano il corpo o tallo lichenico), protegge
l’alga dall’essiccamento e dal congelamento risultando così fondamentale nella
colonizzazione di ambienti “estremi” (Alstrup, 1993; Alstrup and Cole, 1998;
Andreev et al., 1996; Hertel, 1987; Kershaw, 1985).
I licheni giocano così un ruolo fondamentale sia alla base della catena ecologica sia
come precursori della pedogenesi. Essi si comportano come pionieri
dell’insediamento vegetale. Colmate le microcavità o le microfratture con l’humus
prodotto dopo la morte dal lichene stesso essi rendono possibile lo sviluppo di
muschi e, concorrendo alla degradazione della roccia sottostante, quello di piante
erbacee prima e legnose poi.
Durante l’evoluzione i partner lichenici (micobionte e ficobionte) si sono adattati in
modo talmente stretto, che molte alghe dei licheni (ad esempio Trebouxia spp.)
probabilmente non hanno esistenza autonoma. Anche per quanto riguarda il tipi di
1
funghi, circa 14000 per il 96% ascomiceti, solo pochissimi ( < 1% ) sono stati
osservati in condizioni di vita libera (Deacon, 2000).
Il tallo di un lichene è, di solito costituito da una singola specie di fungo e da una
singola specie fotosintetizzante. Ospiti del lichene sono sempre batteri di uno o più
tipi e in diversi casi funghi parassiti (Faltynowicz, 1993; Goward and Thor, 1992).
Tra i funghi dominano, come appena detto, gli ascomiceti, molto meno comuni sono
i basidiomiceti ed infine eccezionali sono i licheni nei quali partecipano i
deuteromiceti. Sotto questo aspetto i licheni si possono dividere in Ascolicheni e
Basidiolicheni.
Il partner fotosintetizzante è costituito da un alga verde (Tribouxia già citata,
Clorella, Pleurococcus, Protocossus, Cystocossus) o da un cianobatterio ( o alga
azzurra del genere Nostoc, Gloeocapsa, Rivularia): si tratta di alghe monocellulari
anche se esistono rari licheni formati da alghe filamentose (ad es. Trentepholia).
Nella costituzione morfologica di un lichene la partecipazione del fungo è dominante
rispetto a quella dell’alga: quello che noi comunemente chiamiamo lichene (il tallo)
non è altro che un fitto intreccio di ife fungine. Secondo diversi A.A. i licheni
dovrebbero far parte del dominio dei Funghi sotto la denominazione di Funghi
lichenizzati.
I licheni, in base all’aspetto esteriore, vale a dire alla morfologia del tallo, possono
essere distinti in crostosi, fogliosi e cespugliosi. Parlando di rocce hanno interesse
praticamente solo i licheni crostosi: in questo caso il tallo si sviluppa sulla roccia
come crosta appiattita a forma spesso subcircolare. Inoltre occorre fare una
distinzione tra licheni epilitici e licheni endolitici: i primi, come dice il nome, vivono
all’esterno incrostando in superficie la roccia, i secondi vivono all’interno (sino a
qualche centimetro nelle pietre più trasparenti dove comunque deve essere garantita
la fotosintesi) della roccia colonizzata. Mentre i funghi liberi crescono molto
velocemente, il fungo del lichene epilitico, e quindi il tallo, sotto l’azione dell’alga si
accresce in modo estremamente lento (solo qualche mm/anno; Ozenda et Clauzade,
1978). Nel caso poi dei licheni endolitici la crescita è lentissima: essi sarebbero gli
organismi viventi del nostro Pianeta di età maggiore (Price, 1992).
Il tallo può essere coriaceo, carnoso o gelatinoso. Nel caso delle rocce è quasi
sempre coriaceo. Molto vario è il colore che va dal bianco al verde marcio, al verde
brillante, al giallo, al rosso ruggine, al rosso vivo, al violetto.
Se si osserva un tallo lichenico in sezione trasversale le cellule fotosintetizzanti
possono essere omogeneamente distribuite o essere disposte in un sottile strato. Nel
primo caso si parla di tallo omomero e nel secondo di tallo eterometro. In genere i
licheni incrostanti superficialmente le rocce presentano un tallo eterometro, mentre è
omomero quello dei licheni endolitici.
2
Per quanto concerne l’azione dei licheni nella degradazione meteorica, questa è una
conseguenza dell’ azione svolta dalle ife fungine che penetrano in profondità: esse
ancorano al substrato il lichene, forniscono i macroelementi e i microelementi della
nutrizione al simbionte estraendoli dai minerali della roccia sottostante alterandoli e
modificano le caratteristiche superficiali della roccia.
Per quanto riguarda la profondità massima di penetrazione delle ife, questa varia a
seconda del litotipo. Ad esempio in calcari e marmi essa raggiunge i 14 mm (Fig. 1;
Del Monte 1990; Del Monte e Ferrari, 1989).
Fig. 1 - Sezione trasversale di un lichene epilitico eterometro crostoso su calcare (Pietra d’Istria).
Le cellule algali si trovano in superficie, protette da un sottile cortex superiore. Le ife fungine
(diametro ≈ 2 µm) penetrano in profondità (max, osservata su questo tipo litologico 14 mm) alla
ricerca degli elementi della nutrizione canalizzando fortemente la pietra, alterandone così le
caratteristiche fisiche superficiali e favorendo la pedogenesi. La funzione di ancoraggio del tallo
alla superficie della roccia è importante ma subordinata a quella nutrizionale.
Nel caso di graniti la profondità è maggiore e può raggiungere i 25 mm (Del Monte,
1996). Le ife fungine penetrano attraverso la dissoluzione dei minerali (chelazione) e
forse anche attraverso la pressione determinata dal turgore cellulare. In ogni caso
l’apice dell’ifa ha dei sensori capaci di distinguere i minerali ricchi di elementi
chimici utili da quelli poveri o da quelli del tutto privi. Nel granito, ad esempio, il
quarzo (che non presenta nella sua struttura alcun elemento della nutrizione) viene
ignorato dalle ife che privilegiano innanzitutto la biotite (fig. 2) seguita dal
plagioclasio e infine dal K-feldspato (Del Monte et al., 1996).
3
Fig. 2 - Cristallo di biotite separata da un granito. Nei graniti le ife del micobionte penetrano a
profondità notevoli alla ricerca degli elementi della nut rizione (max. osservata 25 mm). Il minerale
più attaccato è la biotite che viene fortemente canalizzata. Le ife fungine penetrano sia lungo i piani
di sfaldatura (001), sia trasversalmente a questi ultimi. Seguono nell’ordine il plagioclasio e
l’ortoclasio. Il quarzo, al contrario, privo di elementi utili alla nutrizione, non è interessato
dall’attacco. Gli apici delle ife presentano evidentemente dei sensori chimici che permettono di
riconoscere gli elementi chimici e quindi i minerali utili. La perforazio ne avviene attraverso la
dissoluzione chimica (chelazione) e l’azione meccanica (pressione dovuta al turgore cellulare).
(Cattedrale di S. Giacomo di Compostela, Spagna).
Per quanto riguarda la riproduzione dei licheni essa può essere sia asessuata sia
sessuata. In numerosissime forme di licheni la riproduzione asessuata avviene
mediante i soredi che sono piccole formazioni globulari, ovvero licheni in miniatura,
in cui le ife fungine avvolgono alcune cellule dell’alga o del batterio specifico. I
soredi sono dispersi nell’ambiente dal vento o dall’acqua ruscellante.
Inoltre, per quanto riguarda il fungo, esiste anche la tipica riproduzione sessuata: i
corpi fruttiferi prendono il nome pericarpi (periteci o più comunemente apoteci) che
attraverso gli asci disperdono con forza nell’ambiente le spore fungine. Queste o
incontrano nel loro viaggio il partner fotosintetizzante specifico (ma ciò è poco
probabile) o, più comunemente, viaggiano portandosi dietro alcune cellule algali
dedicate.
Si è parlato subito sopra di organismi compositi: definire i licheni associazioni
mutualistiche o simbiosi è infatti riduttivo: l’unione porta a nuove e distinte unità
viventi.
In altri termini il lichene va ben oltre alla semplice somma delle caratteristiche e
delle funzione dei due partner il micobionte e il ficobionte. Questo “nuovo”
organismo assume pertanto proprietà morfologiche e fisiologiche non possedute dai
due simbionti quando essi siano isolati.
4
I licheni ad esempio hanno una grande versatilità ecologica che manca ai partner
isolati. Si possono sviluppare sui substrati più disparati: oltre alle rocce, i vetri
(Green and Snelgar, 1977), i laterizi (Del Monte, 1989), i metalli (Richardson, 1978)
e naturalmente i tronchi e i rami degli alberi.
Molti di essi mostrano resistenza a temperature estreme, resistenza all’essiccamento,
ecc.. Noto a tutti è il caso di Cladonia rangiferina (“lichene delle renne”) che
rappresenta nelle zone nordiche subpolari l’unico alimento di renne e caribù
(Thomson and Ahti, 1994; Zhurbenko, 1999).
D’altronde i licheni sono estremamente sensibili agli inquinanti aerodispersi e in
particolare ai composti dello Zolfo. Questa loro sensibilità che da una misura
integrata della qualità dell’aria in una data area nel tempo viene da tempo utilizzata
per il monitoraggio ambientale (Del Monte, 1989; Henderson, 1990; Richardson,
1975; Richardson, 1992).
Infatti le varie specie hanno limiti di tolleranza diversa: ad esempio Lecanora
conizaeoides o Lepraria incana sopportano sino a 150 µg/m3 di SO2 in aria,
Lecanora chlarotera sino a 50 µg/m3 e specie come Parmeliella plumbea o Pannaria
rubiginosa meno di 3 µg/m3
I “deserti lichenici” (fondamentalmente le grandi metropoli e le grandi aree
industrializzate come ad esempio la Valle Padana) sono (stati) prodotti
principalmente dagli alti valori di SO2 in atmosfera ( > 150 µg/m3 , negli anni ’90, in
inverno, in città). Sembra che quest’ultima eserciti un effetto deleterio sui cloroplasti
dell’alga portando il lichene ad una rapida morte (Richardson and Nieboer, 1983).
Per quanto riguarda il metabolismo i licheni producono un numero straordinario di
sostanze licheniche o prodotti lichenici o acidi lichenici (Cordoba, 1975; Culberson,
1969) che alga e fungo, quando crescono isolati, non sono assolutamente in grado di
sintetizzare.
Si danno questi nomi, che sono sinonimi, a tutti i composti organici, anaboliti e ai
cataboliti, che hanno a che fare coi licheni. Queste sostanze sono in massima parte
solide, hanno un punto di fusione di diverse decine di °C (i valori più comuni sono
compresi tra 125 e 260 °C) e in genere, ad eccezione dell’acido ossalico, sono da
scarsamente solubili a insolubili in acqua.
Esse ammontano a diverse centinaia e farne un elenco anche parziale sarebbe del
tutto privo di significato. Si rimanda perciò alla letteratura specializzata (Cordoba,
1975).
Le sostanze che comunque, per la (modestissima) esperienza dello scrivente,
compaiono più spesso sono ad esempio gli acidi: ossalico, lecanorico, lobarico,
usnico, pulvinico, olivetorico, evernico, imbricarico, umbilicarico, lobarico,
barbatico, tamnolico, salacinico, galico e molti altri.
Un ruolo del tutto particolare è giocato dall’acido ossalico H2C2O4 (Ethanedioic
acid): quest’acido che sublima a 157 °C e si decompone a 189.5 °C e viceversa
solubile in H2O ( sol.: 3; Lider, 2000). Esso è prodotto in notevole quantità dal
micobionte, ma talora anche dal ficobionte, in particolare dai cianobatteri (Del Monte
5
and Sabbioni, 1983). Si suppone che l’eccesso in Ca venga eliminato dal lichene
attraverso la precipitazione di due ossalati di calcio: la Weddellite (CaC 2O4 . 2H2O,
Tetragonale bipiramidale) e la Whewellite (CaC 2O4 . H2O, Monoclina prismatica).
Questi due minerali (ovvero o l’uno o l’altro) si trovano spesso in piccoli cristalli da
idiomorfi a subidiomorfi all’interno del tallo lichenico.
La Whewellite è statisticamente più diffusa della Weddellite e sembra perciò essere il
minerale più stabile in natura (Del Monte and Sabbioni, 1987).
L’acido ossalico può anche disciogliersi nell’acqua di pioggia, e seguirne la sorte
durante il suo ruscellamento verso il basso, sulla superficie delle rocce. In questo
caso, nei sottosquadri, nelle rientranze, nelle nicchie e più in generale in tutte le zone
a valle bagnate ma non soggette a forte wash-out la soluzione acida può interagire
con la calcite formando sottilissimi veli bruni di ossalato (o ossalati) di calcio (Fig.
3). Queste patine brune (in realtà il colore va dal nocciola, al bruno rosa, al bruno
scuro sino al rosso ruggine) sono estremamente diffuse in natura ma abbastanza
difficili da evidenziare (Del Monte et al., 1987). Il motivo consiste nel fatto che di
solito il loro spessore è dell’ordine della decina di µm e solo eccezionalmente
raggiunge il centinaio di µm. Ne consegue che se si raschia la patina senza le dovute
cautele ( occorre cioè asportarla delicatamente da una superfici abbastanza estesa) si
preleva soprattutto la calcite del substrato: sarà questo alla fine l’unico minerale
messo in evidenza mediante XRD.
Fig. 3 – I licheni crostosi colonizzano tutte le superfici delle rocce più esposte alla circolazione
d’aria, all’acqua di pioggia e alla radiazione solare. L’acqua delle idrometeore raggiungendo e
impregnando il tallo dei licheni si acidifica per acido ossalico, prodotto da questi organismi (in
ragione di alcune centinaia di ppb/g). La soluzione acida così formata, scorrendo verso il basso può
reagire con la calcite del calcare o del marmo: l’acido ossalico, essendo l’ossalato di calcio, meno
solubile della calcite, sposta l’acido carbonico portando alla precipitazione della Weddellite e/o
della Whewellite. Col passare del tempo si formano così sottili veli di Ca-ossalati (10-100 µm
circa) di colore nocciola, bruno rosa, bruno scuro o rosso ruggine. Il colore è dato da corpuscoli
organici imprigionati nel precipitato, come spore fungine, frammenti di ife, colonie batteriche,
ecc..Queste patine sono caratteristiche delle nicchie, dei sottosquadri e delle rientranze in genere in
quanto qui i licheni sono assenti e l’acqua di pioggia acidificata, bagnando la pietra senza
6
ruscellare, può interagire con essa per un tempo sufficientemente lungo a indurre la reazione
chimica Ca++ + 2HCO3 - + C2O4-- ⇒ CaC2O4 .H2 O + CO2 ⇑.
I licheni colonizzano sia le pietre degli affioramenti naturali, sia ovviamente anche
quelle messe in opera nella realizzazione di manufatti (miliari, muretti, mura di cinta,
fabbricati, ecc.) o monumenti (chiese, palazzi, bassorilievi, altorilievi, rilievi a tutto
tondo, ecc.). In queste opere dominano sia le superfici verticali sia i sottosquadri, le
sporgenze e le rientranze, le nicchie ecc.., vale a dire geometrie simili a quelle
schematizzate in figura 3. Per questo le patine a ossalati di calcio sono così comuni
sulle superfici dei monumenti (Fig. 4).
Fig. 4 - Pinnacolo in campo aperto in calcare. La superficie della pietra è quasi totalmente ricoperta
da licheni crostosi epilitici (grigio-neri nella foto). Le due nicchie, non colonizzate dai licheni
(bianche nella foto), sono invece ricoperte da una sottile patina bruno-rosata costituita da ossalati di
calcio. (Nostra Signora della Battaglia, Portogallo, particolare)
I licheni che, come abbiamo detto, non tollerando i composti dello zolfo
aereodispersi hanno da tempo abbandonato le città (Bates and Farmer, 1992.
Del Monte, 1991). Gli ossalati di calcio, così comuni sulle superfici dei monumenti
urbani rappresentano la memoria dei tempi passati quando la qualità dell’aria era
buona, i licheni vivevano felici nelle città, il biodegrado era uno dei principali fattori
di danno e la lotta alle “male erbe” uno dei principali interventi conservativi che si
rendeva necessario per la salvaguardia delle opere d’arte (Del Monte and Sabbioni,
1987; Del Monte and Sabbioni, 1988).
Bibliografia
Ahmadjian V. The Lichen Symbiosis. Wiley, Chichester, 1993.
7
Alstrup V.. News on lichenicolous fungi from the Nordic countries. Graphis
Scripta, 5, 96-104, 1993.
Alstrup V.and M.S. Cole. Lichenicolous fungi of British Columbia. Bryologist,
101, 221-229, 1998.
Andreev M., Y. Kotlov and I. Makarova. Cheklist of lichens and lichenicolous
fungi of the Russian Arctic. Bryologist, 99, 137-169, 1996.
Bates J.W. and A.M. Farmer. Bryophytes and Lichens in a Changing
Environment. University Press, Oxford, 1992.
Brown D. H. (ed.). Lichen physiology and cell biology. Plenum Press, New York,
1984
Carlile M.J., S.C. Watkinson and G.W. Gooday. The fungi. Academic Press,
London, 2001
Cordoba C. V.. Fisiologia de las sustancias liquénicas. Editorial Alhambra, Madrid,
1975.
Culberson C. F.. Chemical and Botanical Guide to Lichen Products. The
University of Nord Carolina Press, Chapel Hill, 1969.
Dalby D. H., D. L. Hawksworth and S. L. Jury (eds). Horizons in Lichenology.
Academic Press, London, 1988.
Deacon J.W.. Micologia Moderna. Calderini Ed agricole, Bologna, 2000.
Del Monte M.. I monumenti in pietra e i licheni. Rass. Beni Culturali, V, 3, 12-17 ,
1989.
Del Monte M.. Microbioerosions and biodeposits on stone monuments: pitting
and calcium oxalate patinas. Advanced Workshop on "Analytical Methodologies
for the Investigation of Damaged Stones", 27 pp, Pavia, 1990.
Del Monte M.. Trajan's Column: Lichens don't live here anymore. Endeavour,
15, 2, 86-93, 1991.
Del Monte M.. Experimental weathering test on granitic rocks. In Conservation
of granitic rocks with application to the megalithic monuments. CE Research
report n. 5 (J. Delgado-Rodrigues ed.), 192-193, 1996.
8
Del Monte M. e A. Ferrari. Patine da biointerazione alla luce delle superfici dei
monumenti. Atti Conv. Int. " Le pellicole ad ossalati : origine e significato nella
conservazione delle opere d'arte", 171-182, Milano, 1989.
Del Monte M. and C. Sabbioni. Weddellite on limestone in the Venice
Environment. Environmental Science & Technology, 17, 518-522, 1983.
Del Monte M. and C. Sabbioni. The so-called "scialbatura" on Roman Imperial
marbles. Studies in Conservation, 32, 114-121, 1987.
Del Monte M. and C. Sabbioni. Superficial Patinas on Ancient Monuments.
Europ. Cult. Herit. N.L.R., 2, 4, 11-13, 1988.
Del Monte M., Ratazzi A., Romao P. and P. Rossi. The role of lichens in the
weathering of granitic buildings. In : " Degradation and conservation of granitic
rocks in monuments", Proceedings of EC Workshop held in Santiago de Compostela,
(J. Delgado-Rodrigues ed.), 301-306, 1996.
Del Monte M., C. Sabbioni and G. Zappia. The origin of calcium oxalates on
monuments, historical buildings and natural outcrops. The Science of the Total
Environment, 67, 17-39, 1987.
Faltynowicz W. A checklist of Polish lichen forming and lichenicolous fungi
including parasitic and saprophytic fungi occurring on lichens. Polish Bot. Stud.,
6, 1-65, 1993.
Gilbert O.. Lichens. Harper Collins, London, 2000.
Goward T. and G. Thor. Notes on lichens and allied fungi of British Columbia.
Bryologist. 95, 33-37, 1992.
Green T.G.A. and W.P. Snelgar. Parmelia scabrosa on glass in New Zeland.
Lichenologist, 9, 170-171, 1977.
Hale M.E.. The Biology of Lichens (3rd edn) , Arnold, London, 1983.
Hawksworth D.L.and D.J. Hill. The Lichens forming Fungi. Blackie, Glagow,
1984.
Henderson A.. Litterature on air pollution and lichens XXXI. Lichenologist, 22,
173-182, 1990.
Hertel H.. Progress and problems in taxonomy of Antartic saxicolous lichens.
Bibliotheca lichenologica, 25, 219-242, 1987.
9
Kershaw K.. Physiological ecology of lichens. Cambridge University Press, 1985.
Laundon J.R.. Lichens. Shire Publication, Aylesbury, 1986.
Lawrey J.W.. The Biology of Lichenized Fungi. Praeger, New York, 1984.
Lide D.R. ed.-in-Chief. CRC Handbook of Chemistry and Physics, CRC Press,
Boca Raton, 2000.
Nash T.H.. Lichen Biology. Cambridge University Press, Cambridge, 1996.
Ozenda P. et G. Clauzade. Les lichens, étude biologique et flore illustrée. Dounot,
Paris, 1978.
Price R.C.. The Methuselah Factor: age in cryptoendolithic communities. Trends
in Ecology and Evolution, 7, 21-24, 1992.
Richardson D.H.S.. The Vanishing Lichens. David & Charles, Newton Abbot,
1975.
Richardson D.H.S.. Pollution monitoring with Lichens. The Richmond Publishing
Company, Slough, 1992.
Richardson D.H.S..Lichens on iron cannon balls. Lichenologist, 10, 233-234, 1978.
Richardson D.H.S.. War in the world of lichens: parasitism and symbiosis as
exemplified by lichens and lichenicolous fungy. Mycological Research, 103, 641650, 1999.
Richardson D.H.S. and E. Nieboer. Ecophysiological response of lichens to
sulphur dioxide. Journal of the Hattori Botanical Laboratory, 54, 331-351, 1983.
Seaward M.R.D. Contributions of lichens to ecosystems. In M. Galun ed., CRS
Handbook of Lichenology, 2, pp. 107-129, CRC Press, Boca Raton, 1988.
Thomson J.W. and T. Ahti. Lichens collected on an Alaska High-way Expedition
in Alaska and Canada. Bryologist, 97, 139-157, 1994.
Zhurbenko M. Lichens and lichenicolous fungi from the Putorama plateau,
Siberian subartic. Folia Crypt. Estonica, 34, 71-79, 1999.
10
http://lichen.com. The North American Lichen Project.
http://mgd.nacse.org/hyperSQL/lichenland/
http://www.nacse.org/lichenair/
http://www.botany.hawaii.edu/cpsu/lichen1.html
http://www.ucmp.berkeley.edu/fungi/lichens/lichens.html
http://www.ndsu.nodak.edu/instruct/esslinge/chcklst/chcklst7.htm
11