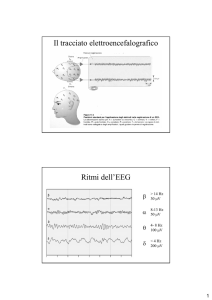
Fisiologia del sonno
• Pavlov (1915): il sonno come risultato di un
condizionamento
• Von Economo (1926): centro della veglia e centro del
sonno (encefalite letargica) . Ruolo dell’ipotalamo
• Nauta (1946) centro del sonno e centro della veglia
(conferma sperimentale)
• Hess (1954) il sonno fenomeno attivo
sperimentale)
(conferma
• Pavlov (1954): il sonno fenomeno passivo ed attivo
Fisiologia del sonno (teorie
“passive” del sonno)
• Kleitman (1929): (Teoria) Il sonno come
fenomeno passivo dovuto ad una riduzione
delle afferenze sensoriali che sono in grado di
mantenere le condizioni di veglia
• Bremer (1935) conferma sperimentale
all’ipotesi di Kleitman: teoria della
deafferentazione (praparati cerveau isolé ed
enecéphale isolé)
Fisiologia del sonno (teorie
“passive” del sonno)
• Bremer (1935) praparato cerveau isolé:
sezione mesencefalica completa fra i
collicoli superiori e inferiori => attività
EEG tipica del sonno (onde lente);
enecéphale isolé: sezione più caudale
tra la parte caudale del bulbo e il
midollo => alternanza S-V.
Fisiologia del sonno (teorie “passive”
del sonno)
• Moruzzi e Magoun (1949): identificano una struttura
diffusa in tutto il tronco dell’encefalo; la Formazione
Reticolare Ascendente, la cui stimolazione in un
reparato enecephale isolé produce un quadro EEG di
veglia attiva
• Lindsey e coll (1950): la lesione della FR,
mantenendo intatte le afferenze sensoriali verso i
centri superiori, produce sonno perenne =>teoria
reticolare :il sonno è un fenomeno passivo risultato
della caduta di tono corticale a seguito della ridotta
attività della FR
• In realtà nel tronco cerebrale ci sono
regioni che favoriscono od ostacolano il
sonno. Ipotesi confermata
dall’anestesia selettiva di parti diverse
del tronco cerebrale.
Teorie “attive” del sonno: Batini et al., 1959
• strutture caudali
bulbopontine
possono indurre
sincronizzazione
EEG e sonno
comportamentale
• strutture
rostropontine e
mesencefaliche
sono responsabili dello
stato di veglia
• “nuclei del fascio
solitario”, se
stimolati, inducono
attivamente la
sincronizzazione
dell’attività EEG
riducendo l’attivazione
della FR ascendente
Componente talamica
Componente talamica
• Esperienze di stimolazione/lesione chimica
ed elettrofisiologiche del nucleo reticolare
del talamo (NRT) nell’animale hanno
messo in evidenza la sua funzione di
pacemaker dell’attività sincrona del talamo
(Steriade e coll 1993).
Componente talamica
• Il passaggio dalla desincronizzazione della
veglia alla sincronizzazione del sonno è
caratterizzato da una variazione di attività
dei neuroni talamo-corticali che da una
scarica continua passano ad una scarica a
raffica (burst)
• Tali oscillazioni sinaptiche sono il risultato di
una interazione in un circuito complesso in cui
gioca un ruolo fondamentale l’attività
inibitoria del GABAergica dei neuroni del
NRT.
Sintesi
• Non esistono centri specializzati per il sonno e
per la veglia, ma la partecipazione coordinata
di numerose componenti:
– Prosencefalica
– Talamica
– Ipotalamica
– Troncoencefalica
Il passaggio dalla veglia al
sonno
(addormentamento)
potrebbe essere dovuto
all’attivazione di neuroni
(GABA?) delle regioni
ipotalamiche anteriori e
prosencefaliche che iniziano
le sequenze sinaptiche
sincronizzanti sia a livello
del talamo che della
corteccia, e parallelamente
coinvolgono neuroni
sincronizzanti caudali del
tronco che entrano in fase,
facilitandole, con le
sequenze talamiche.
Il passaggio dal sonno
sincrono alla veglia
(risveglio) o al sonno
REM può essere visto
come un attivazione dei
neuroni reticolari pontinomesencefalici e ipotalamici
posteriori che rompono le
sequenze talamiche sia
agendo direttamente sui
neuroni talamo-corticali che
inibendo i neuroni del NRT
dove operano neuroni
GABAergici ( e in parte
anche colinergici).
Fisiologia del sonno REM
• Sonno caratterizzato da
desincronizzazione EEG, caduta di
attività dei muscoli antigravitari,
comparsa di movimenti oculari rapidi,
irregolarità del respiro e dell’attività
cardiaca
Fisiologia del sonno REM
• Caratterizzato da componenti toniche
e componenti fasiche
• Fenomeni tonici: che si mantengono per
tutta la durata dell’episodio
– Desincronizzazione EEG
– Atonia posturale
• Fenomeni fasici: che compaiono
saltuariamente durante l’episodio
– Onde PGO (ponto-genicolooccipitali)
– REMs (movimenti oculari rapidi)
Fisiologia del sonno REM
• Gli studi di Jouvet hanno messo in evidenza nel
ponte le strutture responsabili dello
scatenamento e del mantenimento dei fenomeni
tonici e fasici del sonno REM:
Fisiologia del sonno REM
• Localizzazione di gruppi di neuroni in area
dorsolaterale del ponte comprendenti il locus
coeruleus implicati nell’atonia posturale del
sonno REM (Jouvet e Delorme 1965).
• La lesione di queste aree abolisce l’inibizione
motoria: il gatto in sonno REM mostra un
comportamento esplorativo o di attacco-fuga,
come se agisse un vissuto di tipo onirosimile.
Fisiologia del sonno REM
• Ulteriori studi hanno suggerito
l’esistenza nel ponte di neuroni (PS-on,
o REM-on) deputati alla genesi del
sonno paradosso, sottoposti ad un
controllo inibitorio da parte di neuroni
bulbari (PS-off, o REM-off). La
deattivazione di questi ultimi scatena
infatti un episodio di sonno REM.