Evoluzione del concetto di gene II
La complementazione
Un gene = un polipeptide
Definizione scarsamente operativa perché in un organismo
complesso non è possibile corrrelare ogni segmento di DNA
con uno specifico polipeptide tra i tanti prodotti.
Meglio utilizzare un approccio
genetico piuttosto che biochimico
Test di complementazione introdotto da Ed Lewis
e rielanorato da S. Benzer per definire
sperimentalmente il concetto di gene nel fago T4
Per verificare se due mutazioni sono a
carico dello stesso gene (e quindi sono
alleliche) o se sono a carico di geni diversi
si usa il test di complementazione o test
cis-trans o test di allelismo
Mappatura del genoma fagico
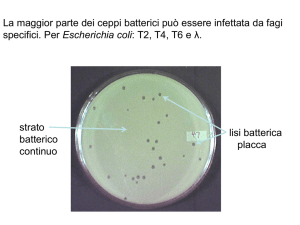
Fenotipi fagici
placche di lisi e specificità d’ospite (ceppo
batterico che un fago è in grado di lisare)
Marcatori del fago T2:
h+ lisa il ceppo B di coli ma non il ceppoB/2
h
lisa sia il ceppo B che il B/2
r+ placche piccole con margini indistinti
r placche grandi con margini netti
quando fagi h+ crescono su uno strato
misto di cellule B e B/2 formano
placche torbide perchè lisano solo B
e i batteri B2 crescono nelle placche
provocando torbidità delle placche. I
fagi h formano invece placche chiare
ceppo B
Il lisato si piastra su una
miscela di ceppi B e B/2
Frequenza di
ricombinazione
placche (h+ r+) + (h r )
tra h ed r
=
placche totali
x 100
Come di effettua il test di
complementazione nei fagi?
Esempio: test di complementazione tra mutazioni del gene rII
del fago T4
I fagi mutanti rII producono placche grandi con margini netti ; sono in
grado di lisare il ceppo B di E.coli ma non il ceppo K(λ).
I fagi selvatici rII+ producono placche piccole con margini irregolari;
sono in grado di lisare sia il ceppo B di E.coli che il ceppo K(λ).
Per fare il test di complementazione tra due fagi mutanti
rII (es. rII1 e rII2) si fa una doppia infezione (o infezione
mista) su K(λ) e si verifica se avviene la lisi oppure no.
I fagi dei due ceppi mutanti vengono piastrati ad alta molteplicità di
infezione cioè ad un concentrazione elevata in modo che i batteri
vengono infettati contemporamenamente da entrambi i tipi di mutanti
Se si ha la lisi delle cellule K(λ) vuol dire che le mutazioni
sono a carico di geni diversi e pertanto complementano. Se
non si osserva lisi le mutazioni sono a carico dello stesso
gene e di conseguenza non complementano
Un gene o cistrone è la regione
genetica all’interno della quale non
c’è complementazione tra
mutazioni
Il cistrone è l’unità di funzione
Il cistrone prende il nome dal test di complementazione
che si chiama test cis-trans
trans
m1
+
cis
+
m2
complementazione
trans
m1
m2
m1
m2
+
+
complementazione
cis
+
+
assenza di
complementazione
+
+
+
complementazione
m1
m2
Test di complementazione in
Drosophila
Nei batteri il test si
effettua costruendo
diploidi parziali stabili
mediante sesduzione
Mediante il test di complementazione è
possibile definire il gene come unità di
funzione
La struttura fine del gene
Benzer (negli anni 50’) utilizzò un sistema molto sensibile per
mettere in evidenza la ricombinazione intragenica (il fago T4)
e costruì una mappa genetica dettagliata di siti all’interno del
gene rII
La sensibilità del sistema era data dal fatto che i fagi
producono progenie molto numerosa pertanto era possibile
mettere in evidenza eventi di ricombinazione rari come quelli
che si potrebbero verificare all’interno di un gene
Che differenza c’è tra complementazione e
ricombinazione?
Nella complementazione i
genotipi della progenie fagica
rimangono mutanti come quelli
dei genitori. Avviene solo un
mescolamento di prodotti genici
o
Nella ricombinazione i genotipi della progenie
sono diversi da quelli dei genitori
rIIA
rIIA
rIIB
rIIB
Analisi genetica della progenie fagica
La progenie fagica si piastra su
E.coli B
Complementazione
Ricombinazione
E.coli K(λ)
tutti i fagi
formano placche
(titolo elevato)
non si osserva
formazione di
placche
tutti i fagi
formano placche
(titolo molto
basso)
formano placche
la metà dei fagi
(i selvatici) che
lisano B. I doppi
mutanti non
formano placche
Benzer isolò circa
3000 mutanti rII e
li saggiò a coppie
per vedere se erano
allelici o no
rII
rII+
K(λ)
assenza di
lisi
placche
piccole
margini
irregolari
B
placche
grandi
margini netti
placche
piccole
margini
irregolari
Benzer fece delle doppie infezioni nel ceppo di E. coli K(λ)
Dai dati di complementazione capì che le mutazioni rII mappavano in due
unità di funzione il cistrone A ed il cistrone B
I mutanti del cistrone A non complementavano tra loro ma
complementavano con tutti i mutanti del cistrone B. I mutanti del cistrone
B non complementavano tra loro ma complementavano con tutti i mutanti
del cistrone A.
cistrone A
cistrone B
rII
Il numero dei mutanti assegnati a
ciascun cistrone risultò più o meno
lo stesso (circa 1500)
Benzer fece doppie infezioni con coppie di mutanti rII (tutti
appartenenti allo stesso cistrone) sul ceppo B ed il lisato
fagico ottenuto lo piastrò sul ceppo K(λ) (sul quale potevano
crescere solo i ricombinanti selvatici) ed ottenne ricombinanti
selvatici che si erano originati da ricombinazione intragenica.
I dati di Benzer confermarono che la ricombinazione può
avvenire anche tra siti all’interno di uno stesso gene.
(Questi siti furono chiamati reconi)
frequenza di
ricombinazione tra
due alleli
=
numero di placche
presenti sul ceppo B
ricombinanti rII+
x
2
numero totale della progenie
Benzer non osservò mai frequenze di ricombinazione
inferiori allo 0,01% anche se il suo sistema era in grado di
mettere in evidenza anche frequenze di ricombinazione
dello 0,0001% . Ciò suggeriva l’esistenza di un limite
fisico al di sotto del quale non può avvenire
ricombinazione.
Successivamente si capì che il limite fisico definito dalla
la più piccola unità di ricombinazione (il recone)
coincideva con la singola coppia nucleotidica