caricato da
annax86
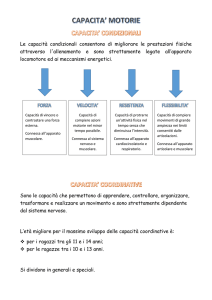
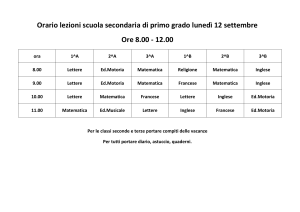
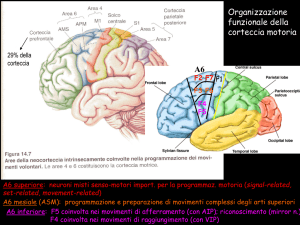
IL CONTROLLO MOTORIO

CAPITOLO 1 I MUSCOLI I muscoli sono gli organi più sviluppati del corpo, tanto che rappresentano la metà del nostro peso corporeo. Essi agiscono sia nel lavoro fisico dell’organismo che in quello intellettuale: infatti in azioni come lo scrivere, parlare o leggere viene impiegato l’apparato muscolare. I muscoli permettono il movimento agendo sulle articolazioni scheletriche e modificando la loro posizione nello spazio. I muscoli sono legati alle ossa tramite i tendini o fasci di tessuto connettivo e in corrispondenza delle articolazioni essi operano secondo azioni opposte: infatti nel momento in cui un muscolo si contrae (ossia si accorcia) un altro muscolo si estende (ossia si allunga) e questa loro relazione viene definita “antagonistica”. La conseguenza di tutto ciò è che nel momento in cui si realizza un’azione coordinata un gruppo di motoneuroni viene eccitato, mentre un altro viene inibito. Tale rapporto è determinato dal fatto che i muscoli esercitano una forza unidirezionale (nel senso che possono solo tirare e non spingere) e, per questo motivo, i movimenti pluridirezionali si realizzano sono se c’è un rapporto antagonista tra muscoli diversi. Per quanto riguarda la relazione tra arti e muscolo è importante sottolineare che in prossimità delle articolazioni ci sono 2 muscoli: uno estensore e uno flessore; la contrazione del primo provoca il raddrizzamento dell’arto mentre, in risposta alla contrazione del muscolo flessore, l’arto si flette. La contrazione muscolare può essere di 2 tipi: isometrica e isotonica. Poiché il prefisso iso indica uguale, per contrazione isometrica si intende un tipo di contrazione in cui non si verifica nessuna variazione nella lunghezza del muscolo; per contrazione isotonica si intende un tipo di contrazione in cui non si verifica nessuna variazione nella tensione del muscolo. Inoltre la contrazione isometrica si ha in attività come il mantenimento della postura; quella isotonica si ha nelle attività relative al movimento, come il camminare. Infine importante evidenziare che una contrazione isotonica viene sempre preceduta da una contrazione isometrica in quanto si può avere un accorciamento del muscolo solo quando la tensione prodotta è maggiore di quella opposto. Anche se da un punto di vista fisiologico si distinguono 3 tipi di muscolatura, in riferimento a questo discorso è opportuno soffermarsi sui muscoli striati. Il funzionamento dei muscoli striati scheletrici è controllato da fibre efferenti somatiche e la loro attivazione dipende dall’impulso nervoso che attraversa le fibre dei nervi motori, raggiunge il muscolo e lo attiva. I muscoli striati, poi, possono essere di due tipi: veloci e lenti. I muscoli veloci sono impiegati nei movimenti rapidi, come i movimenti oculari, e si contraggono in un tempo che va dai 10 ai 40 millesimi di secondo. Essi, di solito, sono rossi a causa dell’alta concentrazione di mioglobina (utile per immagazzinare ossigeno). I muscoli lenti si contraggono in un tempo pari a 100 millesimi di secondo, sono bianche e sono impiegati in attività come l’adattamento della postura. STRUTTURA DEL MUSCOLO E CONTRAZIONE Il muscolo è composto da un insieme di fasci di fibre muscolari; ogni fibra contiene miofibrille e ogni miofibrilla è formata da due mio filamenti: l’actina e la miosina, i quali si sovrappongono l’una sull’altra. I mio filamenti di actina sono più sottili, quelli di miosina sono più grossi e sono dotati di protuberanze a forma di peduncoli che creano un contatto con l’actina nel momento della contrazione. Infatti nel momento in cui un impulso nervoso innesca il potenziale d’azione nelle cellule muscolari ( che coincide con un aumento degli ioni calcio) questi ioni si legano alle molecole di troponina (che insieme ai filamenti di tropo miosina coprono – quando il muscolo è rilassata- i siti di legame tra l’actina e la miosina) che si modificano. Tali molecole modificate spingono verso l’interno dei siti la tropo miosina lasciando scoperti i siti di legami e permettendo ai peduncoli della miosina di legarsi all’actina, di ruotare e spingere tale filamento proteico in avanti. Man mano che l’actina avanza il peduncolo della miosina si piega sino a tornare alla sua posizione iniziale da dove ricomincia a spingere in avanti l’actina, dando luogo alla contrazione del muscolo mediante un processo chiamato modello di scorrimento dei filamenti. Per quanto riguarda il rapporto tra muscolo e impulso nervoso è fondamentale notare che all’interno delle fibre muscolari vi è una zona chimicamente sensibile: la placca terminale. Quando un motoneurone, collegato alla placca terminale, scarica viene prodotta acetilcolina attraverso la quale la fibra muscolare si depolarizza e si genera l’impulso elettrico che si propaga verso le estremità del muscolo; in risposta a tale impulso il muscolo si contrae. Un motoneurone può innervare può innervare più fibre muscolari e l’insieme di fibre muscolari innervate da un singolo motoneurone viene definito unità motrice. Generalmente i muscoli che controllano i movimenti fini (come il movimento delle dita delle mani) sono formate da fibre muscolari che appartengono a unità motrici piccole. Per effettuare dei movimenti rapidi e articolati, il sistema nervoso centrale deve possedere una serie di informazioni sia sui muscoli che sulla posizione degli arti ai quali essi sono legati. Il sistema nervoso riceve tali informazioni da due recettori sensoriali: - i fusi muscolari, che rilevano la variazione della lunghezza del muscolo sia durante la contrazione che durante lo stiramento; - gli organi di Golgi, che – invece- rilevano lo stato di tensione del muscolo. Il fuso muscolare è costituito da un insieme di fibre muscolari più sottili e più corte delle fibre ordinarie; inoltre sono riunite insieme e contenute in una capsula di tessuto connettivo che da vita ad una struttura a forma di fuso. Le fibre muscolari si distinguono in: fibre muscolari intrafusali, le quali formano il fuso muscolare; e le fibre muscolari extrafusali, che sono contenute nel muscolo. Il fuso muscolare è importante perché trasmette le informazioni dallo stato del muscolo attraverso le vie sensoriali afferenti che si avvolgono intorno alle fibre muscolari intrafusali e, fuoriuscendo dalla capsula di tessuto connettivo, raggiungono il midollo spinale. Inoltre quando il muscolo viene striato, cioè si allunga, anche il fuso muscolare si stria, provocando un aumento della frequenza di scarica della fibra sensoriale afferente; quando il muscolo si contrae, cioè si accorcia, diminuisce la tensione all’interno del fuso e diminuisce la frequenza di scarica della fibra sensoriale afferente. Da ciò si evince che il fuso muscolare è il recettore direttamente coinvolto nel riflesso di stiramento che viene anche definito riflesso fusale. L’importanza che i recettori sensoriali dei muscoli ricoprono nell’esecuzione del movimento è stata messa in luce, molti anni fa, dai lavori di Sherrington il quale dimostrò che l’isolamento dei recettori muscolari causa delle paralisi nonostante il mantenimento delle connessioni dai motoneuroni ai muscoli. In questo modo è stato dimostrato che tali recettori trasmettono le informazioni sullo stato dei muscoli ai centri motori del midollo spinale, della corteccia cerebrale e del cervelletto e che, in assenza di queste informazioni, è impossibile inviare qualsiasi comando motorio al muscolo. Infine va detto che i sistemi motori devono controllare sia i muscoli collegati all’articolazione coinvolta nel movimento, sia gli altri muscoli che agiscono su altre articolazioni. Un esempio relativo alla complessità del controllo messo in atto dai sistemi motori si riferisce all’azione che deve essere esercitata sui muscoli antagonisti: infatti nel momento in cui bisogna compiere un’azione non è sufficiente controllare solo i muscoli agonisti, ma anche quelli antagonisti e in quest’ultimo caso tale controllo non avviene in maniera fissa perché mentre in alcuni casi questi devono essere solo rilasciati, in altri casi (come i movimenti veloci) essi devono essere contratti nell’ultima parte del movimento. MOVIMENTO E GRADI DI LIBERTA’ Un aspetto importante inerente il controllo dei sistemi motori riguarda il problema dei gradi di libertà (ossia del numero di possibilità che si hanno per compiere uno stesso movimento). Ad esempio se vogliamo raggiungere la penna posta sul tavolo di fronte a noi possiamo seguire numerose traiettorie, anche perché in un movimento semplice come questo vengono impiegate numerose articolazioni: quella della spalla, del gomito e del braccio. Di conseguenza è possibile raggiungere la penna compensando tra loro gli angoli di apertura delle tre articolazioni oppure si possono mantenere fisse le articolazioni del gomito e del polso e raggiungere la penna con un ampia rotazione della spalla o si può mantenere fissa l’articolazione del gomito e raggiungere la penna mediante un’ampia flessione del gomito e del polso. Questo esempio dimostra che per ogni movimento esiste un altissimo numero di gradi di libertà. Tutto ciò è stato dimostrato sia in fisiologia, grazie agli studi di Bernstein, sia nei collegi dell’800 dove gli educatori obbligavano frequentemente gli allievi ad usare le posate sorreggendo un libro sotto le spalle. Berstein invece ha ipotizzato che per ridurre i gradi di libertà che intervengono nell’esecuzione di un movimento, il sistema motorio agisce compattando dei gradi di libertà appartenenti ai gruppi di muscoli coinvolti nello stesso movimento. ORGANIZZAZIONE GERARCHICA DEI SISTEMI MOTORI Il movimento viene effettuato tramite una contrazione indotta nel muscolo da impulsi nervosi, i quali sono prodotti dai motoneuroni. Tuttavia questa è solo la tappa finale di un processo che viene pianificato, regolato e controllato da tre sistemi distinti: il sistema spinale, il sistema tronco encefalico e il sistema corticale, che si interconnettono tra loro nel generare il movimento. Infine oltre a questi tre sistemi una grossa importanza, nella regolazione dell’attività motoria, è rivestita dai gangli della base e dal cervelletto. CONTROLLO MOTORIO A LIVELLO SPINALE Il midollo spinale è contenuto all’interno del canale midollare della colonna vertebrale e si collega alla periferia mediante i nervi motori e i nervi sensoriali. In corrispondenza della giunzione tra i vari corpi vertebrali due tipi di nervi fuoriescono dalle radici posteriori (o dorsali) e dalle radici anteriori (o ventrali). Attraverso le prime arrivano al midollo spinale le informazioni sensoriali ed è per questo che sono dette anche radici-sensitive; mentre le seconde sono impegnate per trasmettere degli impulsi ai muscoli ed è per questo che vengono definite anche radici-motorie. Nel momento in cui si attiva un muscolo il fuso muscolare riceve un impulso che viene rilevato dalle radici posteriori e, da qui, giunge nella sostanza grigia presente nell’encefalo. Nella sostanza grigia viene attivato il motoneurone che trasmette l’impulso al muscolo, che così si contrae. Mediante l’innervazione reciproca- invece- gli impulsi che hanno inizio nel fuso neuromuscolare non arrivano solo al motoneurone che innerva lo stesso muscolo estensore, ma anche ad un interneurone che inibisce il motoneurone del muscolo flessore antagonista. All’inizio dell’800 due studiosi: Bell e Magendie scoprirono che c’è una distinzione tra le vie sensoriali e le vie motorie, le quali hanno la caratteristica di essere indipendenti. In particolar modo Magendie ha dimostrato che in seguito della sezione delle radici posteriori lombo-sacrali il cane perdeva la sensibilità dell’arto posteriore corrispondente, ma non la motilità. In seguito alla sezione delle radici anteriori ventrali – invece- la zampa diventava completamente paralitica ma non perdeva la sensibilità. Dall’analisi di tutto ciò si evince che il midollo spinale è la sede dei circuiti responsabili delle risposte motorie più automatizzate e stereotipate e, in più, rappresenta il livello gerarchico più basso dell’organizzazione degli schemi motori che è costituita: dal midollo spinale, dal tronco dell’encefalo, dalla corteccia motoria e dall’area premotoria. Da un punto di vista neurofisiologico l’organizzazione degli schemi motori è stata definita gerarchica perché le strutture filogeneticamente più recenti non si sono formate attraverso la riorganizzazione delle strutture già esistenti, ma in seguito ad una sovrapposizione di sistemi di azione e di controllo più efficienti e più sofisticati, stabilendo quindi una gerarchia tra sistemi motori più antiche e quelli più recenti. Tuttavia il funzionamento dei sistemi motori gerarchicamente inferiori mantiene la capacità di operare autonomamente, ossia di controllare elementari atti motori senza l’intervento delle strutture cerebrali più elevate. L’autonomia del sistema motorio spinale è stata dimostrata con esperimenti sugli animali, tra i quali i più famosi sono stati: l’animale spinale e l’animale decerebrato. Nell’animale spinale è stata effettuata una sezione chirurgica ad un qualsiasi livello della colonna vertebrale in modo che il modello spinale risulti separato dall’encefalo. La logica sottostante è che l’eventuale attività motoria residua nella muscolatura controllata dai neuroni midollari che si trovano al di sotto della lesione non può essere comandata dai centri motori superiori, ma solo sai circuiti motori che si trovano nella parte del midollo spinale che si trova al di sotto della sezione chirurgica. Nell’animale decerebrato è stata effettuata una sezione a livello del tronco dell’encefalo, e in particolar modo tra i collicoli superiori e i collicoli inferiori; in questo secondo caso la finalità dell’intervento è lo studio dell’attività motoria in assenza del controllo volontario corticale e dell’azione inibitoria dei nuclei della base. In questa condizione permangono attività motorie semplici, come i riflessi e una generale coordinazione locomotoria, ma il quadro è dominato da uno stato spastico dei muscoli antigravitari dando luogo alla rigidità da cerebrazione . L’attività motoria spinale può realizzarsi seguendo 2 vie diverse: una monosinaptica, in cui il segnale sensoriale in entrata fa sinapsi direttamente sul motoneurone; una multisinaptica in cui il segnale sensoriale in entrata giunge al motoneurone dopo una lunga catena di sinapsi interneurali. Come in qualsiasi altro sistema nervoso centrale coinvolto nell’attività motoria, il sistema motorio del midollo spinale può operare in maniera adeguata soltanto se riceve informazioni afferenti e, in questo caso, le informazioni afferenti (che porta, che conduce) sensitive e le efferenze motorie che sfociano nell’attività muscolare risulta particolarmente stretta perché i recettori muscolari sono direttamente collegati con i motoneuroni. I movimenti riflessi sono comportamenti che fanno parte della struttura stessa del sistema nervoso centrale e che vengono messi in atto dai neuroni situati nella parte filogenetica più antica. Tuttavia esistono anche riflessi acquisiti, i quali coinvolgono i livelli più elevati della gerarchia dei sistemi motori e vengono appresi per permettere all’organismo di rispondere più velocemente e in maniera più adeguata alle richieste ambientali. CONTROLLO MOTORIO A LIVELLO DEL TRONCO DELL’ENCEFALO Il tronco dell’encefalo è il secondo elemento nell’organizzazione gerarchica dei sistemi motori e rappresenta la sede dei circuiti neurali deputati al controllo dei movimenti oculari, dell’equilibrio e di altri movimenti semplici. Per quanto riguarda l’attività motoria la funzione principale del tronco dell’encefalo è il controllo della motilità di sostegno che può avvenire grazie alle informazioni che provengono dall’organo dell’equilibrio. Notando che la postura e l’equilibrio vengono mantenuti senza nessun intervento cosciente è possibile affermare che i centri motori tronco encefalici sono sede di attività motorie riflesse e che a questo livello dell’organizzazione motoria, gruppi di neuroni formano dei specifici nuclei finalizzati al controllo di movimenti stereotipati o non coscienti. Un esempio di nuclei specifici sono i nuclei vestibolari i quali controllano il mantenimento dell’equilibrio. Il tronco dell’encefalo si divide in: midollo allungato, che si collega (in direzione caudale) al midollo spinale; ponte e mesencefalo, che (in direzione rostrale) si collega al diencefalo. Inoltre si può affermare che i più importanti centri motori tronco encefalici sono 3: - il nucleo rosso; - il nucleo vestibolare laterale; - alcuni settori della formazione reticolari. Per individuare la funzionalità motoria legata alle strutture tronco encefaliche (che creano numerose interconnessioni tra loro) sono stati fatti degli esperimenti sui gatti; esperimenti in cui sono state messe a confronto la postura e l’attività motoria in seguito alle sezioni chirurgiche effettuate a diversi livelli del tronco encefalico. Nell’animale decerebrato si osserva che, dopo aver sezionato il tronco dell’encefalo a livello del tentorio del cervelletto (tentorio= struttura anatomica che separa il cervelletto dai lobi occipitali), si manifesta un immediato aumento del tono della muscolatura estensoria e, di conseguenza, l’animale tiene le 4 zampe completamente estese e la testa ripiegata verso il dorso. Questa postura, definita rigidità da decerebrazione, è la conseguenza dell’effetto facilitante esercitato dal nucleo vestibolare (che non viene controbilanciato dagli effetti antagonisti e inibenti esercitati dal nucleo rosso o da altri centri motori superiori) sui motoneuroni estensori. Nell’animale mesencefalico la sezione del tronco dell’encefalo viene fatta in posizione più rostrale (ossia più anteriore) rispetto alla precedente, in modo che anche il mesencefalo ( oltre che il ponte e il cervelletto) possa rimanere legato ai centri spinali inferiori. La condizione posturale e l’attività motoria del gatto mesencefalico è decisamente migliore rispetto a quella dell’animale decerebrato dal quale si distingue per due motivi: prima di tutto l’animale mesencefalico non presenta una rigidità muscolare pari a quella dell’animale decerebrato; in secondo luogo l’animale mesencefalico è in grado di regolare il proprio tono muscolare. Da ciò si evince che se anche in entrambe gli animali sono interrotte le vie provenienti dai centri motori superiori, le condizioni del gatto mesencefalico sono migliori grazie all’azione dei centri motori del mesencefalo, tra i quali molto importante sembra essere il nucleo rosso. CONTROLLO MOTORIO A LIVELLO CORTICALE La corteccia cerebrale rappresenta il livello gerarchicamente più elevato nei sistemi di controllo motorio e le vie nervose che collegano la corteccia ai motoneuroni del livello spinale sono numerose e articolate. Nella corteccia cerebrale di tutti i mammiferi esiste una zona in cui una stimolazione elettrica provoca movimenti nel lato opposto del corpo. Una prima distinzione che si può fare tra le attività motorie integrate a livello spinale e tronco encefalico e quelle integrate a livello corticale è che le prime avvengono a livello subcosciente, mentre le seconde sono – in gran parte – movimenti volontari. Le aree della corteccia cerebrale più interessate al movimento sono 2: - la corteccia motoria o area 4 di Broadmann; - la corteccia premotoria o area 6 di Broadmann. La corteccia motoria o area 4 di Broadmann o corteccia piramidale (perché composta da un’alta concentrazione di cellule piramidali robuste) si trova davanti alla scissura di Rolando e si suddivide in diverse sottoaree in relazione ai diversi gruppi somatici a cui sono collegate. La rappresentazione dell’insieme delle aree specifiche forma l’immagine dell’homunculus di Woolsey e dei suoi colleghi (ossia di un uomo che mantiene le proporzioni dell’area della corteccia deputate al suo controllo), in cui non c’è un rapporto proporzionale tra la grandezza dell’area e la grandezza del muscolo, bensì un rapporto diretto tra la raffinatezza del movimento che il muscolo è in grado di compiere e l’ampiezza dell’area corticale (Esempio: l’area corticale che controlla i muscoli della gamba è meno estesa di quella che controlla i muscoli delle dita, del volto, della bocca o della lingua). Infine è importante sottolineare che la corteccia motoria rappresenta l’area corticale in cui avviene la conversione dei progetti di movimento in comandi motori i quali rendono possibile la realizzazione dei progetti stessi. L’area 6 di Broadmann o corteccia associativa o area premotoria è la seconda area corticale implicata nel controllo dell’attività motoria ed p situata anteriormente alla corteccia motoria. Questa corteccia associativa è responsabile della progettazione del movimento, dell’identificazione dei bersagli nello spazio e della scelta del decorso temporale dell’azione motoria e – a differenza dell’area motoria primaria- la sua stimolazione genera movimenti più complessi. Pertanto mentre la corteccia piramidale è responsabile di movimenti semplici e privi di scopo, la corteccia associativa è responsabile del controllo di movimenti coordinati i quali richiedono l’attivazione simultanea di più masse muscolari. Inoltre quest’ultima corteccia dispone di connessioni sottocorticali con la corteccia motoria e con le aree talamiche contigue e, in più, è collegata al talamo e ai nuclei della base. Di conseguenza il controllo dei movimenti complessi è il risultato dell’attività congiunta di strutture corticali e sottocorticali. Un’altra differenza che c’è tra area 4 e area 6 di Broadmann è visibile nei deficit provocati da lesioni nel cervello: infatti la lesione o l’asportazione di una parte dell’area 4 provoca la paralisi dei muscoli che vi sono rappresentati, anche se l’animale è ancora in grado di compiere dei movimenti grossolani. Quindi la lesione non provoca l’impossibilità di contrarre la muscolatura, ma viene meno la capacità di eseguire volontariamente i movimenti raffinati. Infine con una lesione o l’asportazione dell’area 4 i movimenti involontari e posturali restano inalterati. Grazie a questi esperimenti si è potuto confermare empiricamente che: l’area 4 è deputata ai movimenti volontari fini, mentre l’area 6 ai movimenti involontari e posturali. Come qualsiasi organo di controllo anche la corteccia motoria necessita di una serie di informazioni per poter operare adeguatamente; informazioni che le vengono convogliate da diverse vie afferenti: - le fibre che provengono dall’altro emisfero cerebrale e che connettono aree motorie corrispondenti; - le fibre sottocorticali provenienti dalle aree corticali somatosenistive che si trovano dietro all’area motoria primaria e dalle aree visive e uditive; - i fasci, provenienti dai nuclei ventrolaterali e ventroanteriori del talamo. Le vie efferenti che dalla corteccia trasmettono informazioni ai centri motori inferiori possono essere di due tipi: le vie piramidali o corticospinali, responsabili della mobilità del tronco e degli arti; le vie extrapiramidali le quali trasmettono i segnali motori al midollo spinale e terminano nella parte anteriore della sostanza grigia. Infine va sottolineato che la via piramidale porta la muscolatura verso un ipertono mentre la via extrapiramidale porta la muscolatura verso l’ipotono. CONTROLLO MOTORIO E GANGLI DELLA BASE I gangli o nuclei della base fanno parte del sistema motorio extrapiramidale e formano un’importante connessione sottocorticale tra la corteccia motoria e quella associativa. I principali gangli della base sono: il nucleo caudato; il putamen (che insieme al nucleo caudato forma il corpo striato); il globus pallidus; la sub stantia nigra e il nucleo subtalamico. Le vie afferenti ed efferenti collegate ai gangli della base sono numerose: tra le vie afferenti quelle principali provengono dalla corteccia associativa, dalla corteccia motoria e dai nuclei del talamo ed esse si connettono con il corpo striato. Le principali vie efferenti – invece- partono dal globus pallidus per poi giungere nel talamo; altre vie efferenti partono dal corpo striato e raggiungono la sub stantia nigra e il globus pallidus. Il ruolo svolto dai gangli della base risulta essere molto importante in quanto questi organi svolgono funzioni motorie di altissimo livello e, addirittura, vicarianti le funzioni svolte dalla corteccia cerebrale. A queste conclusioni si è giunti mediante una serie di esperimenti condotti sia sugli animali (e in particolare sui gatti) che sugli uomini. Per quanto riguarda gli studi condotti sugli uomini essi hanno messo in evidenza che i danni corticali compromettono molti movimenti ( e in particolare quelli fini) ma non la possibilità di compiere movimenti automatici, di camminare e di mantenere l’equilibrio. La distruzione del nucleo caudato – invece- provoca la paralisi di tutto l’emisoma opposto al lato della lesione (facendo eccezione per i riflessi spinali o tronco encefalici). I gangli della base operano a stretto contatto sia con la corteccia cerebrale sia con il cervelletto, ma nonostante ciò le funzioni svolte da alcuni dei nuclei descritti possono essere parzialmente isolate. Ad esempio il nucleo caudato e il putamen sono i responsabili del’avvio e della regolazione dei movimenti grossolani intenzionali; il globus pallidus provvede al tono muscolare necessario per eseguire i movimenti intenzionali ( sia quelli provenienti dalla corteccia motoria; sia quelli avviati dalla corteccia; sia quelli avviati dal corpo striato). Questa funzione è molto importante soprattutto per i movimenti fini in quanto qualsiasi movimento fine richiede un aggiustamento precedente della posizione delle varie parti del corpo. Delle lesioni al globus pallidus impediscono l’aggiustamento di queste parti del corpo e, di conseguenza, l’esecuzione di movimenti fini. Infine le cellule nervose del putamen dimostrano un’attività ridotta nell’esecuzione di movimenti rapidi; pertanto ciò indica che tali strutture vengono coinvolte nella programmazione e nel controllo di movimenti lenti. CONTROLLO MOTORIO E CERVELLETTO Il cervelletto è una struttura nervosa molto antica (probabilmente la prima che si è specializzata nella coordinazione motoria) che interviene solo in relazione alle attività che sono state iniziate a livello spinale, tronco encefalico e corticale e che regola la coordinazione motoria. A differenza della corteccia motoria la stimolazione elettrica della corteccia del cervelletto non provoca nessuna contrazione muscolare ed è per questo motivo che per molti anni quest’organo non è stato preso molto in considerazione. Infatti molto recentemente la funzione svolta dal cervelletto è stata molto rivalutata in quanto una serie di studi hanno dimostrato che questa struttura cerebrale regola il tempo delle attività motorie veloci e – in generale- di tutte quelle attività che richiedono una rapidità di esecuzione. In più ulteriori studi hanno messo in evidenza che nelle attività rapide è fondamentale la sincronizzazione dei singoli movimenti che compongono l’azione e il cervelletto è la struttura anatomica che è maggiormente responsabile della componente temporale del movimento. Da un punto di vista anatomico il cervelletto si distingue in tre lobi: il lobo flocculo nodulare, che è la parte più antica ed è particolarmente deputato al controllo dell’equilibrio; il lobo anteriore e il lobo posteriore, il quale è il più sviluppato e forma 2 protuberanze dette emisferi cerebellari, i quali sono i punti di arrivo di un gran numero di vie nervose. Il ruolo principale del cervelletto è quello di confrontare lo stato attuale del muscolo e dell’arto coinvolto nel movimento con la situazione che deriva dall’attivazione del sistema motorio. Se le due condizioni sono diverse il cervelletto si attiva per trasmettere le correzioni al sistema motorio che ha il compito di incrementare, inibire o diminuire i livelli di attivazione dei muscoli. Per poter inviare al sistema motorio i segnali correttivi del movimento durante la sua esecuzione, il cervelletto deve ricevere un feedback sullo stato del muscolo in un tempo molto veloce ed è per questo che è necessario un sistema afferente molto rapido. Oltre ad un rapido sistema afferente il cervelletto deve essere dotato anche di un ottimo sistema efferente per poter trasmettere le correzioni al sistema motorio. Le vie efferenti del cervelletto coincidono con 3 circuiti: il primo è legato all’attività motoria volontaria; il secondo è coinvolto nel controllo dell’equilibrio e della postura; il terzo ha il compito di coordinare le altre due vie efferenti. Anche se grazie a numerosi studi si è giunti a conoscere e descrivere come i circuiti neurali trasmettono gli impulsi ai motoneuroni, ciò non risulta sufficiente per capire come il pensiero possa trasformarsi in un impulso capace di attivare cellule nervose che danno il via al movimento. Nonostante ciò la scienza non si è persa nel completo pessimismo: infatti molto importanti sono stati due esperimenti. 1) se viene chiesto di eseguire un movimento dopo la presentazione – in successione- di due stimoli, immediatamente prima dell’emissione della risposta si registra una lenta onda negativa nella corteccia cerebrale. Questa onda è stata denominata potenziale di aspettativa in quanto è un potenziale associato all’attesa del secondo stimolo la cui comparsa è preannunciata dallo stimolo che lo precede. 2) Se ad un partecipante ad un esperimento viene chiesto di compiere un movimento semplice ad intervalli temporali irregolari e scelti autonomamente, prima del movimento si registra su tutta la superficie della corteccia cerebrale un’onda negativa che aumenta lentamente. Quest’onda è stata definita potenziale di preparazione ed è stato notato che essa precede il momento in cui la corteccia motoria invia i comandi di attivazione ai muscoli scheletrici. Da ciò è possibile ammettere: primo, che il potenziale di preparazione può essere inteso come il correlato neurale della volontà di compiere un movimento; secondo, che se l’onda si sviluppa su tutto il cervello significa che la realizzazione di un’azione volontaria richiede la collaborazione di più (se non tutte) le aree della corteccia cerebrale. CAPITOLO II LE CLASSI DI MOVIMENTO Una distinzione specifica dei vari movimenti non può essere realizzata in quanto il numero dei movimenti è pari al numero dei ricercatori che si occupano di comportamento motorio. Infatti, in seguito ad una distinzione generale, è possibile distinguere 5 classi di movimenti: - i movimenti discreti: essi sono quei movimenti in cui può essere individuato con precisione sia l’inizio che la fine e degli esempi sono: il calciare, portare un oggetto alla bocca, dare uno schiaffo. Due caratteristiche principali di questi movimenti sono: 1) la velocità, la quale è quasi sempre maggiore della velocità osservata nei movimenti continui; 2) l’importanza dell’aspetto cognitivo rispetto all’aspetto motorio. Quest’ultimo punto può essere chiarito meglio con un esempio: se ad un addetto di una torre di controllo si dice di premere un pulsante verde nel momento in cui riceve un dato segnale e un pulsante rosso nel momento in cui riceve un segnale d’emergenza, l’impegno maggiore dell’addetto è quello di riuscire a discriminare i due segnali e selezionare il pulsante corretto, mentre è poco importante come preme il pulsante o con quale dito lo pigia. - I movimenti continui: essi sono quelli in cui non è possibile individuare esattamente il momento d’inizio e quello della fine e, in questo caso, il tempo di esecuzione è più lungo di quello impiegato nei movimenti discreti. Esempi di questi movimenti sono: la corsa, il nuoto - I movimenti seriali: essi possono essere definiti come una somma di più movimenti discreti legati tra loro. In questo caso anche se può essere individuato il momento d’inizio e il momento della fine, il tempo d’esecuzione può essere anche molto lungo in quanto è il risultato della concatenazione di più movimenti discreti. Esempi di questi movimenti sono: la preparazione del caffè, allacciarsi le scarpe, cucire a macchina… I movimenti si distinguono, poi, in movimenti aperti e chiusi in base alla classificazione delle abilità motorie proposta da Poulton. Tuttavia questa classificazione più che riguardare i movimenti in sé, fa riferimento al contesto in cui i movimenti vengono messi in atto. - I movimenti aperti sono quelli che si realizzano in un contesto che presenta un alto numero di variabili che non possono essere controllate. Tipici esempi di movimenti aperti si hanno in tutte quelle attività sportive in cui l’atleta è opposto ad un avversario e, di conseguenza, egli si ritrova nella condizione in cui nulla può essere programmato in anticipo perché la sua esecuzione deve adattarsi ai movimenti dell’avversario. Ne deriva che il successo o meno nei movimenti aperti dipende in massima parte dalla capacità e dalla velocità di adattamento all’ambiente esterno, ossia dal repertorio di programmi motori posseduti e dalla capacità di modificare in tempi brevi i valori delle variabili di movimento specificate all’interno del programma stesso. - I movimenti chiusi sono quelli che si realizzano in un ambiente stabile, cioè in un ambiente in cui le variabili sono costanti e controllabili. Nell’ambito delle attività sportive i movimenti chiusi si hanno nel bowling, nelle gare di ginnastica e in tutte quelle attività in cui le variabili esterne sono fisse e, di conseguenza, possono essere previste e controllate. Data la prevedibilità ambientale, il requisito fondamentale per eseguire un movimento in modo adeguato coincide con la corretta programmazione dell’azione; mentre l’insuccesso può essere dovuto o alla scelta di un programma motorio non idoneo allo scopo che vogliamo raggiungere, o alla scelta di un programma motorio corretto ma all’interno del quale ci sono valori errati. METODI DI MISURAZIONE I metodi impiegati per la misurazione dei movimenti si distinguono in due gruppi: 1) da un lato ci sono quei metodi che si propongono di osservare e descrivere le caratteristiche intrinseche al movimento; 2) dall’altro ci sono quei metodi che misurano l’effetto che il movimento produce, ossia le conseguenze dell’azione messa in atto. I metodi di misurazione appartenenti al primo gruppo derivano dalla cinematica, una branca della meccanica che studia le proprietà del moto indipendentemente dalle cause che lo producono. Applicata al controllo motorio la cinematica si occupa delle diverse posizioni che il corpo o alcuni segmenti corporei possono assumere nello spazio. METODI DI MISURAZIONE DELLE CARATTERISTICHE DEL MOVIMENTO I metodi di misurazione legati alle caratteristiche del movimento sono particolarmente due: la registrazione cinematica e la registrazione elettromiografica. La registrazione cinematica è un metodo usato per rilevare le diverse posizioni spaziali che compongono la traiettoria di un movimento e le successive posizioni occupate dai segmenti corporei durante il movimento. Per la rilevazione dello spazio occupato dall’arto si possono utilizzare varie tecniche, come la ripresa cinematografica la quale consiste nel compiere delle osservazioni molto dettagliate su una specifica parte del corpo per verificare le modificazioni spaziali x unità di tempo; in questo modo si riesce a ricostruire la traiettoria seguita e i tempi impiegati per ogni segmento della traiettoria. Un’altra tecnica simile è quella dei leds, i quali vengono applicati in varie parti del corpo e forniscono la successione degli spostamenti eseguiti dall’arto osservato e la completa traiettoria percorsa durante il movimento. Con questo metodo possono essere misurate anche altre variabili come: la velocità e l’accelerazione nei vari momenti d’esecuzione dei movimenti. La registrazione elettromiografica è un metodo che parte dal presupposto che ogni movimento comporta attività muscolare la quale può essere rilevata, amplificata e misurata. Ne consegue che i movimenti possono essere suddivisi in base al tipo di attività muscolare necessaria per la loro esecuzione. Questa tecnica è importante per tre motivi. 1) oltre ad individuare quali muscoli sono impiegati in un dato movimento essa indica la forza che il muscolo ha prodotto per eseguire un movimento; 2) la registrazione dell’attività muscolare permette di individuare la successione e la durata di attivazione e inibizione dei muscoli agonisti e antagonisti implicati nel movimento; 3) infine la misurazione elettromiografica è importante perché è l’indice più preciso riguardo al momento d’inizio del movimento. Confrontando la misura elettromiografica con il tempo di reazione si nota che il momento della prima contrazione muscolare (tempo di reazione motoria) precede di un tempo abbastanza lungo la reazione motoria o tempo di reazione, il quale è rilevabile al momento della risposta motoria osservabile. Invece la misura dell’attività elettrica del muscolo può essere rilevata nel momento in cui c’è la prima contrazione muscolare, ossia quando il movimento è già innescato ma non è osservabile. MISURAZIONE DELL’AMPIEZZA DEL MOVIMENTO Alcuni metodi di misurazione del movimento si propongono di rilevare gli effetti che esso provoca sull’ambiente esterno, e in questo caso l’esecuzione del movimento viene misurato lungo 3 parametri principali: l’ampiezza, il tempo impiegato e l’errore commesso. AMPIEZZA: L’ampiezza del movimento è il più macroscopico dei parametri presi in considerazione per misurare il risultato dell’attività motoria e degli esempi sono: l’altezza dell’asticella nel salto in alto o la distanza che deve essere percorsa dal braccio per prendere un libro riposto su una libreria. Anche se questo parametro è molto importante esso non è stato studiato in maniera estesa e approfondita dagli scienziati in quanto affrontare lo studio del movimento sotto questo punto di vista è difficile perché bisogna prendere in considerazione una serie di variabili difficilmente separabili tra loro e, di conseguenza, diventa complicato saper individuare quella responsabile della prestazione. Tuttavia è sbagliato ritenere l’ampiezza del movimento una misura troppo generale e grossolana: infatti tale considerazione si basa sull’idea secondo la quale per migliorare la prestazione è sufficiente migliorare la potenza muscolare della prestazione stessa. È, invece evidente, che si può ottenere un miglioramento della prestazione anche attraverso meccanismi cognitivi. Anche se nel corso della storia gli scienziati non si sono impegnati a studiare il parametro dell’ampiezza, un eccezione è stata fatta da Bachman, il quale ha ideato un compito motorio in cui venivano testate le capacità di equilibro e di coordinazione motoria. I soggetti sottoposti a questo studio dovevano salire una scala a pioli sfalsati e non appoggiata alla parete sino a quando erano in grado di a mantenere l’equilibrio; in quel momento veniva misurata l’ampiezza del movimento effettuato sino a quel punto, ossia sino al piolo raggiunto. La scala di Bachman può essere vista come un buon esempio di incremento di incremento dell’ampiezza del movimento e, in più, ha confermato che il miglioramento dell’ampiezza è il risultato di un progressivo apprendimento della coordinazione e dell’equilibrio e non di un miglioramento della potenza muscolare. MISURAZIONE DEL TEMPO DI MOVIMENTO E DEL TEMPO DI REAZIONE Un altro modo per testare la maggiore destrezza nell’esecuzione di un movimento è quello di misurare il tempo impiegato ad eseguirlo o tempo di movimento. Infatti oltre all’ampiezza, una variabile che può differenziare due prestazioni motorie è proprio il tempo impiegato per la loro esecuzione. Se prendiamo in considerazione la scala di Bachman, misurando il tempo che un soggetto impiega per raggiungere il terzo piolo si ottiene una stima del tempo necessario per eseguire quel movimento. Un altro metodo per studiare la velocità del movimento è quello di misurare il numero delle volte in cui il movimento viene ripetuto nel corso di un’unità di tempo stabilita: ad esempio quante volte si riesce a battere le mani in 20 secondi. In questo caso se si divide il tempo totale (20 sec) per il numero delle volte in cui le mani sono entrate in contatto, si ottiene il tempo medio di movimento di ogni singola battuta. Evidentemente, a parità di altre condizioni, il tempo di movimento sarà funzione dell’ampiezza del movimento stesso in quanto il tempo impiegato dalle mani x venire in contatto sarà più lento quanto maggiore sarà la distanza tra le due mani. Il tempo di reazione, a differenza del tempo di movimento, non risente dell’ampiezza del movimento e si identifica con il tempo necessario per decidere di eseguire un dato movimento. Più specificamente, il tempo di reazione misura l’intervallo che va dalla presentazione della stimolazione sensoriale (input) alla prima reazione osservabile del movimento stesso (output). Esempio: se ad un soggetto viene chiesto di spostare una leva verso dx in risposta ad una luce rossa e verso sx in risposta ad una luce verde, per una distanza di 10 cm, il tempo di reazione sarà l’intervallo di tempo che va dalla presentazione della luce alla prima variazione della posizione della leva; mentre il tempo di movimento sarà quello che intercorre tra il tempo di reazione (ossia dal primo mutamento della leva) e il momento in cui il movimento della leva è stato completato. È importante sottolineare che il tempo di reazione è composto da due fasi: la prima fase è quella centrale-cognitiva, durante la quale hanno luogo i processi di elaborazione e di codifica dell’informazione sensoriale in entrata sino allo stadio di selezione della risposta da effettuare. La seconda fase è quella periferico-motoria, durante la quale si realizza l’attività muscolare dalla prima contrazione registrabile con l’elettromiografo sino al primo mutamento osservabile esternamente (ossia sino all’inizio del movimento). Da ciò possiamo dedurre che il tempo totale dell’esecuzione di un movimento è caratterizzato da diversi stadi: 1) tempo di reazione premotorio, che coincide con l’intervallo di tempo che va dalla presentazione dello stimolo al primo segnale elettromiografo registrabile dall’elettrodo posto sul muscolo impiegato nel movimento; 2) tempo di reazione motorio, il quale coincide con l’intervallo che va dal primo mutamento rilevabile del muscolo all’inizio del movimento; 3) il tempo di reazione, con il quale si intende l’intervallo di tempo che intercorre tra la presentazione dello stimolo e l’inizio della risposta motoria; 4) e, infine, il tempo di movimento che indica l’intervallo tra l’inizio del movimento e il suo completamento. In generale, il presupposto dalla quale si parte nella misurazione temporale dell’attività motoria è che quanto più elementari sono i processi cognitivi coinvolti nell’elaborazione dello stimolo, tanto più veloci sono i tempi di reazione del soggetto. Allo stesso modo, tanto più semplice è il movimento di risposta richiesto, tanto più rapido è il tempo della sua esecuzione. Mentre sembra ovvio che due movimenti diversi richiedono tempi di esecuzione diversi, lo sembra meno che in alcune condizioni il tipo di movimento può influenzare il tempo di reazione. Su questo ultimo aspetto si è soffermato Klapp, che con un suo studio ha messo in evidenza che dei movimenti difficili possono provocare un rallentamento nel tempo di reazione; e ciò accade particolarmente nel momento in cui tale movimento viene programmato in anticipo. Inoltre la programmazione dipende dall’ampiezza e dalla precisione del movimento stesso ed è per questo che nel caso di un movimento ampio e per il quale è richiesta un’ampia precisione, non è possibile compiere una perfetta programmazione in anticipo perché la correttezza del movimento si basa sul feedback visivo che permette degli aggiustamenti continui durante l’esecuzione. Invece, dei movimenti che vengono eseguiti in tempi veloci non permettono di elaborare un feedback visivo ed è per questo che devono essere programmati in anticipi. Dall’analisi di ciò si evince che Klapp ha ipotizzato che i movimenti ad ampiezza e precisione elevata non provocano un rallentamento del tempo di reazione; mentre i movimenti molto veloci generano un rallentamento del tempo di reazione. Per affermare questa sua ipotesi Klapp ha realizzato un esperimento i cui risultati hanno messo in evidenza che: 1) per i movimenti molto veloci tutti i parametri devono essere specificati e programmati in anticipo perché durante l’esecuzione del movimento non c’è tempo per effettuare una qualsiasi correzione. 2) per quanto riguarda la programmazione, tanto maggiore è la precisione richiesta, tanto più accurato dovrà essere il programma e maggiore il tempo necessario alla sua costruzione. Inoltre siccome il programma deve essere completato prima dell’inizio del movimento, il tempo della sua preparazione rientra nel tempo di reazione ed è per questo che i programmi motori più complessi richiedono tempi di reazione più lunghi. 3) per i movimenti lenti – invece- la traiettoria può essere guidata dal feedback visivo e anche gli errori possono essere corretti durante l’esecuzione del movimento stesso. Di conseguenza, in questo caso, non risulta necessaria la costruzione di un programma motorio. Infine ci sono alcuni studiosi i quali pensano che il tempo di reazione e il tempo di movimento dipendono anche da fattori inerenti l’organismo di chi effettua la prestazione. Un lavoro molto particolare è stato quello di Anson, il quale ha affermato che l’effettore corporeo con il quale si effettua il movimento sia irrilevante per il tempo di reazione. E un discorso simile vale anche per l’età, la quale non sembra incidere in modo significativo sui tempi di reazione. Una posizione diversa è stata presa dalla Hodgkings, la quale ha condotto degli studi su soggetti diversi di età compresa tra i 6 e gli 83 anni e ha dimostrato che una riduzione dei tempi di reazione e di movimento si registra nel periodo che va dai 6 ai 20 anni; mentre dopo i 30 anni si osserva una regolare tendenza al rallentamento di entrambe i parametri. METODI DI MISURAZIONE DELL’ERRORE DI MOVIMENTO Un altro modo per misurare l’abilità e la destrezza nell’esecuzione di un movimento è quello che misura gli errori commessi. Ogni movimento affinchè possa essere considerato corretto deve essere accurato, cioè deve essere eseguito secondo precisi parametri spazio-temporali. Se, durante l’esecuzione del movimento, si esce fuori dai calori di tolleranza ammessi si ha l’errore e il mancato raggiungimento dello scopo previsto. I principali metodi di misurazione dell’errore sono quattro: 1) il metodo dell’errore costante; 2) il metodo dell’errore variabile; 3) il metodo della variabilità totale; 4) il metodo dell’errore assoluto. L’errore costante è uno dei metodi che permette di valutare l’accuratezza del movimento misurando l’errore medio che viene commesso durante ripetute esecuzioni. Tuttavia anche se esso fornisce un’accurata misura della prestazione ottenuta nell’esecuzione del movimento, non è in grado di fornire una misura attendibile della distribuzione dei risultati nelle diverse prove. Questo aspetto viene definito disuguaglianza delle risposte e mentre qui viene ignorato, nel metodo dell’errore variabile viene preso in considerazione. L’errore variabile misura la variabilità relativa alla media delle prove effettuate, ed è per questo che viene riconosciuto come indice della disuguaglianza. Inoltre, siccome questo metodo tiene conto della variabilità delle risposte, il suo valore sarà tanto alto quanto maggiore è la variabilità; tanto minore quanto più le risposte risulteranno omogenee. Nonostante la sua grande efficienza, anche questo metodo di misurazione ha una pecca: non tiene conto della distanza che c’è tra ogni singola prova e la prestazione corretta; parametro che – invece- viene presa in considerazione dal metodo dell’errore costante che, però, ignora la variabilità delle prove. La variabilità totale è un metodo che si ottiene dalla combinazione dell’errore variabile e dell’errore costante ed essa viene considerata la misura più affidabile dell’errore di movimento. Molto spesso il valore della variabilità totale e quello dell’errore variabile sono simili, ma nel caso in cui i dati sono molto distanti dalla prestazione corretta (anche se sono omogenei) i due metodi forniscono risultati diversi. Infine, l’ultimo metodo di misurazione dell’errore, è quello dell’errore assoluto, il quale è composto da elementi che hanno lo stesso significato di quelli dei metodi precedenti ed è molto simile a quello della variabilità totale perché entrambe si basano sulla differenza tra la prova effettuata e la prova corretta. Nonostante il metodo dell’errore assoluto sia quello più usato in letteratura sull’errore di movimento, Schutz e Roy hanno messo in luce alcune riserve relative alla sua correttezza. Questi autori, a tal proposito, hanno sostenuto che nella formula dell’errore assoluto l’errore costante e l’errore variabile si combinano in modo complesso e, di conseguenza, può essere difficile riconoscere l’influenza apportata dai due fattori. Mentre nella formula della variabilità totale viene effettuata una semplice somma dei due aspetti dell’errore di movimento e quindi la combinazione tra errore costante ed errore variabile è più chiara. CONSIDERAZIONI GENERALI SUI METODI DI MISURAZIONE DELL’ERRORE Dall’analisi di quanto detto sin’ora le principali componenti dell’errore sono due. 1) la misura dello scostamento tra la prestazione eseguita e la prestazione corretta; 2) la misura della variabilità tra le varie prestazioni eseguite. Se da un lato non si può affermare che una componente di errore può essere più importante di un’altra, da un punto di vista applicativo- invece- è importante individuare quali delle due misure (errore costante ed errore variabile) è la più utile nella valutazione dell’errore nell’esecuzione di un dato movimento. Se due atleti di tiro con l’arco raggiungono lo stesso risultato, misurato con il metodo della variabilità totale, e bisogna selezionarne uno, è meglio scegliere il primo, che ha un’alta variabilità tra le prove e un errore costante; o il secondo, che fornisce risposte omogenee tra loro ma con un alto errore costante? La scelta dovrebbe ricadere sul secondo atleta in quanto nel miglioramento della prestazione l’omogeneità delle prestazioni è più importante della costanza dell’errore. Infatti mentre al primo atleta manca la coordinazione percettivo-motoria necessaria per il compito, il secondo atleta possiede il corretto programma di movimento che deve essere solo tarato per migliorare la prestazione. Infine una serie di studi sull’apprendimento hanno dimostrato che mentre il miglioramento dell’errore costante viene ottenuto fin dalle prime prove, quello dell’errore variabile si ottiene dopo un lunghissimo allenamento ( e spesso non viene ridotto in maniera significativa). CAPITOLO III CONTROLLO MOTORIO A CIRCUITO CHIUSO Con l’espressione “controllo motorio” si indica, da un lato (volendo utilizzare una definizione di Schmidt) l’area di ricerca che si occupa dello studio di aspetti fisiologici, fisici e psicologici del movimento; dall’altro indica lo studio della posture e dell’equilibrio, e in particolare l’indagine che la mente e il corpo svolgono per governarli. Nella max parte dei casi la postura e il movimento sono determinati e guidati dall’effetto concomitante di feedback periferici e programmi motori centrali. Pertanto l’attività motoria viene continuamente controllata e corretta grazie al confronto tra pianificazione centrale e percezione sensoriale periferica. Per quanto riguarda il ruolo che il controllo motorio esercita sull’esecuzione del movimento, non esiste nessun tipo di divergenza; mentre una serie di problemi cominciano a nascere nel momento in cui si cerca di spiegare il modo in cui tale controllo viene esercitato. Su quest’ultimo punto sono nate una serie di opinioni che, ancora oggi, risultano essere divergenti ma una delle ipotesi che ha avuto maggiore spessore nel corso della storia è quella che afferma che esistono dei modelli di funzionamento (e cioè mediante i quali il sistema nervoso centrale organizza, produce e controlla il movimento) che si distinguono in due categorie: il modello a circuito chiuso e il modello a circuito aperto. Il controllo motorio a circuito chiuso o teoria del controllo periferico si basa sulle informazioni sensoriali che dalla periferia vengono trasmesse al sistema centrale, il quale trae – da questi feedback- delle informazioni utili x verificare se il movimento viene compiuto in modo corretto. Se, eventualmente, non è così il sistema nervoso centrale effettua delle correzioni sul movimento in corso. L’atto di nascita del modello a circuito chiuso può essere fatto risalire al lavoro di Mott e Sherrington, i quali dimostrarono il ruolo determinante che le sensazioni muscolari e cutanee ricoprivano per la corretta esecuzione del movimento di un arto di una scimmia. Il controllo motorio a circuito aperto o teoria del controllo centrale sostiene che il feedback periferico non nessuna influenza sulla corretta esecuzione del movimento in quanto il sistema è capace di specificare tutte le informazioni necessarie all’esecuzione del movimento in un programma. Ciò significa che una volta avviato il programma il movimento è in grado di essere portato a termine senza l’intervento di informazioni provenienti dai centri periferici. Questo modello è stato ideato, per la prima volta, da Lashley il quale verificò la presenza di movimenti corretti anche in totale assenza di feedback periferici. IL CONTROLLO MOTORIO A CIRCUITO CHIUSO La teoria del controllo periferico del movimento si basa si un principio fondamentale: il feedback, in quanto il controllo dei movimenti sensoriali avviene (secondo questo modello) grazie ai feedback inviati dai recettori periferici che permettono all’organismo di rilevare gli errori commessi e correggerli. Il circuito che si viene a formare tra: azione da compiere, rilevamento del primo movimento effettuato, relativo feedback retroattivo, confronto tra movimento pianificato e movimento eseguito, conseguente modificazione dell’azione, da luogo al circuito chiuso i cui componenti possono essere isolati e il quale necessita dell’informazione periferica relativa al livello di prestazione raggiunta. Questa informazione viene poi confrontata con lo scopo iniziale, la parte esecutiva rileva eventuali errori e, se questi ci sono, invia i comandi di correzione alla parte effettrice la cui preparazione darà luogo al feedback e così via secondo uno schema a circuito chiuso. ESEMPIO: Se vogliamo muovere la mano da un punto A ad un punto B, il sistema motorio (parte esecutiva) inizia il movimento inviando i comandi ai muscoli interessati ( che coincidono con la parte effettrice). L’attività dei muscoli da luogo ad un feedback, che informa la parte esecutiva sul movimento che si sta attuando; la parte esecutiva analizza il feedback e, in caso di discrepanza tra il movimento voluto e quello attuato, verifica l’errore e modifica i comandi inviati ai muscoli. Pertanto il feedback può essere definito come un’informazione sensoriale relativa al risultato dell’atto motorio; ma nonostante ciò esistono diverse informazioni che, anche se non si riferiscono strettamente all’atto motorio, possono assumere grande importanza x il controllo del movimento. I recettori che rilevano e trasmettono queste informazioni sono: i propriocettori, i quali inviano al sistema informazioni relative alla posizione del corpo e dei singoli arti nello spazio. Esempi di propriocettori sono i fusi muscolari e gli organi tendinei del Golgi; gli esterocettori, che forniscono informazioni relative agli oggetti che si muovono o che vengono a trovarsi nelle immediate vicinanze del corpo. Esempi di esterocettori sono: i recettori della pelle, dell’occhio, dell’orecchio, le papille gustative. L’insieme delle informazioni provenienti dai propriocettori e dagli esterocettori, che il sistema nervoso centrale è in grado di raccogliere e integrare, permette al soggetto di avere consapevolezza della posizione delle proprie parti del corpo rispetto allo spazio, dei movimenti che gli arti stanno compiendo e della posizione del corpo in relazione agli oggetti dell’ambiente circostante. Tuttavia le percezioni che dalla periferia raggiungono il centro, non forniscono solo informazioni inerenti alla posizione del proprio corpo rispetto agli oggetti dell’ambiente esterno, ma anche la posizione degli arti rispetto a noi stessi. Come già abbiamo detto, il sistema a circuito chiuso si basa sul feedback relativo al movimento che si sta eseguendo e per ottenere tale feedback è necessario l’intervento dei recettori propriocettivi ed esterocettrici di cui l’organismo dispone. Ma se la funzione del feedback è quella di correggere il movimento durante la sua esecuzione, ne deriva che questo tipo di controllo è utile solo per quelle attività motorie che richiedono un tempo di esecuzione relativamente lungo o, comunque, sufficientemente lungo da permettere al feedback di raggiungere il sistema nervoso centrale prima che il movimento sia portato a termine. Un buon esempio può essere la guida dell’auto. Dall’analisi di questa costatazione si evince che per tutti i movimenti veloci la modalità di controllo a circuito chiuso si dimostra inadeguata perché il feedback non sembra in grado di trasmettere l’informazione al sistema centrale prima che l’azione sia completata. Nonostante questa posizione sia accettata dalla maggior parte degli autori è importante sottolineare che non può essere accettata in maniera univoca, in quanto ci sono stati una serie di esperimenti e studi che hanno dimostrato che questo sistema di controllo a circuito chiuso risulta efficiente anche in alcune azioni motorie veloci. LA TEORIA DI ADAMS Una delle teorie più importanti, legata al modello al circuito chiuso, è quella di Adams la cui posizione è di particolare interesse perché è quella che più spesso è stata presa come termine di paragone da posizioni discordanti o alternative ed è anche quella maggiormente supportata da evidenze empiriche. La teoria a circuito chiuso di Adams nasce come risposta alle posizioni dei comportamentisti, i quali riducevano il comportamento ad un rapporto di causa-effetto tra lo stimolo e la risposta. La teoria di Adams – invece- tiene conto delle elaborazioni che hanno luogo tra il momento della presentazione dello stimolo e l’emissione della risposta e considera il soggetto che deve apprendere un dato movimento come una parte attiva e non come parte passiva, in grado di rilevare e correggere l’errore. La teoria (che si basa sulla rilevazione e sulla correzione dell’errore) prevede la presenza di un meccanismo che indica la correttezza del movimento che si vuole effettuare; in secondo luogo è necessaria l’informazione relativa ai risultati del movimento che si sta eseguendo, in modo che questa possa essere confrontata con il movimento che si vuole eseguire. Il meccanismo che indica il movimento corretto viene definito, da Adams, traccia percettiva la quale rappresenta un’immagine del movimento che viene messa continuamente a confronto con i feedback provenienti dalla periferia durante l’esecuzione dell’atto motorio. Se la traccia percettiva e il feedback coincidono, non ci sono errori; se – invece- questi due elementi non coincidono il sistema corregge il movimento fino a farlo coincidere con la traccia percettiva. Da ciò è possibile dedurre che la traccia percettiva è un magazzino in cui specificare le conseguenze sensoriali che il movimento, per essere corretto, deve aver ottenuto. Un altro concetto di fondamentale importanza nella teoria di Adams è quello della traccia mnestica, in cui è contenuta la rappresentazione del movimento che si vuole eseguire. Inoltre il suo livello di dettaglio migliora con la pratica (ossia con la ripetizione del movimento) perché ad ogni ripetizione le conseguenze sensoriali vengono implementate e così, attraverso la traccia percettiva, la traccia mnestica si arricchisce di dettagli che permettono al movimento in essa contenuto di essere rappresentato ad un livello migliore. Il ruolo della traccia mnestica, nella teoria di Adams, è quello di selezionare e iniziare la risposta motoria. In sintesi, si può affermare che il sistema di controllo a circuito chiuso si basa su due elementi: 1) la presenza di un sistema periferico in grado di rilevare l’esecuzione del movimento e trasmettere i parametri a livello centrale; 2) un tipo di rappresentazione interna del movimento in modo da confrontare le informazioni provenienti dalla periferia con quelle presenti nel programma motorio e correggere eventuali errori. Infine si può dire che il controllo effettuato mediante il modello a circuito chiuso è un controllo autoregolante e che i parametri relativi alla prestazione desiderata, anche se sono determinati a livello centrale, sono continuamente sottoposti ad aggiustamenti e modificazioni in conseguenza del feedback. Pertanto il feedback è un elemento fondamentale in questo tipo di sistema di controllo e, per questo motivo, molti studi sono stati condotti sull’importanza di esso nel controllo del movimento. I paradigmi sperimentali più spesso impiegati sono : 1) la misura delle prestazioni motorie in assenza di feedback; 2) lo studio dell’apprendimento motorio in condizioni in cui il feedback era manipolato artificialmente. Una serie di esperimenti all’interno dei quali il feedback non era chiaro hanno messo in evidenza che se il compito di questo è quello di mandare al sistema nervoso centrale delle informazioni relative all’esecuzione del movimento, ne consegue che la conoscenza di dei risultati parziali è un altro presupposto fondamentale della teoria del controllo a circuito chiuso. Per confermare questo aspetto sono stati condotti una serie di studi (soprattutto da Newell), i quali hanno riconfermato (empiricamente) che la conoscenza dei risultati è un presupposto fondamentale. Tuttavia Adams prevede alcune eccezioni per questo principio, affermando che la conoscenza dei risultati è necessaria nei primi stadi di apprendimento motorio, mentre può essere trascurata negli ultimi stadi. Ancora, per confermare questo dato, Newell ha fatto un altro esperimento incui 6 gruppi di persone erano chiamati a svolgere 77 prove. Ai primi 5 gruppi vennero fatti conoscere i risultati solo di alcune prove; al sesto gruppo i risultati di tutte e 77 le prove. L’esperimento mise in luce che: quando la conoscenza del risultato viene negata al soggetto nelle prime fasi dell’apprendimento, la prestazione peggiora; quando invece il soggetto viene privato della conoscenza dei risultati in una fase avanzata dell’apprendimento, la prestazione non subisce nessun calo. In accordo alla teoria di Adams, questi risultati sono stati interpretati come l’effetto di un progressivo consolidamento della traccia percettiva, il quale rende influente l’informazione relativa all’andamento dei risultati. Questo aspetto è strettamente legato all’apprendimento motorio, il quale rappresenta uno dei punti cruciali della teoria di Adams. Infatti l’esperimento dimostra che nel momento in cui un movimento viene ripetuto numerose volte, si verificherà una coincidenza tra i feedback relativi ai livelli parziali della prestazione e la traccia percettiva (ossia i feedback teorici del movimento corretto). Dato che il consolidamento della traccia percettiva si riflette in una traccia mnestica sempre più dettagliata, il movimento rappresentato nella traccia mnestica sarà sempre più simile a quello desiderato, sino a raggiungere un livello di correttezza tale da non essere corretto dai feedback periferici. Di conseguenza le informazioni riguardo al risultato sono trascurabile e solo in questo caso si può parlare di apprendimento motorio. Tuttavia Adams ritiene che questo aspetto non va mai interpretato in maniera rigida, in quanto anche un movimento appreso può essere esercitato in modo errato e non raggiungere lo scopo previsto. Perciò è più giusto affermare che il feedback svolge sempre una funzione di controllo, che –però-entra in gioco solo quando la differenza tra l’informazione proveniente dalla periferia (riferita al movimento in esecuzione) e la traccia percettiva supera una certa soglia. CONTROLLO MOTORIO A CIRCUITO APERTO Oltre al controllo motorio a circuito chiuso c’è anche il controllo motorio a circuito aperto, il quale si differenzia dal primo perché non si basa sul feedback, ma in esso il movimento viene effettuato o in assenza totale di feedback o in presenza di un feedback che, però, non svolge nessuna funzione di controllo durante l’esecuzione dell’atto motorio. Un primo modello di controllo motorio a circuito aperto in cui il feedback è presente ma non svolge nessuna funzione correttiva è quello ipotizzato da James, il quale è stato definito “ipotesi delle risposte a catena”. L’idea di James può essere riassunta in 3 punti: - quando si effettua un movimento (in cui inizio è provocato da uno stimolo interno o esterno all’organismo) questo si accompagna alla contrazione di un muscolo o di un gruppo di muscoli; - la contrazione dà luogo ad un’informazione sensoriale che i recettori muscolari trasformano in un segnale (feedback), il quale funge da stimolo per un nuovo movimento; - la contrazione necessaria a produrre il nuovo movimento dà luogo ad un nuovo feedback che fa scattare il movimento successivo e così via. Da ciò si evince che l’organizzazione del movimento in base alle risposte a catena prevede che il feedback sia in grado di dare il via al successivo movimento, sino a che non si giunge alla completa esecuzione dell’azione; ed è per questo che l’ipotesi delle risposte a catena può essere un’interessante spiegazione per quelle azioni costruite da concatenazioni di singoli movimenti (come allacciarsi le scarpe). In questo modo appare chiaro che mentre nel modello di controllo a circuito chiuso il sistema esecutivo prende atto dall’eventuale errore, mettendo a confronto il movimento che si sta effettuando e quello richiesto e correggendo la prestazione in atto in caso di errore; nell’ipotesi delle risposte a catena il feedback esercita una funzione diversa: esso non viene impiegato come correttore del movimento in atto, ma come avviatore del movimento successivo. MODALITA’ DI CONTROLLO IN RELAZIONE AL TIPO DI MOVIMENTO Una modalità di controllo ancora più probabile è quella secondo la quale l’intero movimento sia completamente rappresentato e programmato ai livelli più alti del sistema nervoso centrale e, dopo essere stato attivato, sia in grado di arrivare a compimento senza l’intervento di nessun tipo di feedback. In questo caso la funzione del feedback sarebbe quella di correggere eventuali errori del movimento successivo; mentre il vero perno di tutto il sistema è rappresentato dal programma motorio, il quale può essere compreso dopo aver fatto un confronto tra sistema di controllo a circuito chiuso e quello a circuito aperto. Una prima distinzione che si fa tra i due sistemi di controllo è che quello a circuito chiuso sembra essere la spiegazione più idonea per i movimenti più complessi e di lunga durata; mentre quello a circuito aperto sembra essere la spiegazione più idonea per i movimenti veloci. Tale differenza, che fa riferimento alla velocità di esecuzione del movimento, si basa sul tempo necessario alla trasmissione del feedback. Se, per esempio, si prende in considerazione un movimento veloce, come l’estensione del gomito, l’esame elettromiografico rileverà una serie alternata di attivazioni-inibizioni del muscolo agonista e di quello antagonista sino alla completa esecuzione del movimento. In questa condizione è naturale chiedersi in che modo il sistema motorio può sincronizzare l’attivazione e l’inibizione dei muscoli agonisti e antagonisti in assenza di feedback. Un concetto legato a questo problema è quello dei gradi di libertà: a tal proposito è importante citare Bernstein, il quale teorizzò che se per ogni movimento possibile i livelli più alti del sistema nervoso dovessero specificare sia i tempi di attivazione-inibizione dei muscoli agonisti e antagonisti, sia la loro corretta coordinazione, si verificherebbe un forte allungamento dei tempi di risposta. A questo punto Bernstein (con altri due studiosi) ha ipotizzato la presenza di una struttura di coordinamento mediante la quale i vari gradi di libertà verrebbero condensati dal sistema motorio. Ciò comporterebbe che i vari muscoli che intervengono nel movimento di un’articolazione e le diverse articolazioni che intervengono in un movimento più complesso verrebbero raggruppate e trattate come una singola unità e, di conseguenza, la funzione svolta dalla struttura di coordinamento è quella di coordinare le varie articolazioni coinvolte in un movimento in modo che la variazione di un’articolazione può essere bilanciata da una variazione di senso opposto di altre articolazioni e dar vita ad un movimento che – in questo modo- viene controllato nella sua completezza. FFEDBACK O PROGRAMMA MOTORIO? Nel sistema di controllo a circuito chiuso, l’elemento fondamentale è rappresentato dal feedback; nel sistema di controllo a circuito aperto, l’elemento fondamentale è rappresentato dal programma motorio, il quale rappresenta un’alternativa all’ipotesi del feedback solo per quei processi che avvengono durante l’esecuzione del movimento in quanto l’importanza del feedback, relativa alla modifica e alla correzione del movimento dopo la sua esecuzione, non viene mai messa in discussione. Lo schema fondamentale del sistema di funzionamento del movimento mediante il programma motorio è quello ideato da Keele e da Summers, il quale mette in luce che: nel momento in cui l’organismo viene stimolato da un input interno o esterno, ciò provoca il bisogno di eseguire un certo movimento; tale volontà di esecuzione si traduce nell’attivazione delle aree cerebrali (dove hanno sede i programmi motori) e attraverso la corteccia motoria e le vie motorie efferenti, il comando di movimento viene inviato ai muscoli. Il movimento provocato dai muscoli provoca vari segnali di feedback (che possono essere propriocettivi o esterocettivi), i quali nel momento in cui arrivano in un centro di confronto, vengono comparati con i feedback previsti dal programma motorio; se i feedback non coincidono con quelli attesi, ciò significa che il movimento non è stato eseguito correttamente ed è per questo che bisogna correggere il programma di movimento o selezionarne uno completamente nuovo. Dopo aver analizzato le ipotesi relative ai due sistemi di controllo: quello chiuso e quello aperto, e possibile affermare definitivamente che nel primo la presenza del feedback è necessaria in quanto senza di esso mancherebbe l’esecuzione stessa del movimento. Nel secondo, invece, il feedback viene impiegato solo per la correzione del movimento successivo: pertanto l’esecuzione del movimento si realizza anche in assenza di feedback. CONFERME AL PROGRAMMA MOTORIO: STUDI SUGLI ANIMALI Per confermare le ipotesi legate al sistema di controllo a circuito aperto sono stati condotti una serie di studi (sia sugli animali che sugli uomini) che – se pur in contrasto tra loro- hanno fornito numerose indicazioni a favore dell’ipotesi della presenza di un programma motorio. Gli animali rappresentano un terreno di studio molto favorevole per verificare quanto il feedback influenzi l’esecuzione del movimento, ed è per questo che sono stati condotti diversi esperimenti i quali hanno confermato l’esistenza di un programma motorio. ESPERIMENTO DELLE SCIMMIE: Taumb e Berman hanno reciso le vie sensoriali afferenti ad un arto ad una scimmia, in modo da impedire il normale feedback cinestesico ed osservare i successivi movimenti dell’arto stesso. Generalmente, dopo la lesione, la scimmia non utilizzava l’arto né per muoversi né per svolgere altre attività; ma se le veniva immobilizzato anche l’altro arto, dopo alcuni giorni la scimmia ricominciava ad utilizzare l’arto privo di feedback e continuava ad utilizzarlo anche quando l’altro veniva liberato. Da questo esperimento, ed altri simili, gli autori conclusero che, siccome le scimmie erano in grado di eseguire tutti i movimenti anche in assenza di feedback, era lecito sostenere che il movimento è guidato e controllato da un programma motorio centrale. Tuttavia è importante sottolineare che ciò vale soprattutto per le attività manipolatorie e meno per le attività legate alla deambulazione, in quanto questa viene integrata – per gran parte- a livello spinale. ESPERIMENTO DEL GATTO : Un altro esperimento molto importante è stato quello di Fentress, il quale analizzò il “grooming” del gatto, ossia quel comportamento innato che è costituito da tutti i movimenti di pulizia dell’animale. Lo studioso osservò il gatto durante il grooming e individuò una tipica catena di movimenti che esso eseguiva con la zampa; allora imputò la zampa all’animale (in quanto essa rappresentava l’organo che generava il feedback) e notò che – nonostante tutto- il gatto continuava a compiere quella stessa catena di movimenti anche in assenza di feedback. Dall’analisi di ciò Fentress confermò la centralità del programma motorio. ESPERIMENTO SUGLI UCCELLI: Lo studioso Nottebohm ha studiato l’apprendimento del canto degli uccelli perché oltre ad essere una forma di linguaggio, si basa su una complicata attività motoria sottostante. Questo studioso dimostrò che se si preleva un uccello dal nido pochi giorni dopo la nascita e lo si alleva, nel momento in cui gli organi del canto maturano, il canto dell’uccello allevato in isolamento sarà più rozzo e carente rispetto a quello dei suoi coetanei. Se –però- questo uccello viene messo a contatto con uccelli adulti, il suo canto migliorerà sino a raggiungere livelli normali. Con questo esperimento Nottebohm ha dimostrato che gli uccelli, per poter sviluppare una capacità di canto normale, hanno bisogno di stare con gli uccelli più adulti in da modificare e incrementare la propria prestazione attraverso il confronto. Se poi l’uccello, dopo essere stato esposto al canto dell’adulto - ancor prima di riuscire a cantareviene reso sordo, la sua capacità canora sarà compromessa per sempre. Se – però- l’uccello viene reso sordo dopo aver imparato a cantare, la capacità canora resta inalterata per almeno un anno dalla lesione. Con questo suo esperimento Nottebhom ha messo in evidenza che se il programma motorio non si è formato, la prestazione non può realizzarsi senza le presenza del feedback; se – invece- il programma motorio si è formato la prestazione può andare avanti anche in totale assenza di feedback. Inoltre gli esperimenti di questo studioso hanno messo in luce un altro aspetto rilevante: l’importanza del feedback per la formazione del programma motorio. Infine un importante lavoro, condotto da Taumb e Berman sulle scimmie, ha dimostrato come il controllo del movimento sia centrale. In questo esperimento gli animali sono stati deafferentati x un lato del corpo, in modo da privare l’arto superiore di qualsiasi afferenza o feedback sensoriale. Il risultato è stato che l’animale usava l’altro arto superiore, trascinando passivamente quello lesionato. Allora lo studioso ha immobilizzato anche l’altro arto e, a questo punto, l’animale ha ripreso ad utilizzare entrambe gli arti per svolgere tutte le attività che per lei sono vitali. Questo esperimento ha dimostrato che i feedback sensoriali non sono indispensabili per il movimento, anche se nel caso in cui un solo arto è lesionato l’animale tende ad utilizzare quello che mantiene la ricezione dei feedback; e che il controllo centrale del movimento può essere mantenuto grazie ad una rappresentazione mentale dei pattern d’azione che rende il feedback non necessario. CONFERME AL PROGRAMMA MOTORIO: STUDI SUGLI UOMINI Per quanto riguarda gli studi condotti sugli uomini, un primo lavoro è stato quello di Lashley: ad un paziente (colpito da un arma da fuoco) che aveva subito l’interruzione delle vie sensoriali afferenti agli arti inferiori, Lashley chiese di muovere le gambe fino ad assumere determinate posizioni del ginocchio. Nella prova veniva escluso anche il feedback esterocettivo, ossia il controllo visivo della posizione delle gambe; e i risultati dimostrarono che il paziente era capace di eseguire il movimento in modo corretto, al pari di un soggetto di controllo. La conclusione alla quale si giunse è stata quella che il feedback propriocettivo dell’arto impegnato nel movimento non è necessario per l’esecuzione dell’atto motorio. Un atro autore molto importante è stato Rothwell che, con i suoi colleghi, ha studiato soggetti umani che – colpiti da gravi neuropatie- si trovavano in condizioni simili alle scimmie deafferentate di Taumb e Berman. Questi soggetti non conservano né la percezione della posizione nello spazio, né la sensazione tattile, ma nonostante questo mantengono la capacità di effettuare movimenti complessi. Il risultato di tale esperimento dimostra che i movimenti sono generati a livello centrale , senza l’intervento di feedback; ma è importante sottolineare che l’esperimento non dimostra la totale inutilità del feedback in quanto dimostra che nei movimenti che coinvolgono più articolazioni, la precisione diminuisce in maniera evidente se manca il feedback. Circa 50 anni fa, Henry e Rogers crearono una condizione sperimentale in grado di mettere in luce la presenza o meno del programma motorio. Essi partirono da un presupposto preciso: il tempo di caricamento del programma motorio, e – in riferimento a ciò- ipotizzarono che se un movimento è guidato da uno specifico programma motorio, dei movimenti più complessi sono guidati da programmi motori più complessi i quali dovranno essere selezionati e caricati prima di iniziare il movimento. Da ciò si può dedurre che la selezione e il caricamento di programmi motori più complessi richieda un tempo maggiore e quindi, partendo da questa ipotesi, Henry e Rogers hanno dimostrato che il tempo di reazione ( intervallo che intercorre tra la presenza dello stimolo e l’inizio del movimento) di un movimento aumenta con l’aumentare della complessità di esso. Per confermare questa loro costatazione, i due studiosi hanno fatto un esperimento in cui dei soggetti erano chiamati a svolgere un atto motorio dopo la comparsa di uno stimolo. Nella prima condizione l’atto motorio richiesto era abbastanza semplice, nella seconda un po’ più complesso, nella terza ancora più complesso. Il risultato fu che mano a mano che aumentava la complessità del movimento, aumentava anche il tempo di reazione, mettendo – così- in evidenza che il ritardo era dovuto o all’incremento del tempo di programmazione del movimento richiesto, o all’incremento del tempo necessario al caricamento di un programma. Anche Klapp, Anderson e Berrian hanno condotto un esperimento che, basandosi sulla relazione tra la complessità del programma motorio e del tempo di coordinamento, ha confermato l’ipotesi secondo la quale l’aumento della complessità del movimento e l’aumento del tempo di reazione sono direttamente proporzionali. Un obiezione a questi risultati potrebbe essere quella che mette in luce che la differenza nei tempi non è dovuta ai tempi di caricamento dei diverdsi programmi motori, ma al maggior tempo necessario per la percezione e l’elaborazione di uno stimolo visivo più complesso. Per rispondere a questo tipo di critica, gli autori sottoposero i soggetti ad un secondo compito in cui lo stimolo era rappresentato dalla figura di un oggetto o di un animale e i soggetti dovevano dire il nome di quell’oggetto o di quell’animale. Il risultato di questo esperimento non fece altro che confermare quello del compito precedente ed è per questo che gli autori conclusero che l’attività motoria implicata nella verbalizzazione di parole non è guidata dal feedback, ma da un programma motorio e il tempo di caricamento del programma è funzione della sua complessità. L’ipotesi della presenza di un programma motorio è stata testata anche in un lavoro di Keele e Posner, i quali hanno misurato l’effetto del feedback su movimenti effettuati a diverse velocità. In questo caso il compito dei soggetti era quello di spostare la mano da un punto A ad un punto B in tempi prestabiliti dallo sperimentatore. Dopo un lungo addestramento i soggetti imparavano ad eseguire i movimenti nei tempi richiesti e, una volta che i tempi di movimento furono correttamente appresi, occasionalmente veniva spenta la luce durante l’esecuzione dell’atto motorio in modo da impedire il feedback su di esso. Il risultato di questo esperimento ha messo in evidenza che la probabilità di errori – causati dall’assenza del feedback visivo- era maggiore nei movimenti effettuati in un tempo superiore ai 250 millesimi di secondo; mentre al di sotto di questo tempo, la presenza o l’assenza del feedback non influiva sulla correttezza della risposta. Dall’analisi di questo esperimento e di altri simili si può concludere che: mentre per i movimenti lenti il tipo di controllo esercitato può avvenire mediante un modello a circuito chiuso; per i movimenti veloci è opportuno un modello di controllo a circuito aperto, dotato di un programma motorio preparato in anticipo. STRUTTURA GERARCHICA E CONTENUTO DEL PROGRAMMA MOTORIO Da questi diversi esperimenti (condotti sia sugli animali che sugli uomini) è stato possibile verificare che una gran parte dei movimenti sono guidati dal programma motorio. Ma in relazione a ciò nasce spontanea una domanda: in che modo le singole caratteristiche del movimento sono specificate all’interno del programma motorio? Per rispondere a tale quesito è opportuno definire, dapprima, cosa si intende per programma motorio. Molti autori hanno definito quest’accezione in base alle loro idee, ma – in generale- x programma motorio si intende una rappresentazione astratta della sequenza di un’azione. Inoltre si è ritenuto importante stabilire- su di un livello più specifico- quali informazioni sono contenute all’interno del programma e come esso sia organizzato. A tal proposito, numerosi studi hanno cercato di dimostrare che nel programma motorio non ci sono informazioni relative ai muscoli implicati nel movimento, bensì delle informazioni che si riferiscono a valori di variabili più generali come: la velocità, la forza e l’ampiezza del movimento. Da ciò si può dedurre che i muscoli e gli arti impiegati in un dato movimento vengono specificati solo in uno stadio successivo del programma. Dato che le info relative ai muscoli si trovano su di un livello gerarchicamente basso, ne consegue che programmi motori relativi a muscoli effettori diversi possono condividere tutti i livelli più alti del programma motorio. Per questo motivo gli studi che si sono occupati dell’organizzazione gerarchica del programma motorio hanno affermato che: se esso è rappresentato centralmente e in esso sono contenute le informazioni relative agli aspetti più generali del movimento, allora è possibile pensare che non esistono tanti programmi quanti sono i movimenti possibili, ma movimenti simili fanno capo allo stesso programma motorio, differenziandosi solo x quanto riguarda i muscoli impegnati. Questa ipotesi è stata confermata dall’esperimento di Raibert il quale chiese ad un soggetto di scrivere una frase prima con la mano dx su un foglio; poi con la mano dx su una lavagna; poi con la mano sx su un foglio; poi la penna fu tenuta nella bocca e, infine, con il piede destro. Il risultato fu che la calligrafia, nelle 5 condizioni, era simile e ciò è stato importante perché da un lato ha dimostrato che esiste un programma motorio per la scrittura e- dall’altro- ha confermato l’organizzazione gerarchica del programma. Infatti la straordinaria somiglianza della calligrafia si baserebbe – secondo l’autore- sull’unicità del programma motorio impiegato il quale varierebbe solo nelle zone periferiche. Una seconda ipotesi che si può formulare relativamente all’organizzazione gerarchica del programma motoria è quella secondo la quale un dato movimento è dato e guidato dallo stesso programma motorio, anche nel caso in cui si cambia l’arto effettore. Se ciò è vero, ne consegue che, quando il movimento viene svolto da un arto diverso deve trasferirsi, dal vecchio al nuovo programma, oltre ai comandi relativi ai muscoli anche eventuali modifiche del programma stesso. Uno studioso che ha cercato di sostenere tale ipotesi è stato Rosenbaum, il quale ha dato vita ad un esperimento in cui chiese ad un gruppo di soggetti di girare una manovella x 30 sec; ad un secondo gruppo di spostarla avanti e indietro sempre in 30 sec. La prestazione fu misurata in numero di movimenti svolti nel tempo concesso per la prova, la quale ha provocato nei soggetti un affaticamento (non solo muscolare, ma anche mentale). Nella seconda parte dell’esperimento ad un gruppo è stato chiesto di ripetere il movimento precedente; ad un altro gruppo è stato chiesto di eseguire un movimento diverso e il risultato ha messo in evidenza che la prestazione peggiore si è registrata quando il movimento da effettuare nella seconda parte dell’esperimento era analogo a quello svolto nella prima parte. Secondo Rosenbaum, il fatto che nella condizione in cui deve essere ripetuto il movimento si ottiene una prestazione peggiore rispetto a quando i due movimenti differiscono è una indiretta conferma dell’esistenza del programma motorio e della sua organizzazione gerarchica. Inoltre se il trasferimento di uno stesso movimento da un arto all’altro richiede un costo più elevato rispetto al trasferimento di movimenti diversi, ciò significa che nel primo caso è lo stesso programma che deve essere impiegato nelle 2 condizioni e quindi la stanchezza accumulata nella prima parte viene trasferita nella seconda condizione sperimentale. Benché la conclusione alla quale è giunto Rosenbaum rappresenta un passo avanti verso la comprensione dell’organizzazione gerarchica del programma motorio, non si può dire che mancano delle parti oscure relative – in particolar modo- al concetto di affaticamento. Innanzitutto bisogna chiarire che il termine affaticamento non fa riferimento solo all’aspetto muscolare del movimento, ma anche alla sua rappresentazione astratta; in secondo luogo sarebbe opportuno spiegare come l’affaticamento viene rappresentato all’interno del programma motorio e come l’esecuzione del movimento risentirebbe di questa rappresentazione. Tutti gli esperimenti fin qui citati hanno messo in evidenza che nel momento in cui viene attuato un controllo del movimento a circuito aperto, l’esecuzione del movimento avviene anche in assenza di feedback. Tuttavia, per eseguire un movimento corretto in assenza di feedback occorre che l’organismo si crei una rappresentazione mentale del movimento in cui sono rappresentati due parametri particolarmente: 1) la distanza; 2) la posizione finale. A tal proposito è importante prendere in considerazione l’esperimento di Bizzi e colleghi, durante il quale ai soggetti veniva mostrata una luce – che dopo si spegneva- e, dopo, gli veniva chiesto di raggiungere la posizione in cui era stata rappresentata la luce. Le modalità attraverso le quali i soggetti giungevano a compiere il movimento correttamente (dopo diverse prove) coincidevano con la costruzione di una rappresentazione mentale o del movimento desiderato basato sulla distanza, o della posizione finale basata sulla posizione spaziale di arrivo del movimento, che è quella più probabile secondo gli sperimentatori. Un altro studio è stato effettuato su delle scimmie deafferentate le quali sono state sottoposte ad un compito di “pointing” (indicazione) durante il quale hanno applicato all’arto che non riceveva il feedback una forza contraria , in modo da mantenerlo – per alcuni secondi- nella posizione iniziale. Il dubbio era se l’animale riusciva a portare l’arto nella posizione corretta; il risultato fu favorevole e con ciò è stato dimostrato che la rappresentazione dei comandi d’azione che sottostanno ad un movimento può essere visto come il passaggio dalla posizione di partenza alla posizione di arrivo del movimento. CAPITOLO V APPRENDIMENTO MOTORIO Molto spesso si fa confusione tra controllo motorio, sviluppo motorio e apprendimento motorio in quanto i confini di queste aree sono molto sfumati e le competenze di ognuno dei 3 settori si sovrappongono agli altri due. Per controllo motorio si intende lo studio dei substrati neurofisiologici del sistema motorio; per sviluppo motorio si intende lo studio delle abilità motorie del bambino (con particolare riferimento allo sviluppo filogenetico - o evolutivo- dei movimenti); per apprendimento motorio si intende lo studio delle modalità di acquisizione di nuove abilità da parte di soggetti adulti. STADI DI APPRENDIMENTO DELLE ABILITA’ MOTORIE Prima di capire cosa si vuole intendere parlando di apprendimento motorio, è importante chiarire – prima- cosa si intende per abilità motoria. A tal proposito si può fare riferimento alla definizione di Bennet, il quale ha affermato che ci si trova in presenza di un’abilità motoria quando il movimento raggiunge un livello elevato nelle caratteristiche di economicità (ossia minor sforzo) e velocità. Molti anni dopo tale definizione è stata confermata empiricamente da una serie di studiosi: Kamon e Gormley hanno messo in evidenza come durante l’esecuzione di un esercizio ginnico la durata dell’attivazione di vari muscoli di ginnasti esperti risultava inferiore a quella degli stessi muscoli di soggetti principianti. Per quanto riguarda la velocità, importante è stato il lavoro di Crossman durante il quale fu messa in relazione la velocità di esecuzione di un compito motorio con la durata del periodo di pratica effettuata per quel compito. Con un esperimento – condotto in una fabbrica di sigari- Crossman ha dimostrato che il miglioramento della prestazione può continuare anche dopo periodi di pratica estremamente prolungati. Un altro grande studioso è stato Fitts, il quale ha affermato che è possibile distinguere 3 fasi dell’apprendimento motorio: la fase cognitiva, la fase associativa e la fase di automazione. Nella fase cognitiva vengono prese le decisioni che permettono i primi tentativi di esecuzione del movimento. Tali decisioni sono facilitate dalla verbalizzazione del movimento che si sta compiendo e dalle strategie adottate per compierlo. Nella fase associativa i vari movimenti che compongono la prestazione vengono condensati in un’unica azione e nell’ultima fase ha luogo l’automazione dei processi cognitivi sottostanti l’attività motoria in modo che il sistema di controllo operi in modo molto ridotto. L’automazione dei processi raggiunta nella terza fase è testimoniata anche dalla difficoltà dei soggetti a verbalizzare le operazioni che vengono svolte durante la prestazione. Ciò accade perché le risorse attentive hanno una capacità limitata, pertanto nel momento in cui un movimento viene automatizzato tali risorse vengono impiegate in attività diverse, che si svolgono parallelamente a quella automatizzata. FASE COGNITIVA Nella fase cognitiva viene messo in atto il modo più diretto per insegnare un’attività motoria a chi ne è completamente all’oscuro; tale modo è quello di dare una dimostrazione pratica dell’attività stessa, ossia mimare il movimento. Infatti, nelle prime fasi di apprendimento motorio è molto difficile insegnare il movimento attraverso le parole, ed è per questo che i movimenti vengono mostrati e mimati, in modo da dare al soggetto di percepirli e memorizzarli. In questo caso si parla di apprendimento per imitazione. Per quanto riguarda la percezione del movimento non ci sono particolari problemi; per quanto riguarda la memorizzazione del movimento è importante sottolineare che x memorizzare l’attività motoria che deve essere appresa, il soggetto deve individuare e sottolineare le caratteristiche cruciali del movimento osservato. Quest’ultimo aspetto è legato al livello di conoscenza motoria posseduto da chi deve apprendere, il quale deve essere preso in considerazione da chi imita, in modo da modulare la rappresentazione secondo diversi gradi di specificità. Tuttavia, mimare il gesto che deve essere appreso non è l’unico modo per trasferire un’abilità motoria ad un soggetto, il quale nelle prime fasi di apprendimento è costretto ad impiegare molte risorse attentive per imparare un’azione motoria, i cui diversi movimenti devono essere isolati e memorizzati singolarmente. Newell ha affermato che i compiti motori che hanno nel suono la loro caratteristica principale possono essere appresi più velocemente se chi li deve apprendere viene sottoposto all’ascolto del suono prodotto dal movimento numerose volte. Un’ulteriore modalità di apprendimento è quella del movimento guidato: il braccio, la gamba o il corpo di chi deve apprendere è guidato passivamente dall’insegnante durante l’esecuzione del movimento. Tale tecnica viene impiegata nel campo dello sport (come il tennis), ma nonostante tutto fornisce risultati modesti, forse a causa della passività a cui il soggetto è costretto. Infatti durante l’esecuzione autonoma il soggetto compie una serie di errori che funzionano da feedback per il miglioramento della prestazione; mentre durante l’esecuzione del movimento guidata il soggetto non compie errori e, di conseguenza, vengono a mancare quei feedback che possono condurre ad un miglioramento della prestazione FASE ASSOCIATIVA La fase associativa è uno dei momenti più importanti dell’apprendimento motorio in quanto è proprio in questo momento che avviene la fusione di ogni singolo movimento in un unico insieme che costituisce l’abilità motoria. Il principio della fusione delle singole parti costituenti l’attività motoria è in relazione con il problema del trasferimento delle capacità apprese da un movimento ad un altro. Per poter condensare e rendere più veloci due o più movimenti bisognerebbe trasferire le variabili (già apprese) del primo movimento al secondo; tuttavia il trasferimento di tali variabili deve fare i conti con l’organizzazione gerarchica del controllo motorio, secondo la quale al livello più basso ci sono le variabili specifiche (come i muscoli implicati nel movimento), al livello più alto c’è una rappresentazione del movimento che prescinde dallo specifico arto o dai particolari gruppi di muscoli impiegati nel movimento. Questi livelli si differenziano ulteriormente e nettamente per quanto riguarda la possibilità di trasferimento delle abilità motorie apprese: infatti mentre il trasferimento dei parametri contenuti ai livelli più bassi è nullo, la più generale rappresentazione dell’azione contenuta ai livelli più alti può essere trasferita tranquillamente ad altre azioni simili. Un esempio che può chiarire questo punto è quello della guida dell’automobile: se chiediamo ad un soggetto di frenare con il piede sinistro, egli effettuerà un movimento corretto in alcune parti (ossia nell’azione di pressione) e molto approssimativo e impreciso in altri (la frenata sarà o troppo immediata o troppo ritardata). Da ciò si deduce che ad un livello più alto e più generale è rappresentata la finalità dell’azione, che è centralizzata e comune ad entrambe gli arti; su ad un livello più basso e più specifico non è possibile trasferire un’azione da un arto all’altro. Pertanto si può concludere che la fase associativa è contraddistinta da un processo di compattamento delle attività motorie. Tale compattamento avviene mediante il trasferimento di abilità contenute in movimenti già appresi ad altri movimenti che non sono stati appresi e appartengono a classi generali di azioni comuni. FASE DI AUTOMATIZZAZIONE Il problema dell’apprendimento di abilità motorie può essere messo in relazione alla categorizzazione di Poulton, il quale distinse i movimenti in aperti e chiusi. I movimenti chiusi sono quelli messi in atto in un ambiente stabile, ossia in un ambiente dove le variabili sono costanti e prevedibili; i movimenti aperti sono quelli messi in atto in un ambiente che presenta dei fattori che possono cambiare continuamente e che quindi non sono prevedibili. Anche nell’ambito dell’apprendimento motorio i movimenti seguono tale distinzione: infatti un movimento si definisce aperto nel momento in cui durante la sua esecuzione il soggetto non è in grado di predirne l’andamento e il valore delle variabili presenti; mentre un movimento si definisce chiuso quando l’autore del movimento è capace di effettuarlo (dopo un periodo di pratica) in un ambiente per lui prevedibile e controllabile. Il passaggio dal movimento aperto a quello chiuso rappresenta il processo di automazione del movimento, ossia la possibilità di effettuarlo in maniera automatica concentrando le risorse attentive su altre attività. Una dimostrazione del passaggio dal movimento aperto al movimento chiuso coincide con l’esperimento di Lang in cui c’erano due gruppi di soggetti posti davanti allo schermo di un Pc il cui compito era quello di muovere una leva che attivava un cursore nello schermo. Il compito consisteva nel raggiungere con il cursore uno stimolo visivo che si spostava o in direzione orizzontale o in direzione verticale. Per un gruppo la traettoria seguita dallo stimolo variava in maniera casuale e, quindi, nulla era prevedibile; per l’altro gruppo la variazione avveniva solo lungo la dimensione orizzontale, attorno la quale lo stimolo seguiva una traiettoria ciclica che si ripeteva ogni due secondi. Il risultato di quest’esperimento dimostrò che il primo gruppo, nonostante le numerose prove, non raggiunse nessun miglioramento nella prestazione; mentre la prestazione del secondo gruppo migliorò in relazione alla dimensione orizzontale, in quanto questo dato era prevedibile; mentre per la direzione verticale non ci fu nessun miglioramento in entrambe i gruppi. Tuttavia i due gruppi si differenziarono anche per la dimensione verticale perché - anche se nessuno dei due gruppi presentò un miglioramento nella prestazione legata a questa traiettoria- il secondo gruppo riuscì a raggiungere dei risultati migliori rispetto al primo anche sotto questo punto di vista. Ciò si è verificato perché automatizzando il movimento legato alla traiettorie orizzontale, tutti i soggetti hanno impiegato la maggior parte delle loro risorse attentive sul movimento legato alla traiettoria verticale., con un conseguente miglioramento delle risposte anche sotto questo punto di vista. TEORIE DELL’APPRENDIMENTO MOTORIO L’apprendimento motorio – nella sua assunzione più generale- può essere considerato la conseguenza dell’acquisizione di una più corretta rappresentazione dell’azione, che può riferirsi sia allo specifico movimento appreso sia ad una più generale classe di azioni a cui il movimento appartiene. Nel primo caso il rapporto tra rappresentazione ed esecuzione dell’azione è di “uno ad uno” e ad esso si lega la teoria di Adams; nel secondo caso tale rapporto è di “uno a molti” e ad esso – invece- si lega la teoria di Schmidt. TEORIA DI ADAMS La teoria dell’apprendimento motorio a circuito chiuso di Adams prevede che l’atto motorio si basi su due unità: la traccia mnestica, che seleziona e da avvio ad un movimento in seguito ad un atto di volontà; la traccia percettiva, la quale controlla e modifica il movimento confrontando quello voluto con quello effettuato. La traccia percettiva si forma mediante i feedback interni ed esterni del movimento effettuato ed è x questo che Adams ritiene che nelle prime fasi dell’apprendimento motorio la percezione del movimento rappresenta un feedback privilegiato per la costruzione della traccia percettiva, ma con l’aumentare della pratica la traccia percettiva diventa un’immagine motoria che si imprime nella nostra memoria. L’ipotesi dell’esistenza di due strutture diverse nell’azione motoria è giustificata da 3 ragioni teoriche: 1) prima di tutto la struttura che seleziona e da inizio al movimento non può essere la stessa che controlla e confronta il movimento in atto, perché se così fosse si otterrebbe il paradosso di un movimento che si confronta con se stesso. 2) in secondo luogo la traccia percettiva si basa sui feedback provenienti dal movimento, i quali hanno luogo durante o dopo l’esecuzione del movimento ed è per questo che essa non può essere responsabile dell’esecuzione del movimento stesso. 3) in terzo luogo le due strutture sono responsabili di due diversi processi: la traccia mnestica è deputata al ricordo-richiamo del movimento attraverso il quale permette di iniziare il movimento; la traccia percettiva è deputata al riconoscimento del movimento in atto, dando luogo ad un processo di riconoscimento e valutando la correttezza rispetto al movimento voluto. Secondo Adams, l’apprendimento motorio avviene mediante due stadi successivi: lo stadio verbalemotorio e lo stadio motorio. Lo stadio verbale-motorio avviene nelle prime fasi dell’apprendimento ed è caratterizzato da un controllo verbale cosciente e da un numero elevato di errori. A questo stadio di apprendimento il soggetto non ha avuto un’esperienza sufficiente per verificare le conseguenze del proprio movimento ed è per questo che deve basarsi sulla percezione dei risultati della propria azione per decidere se questa è stata effettuata in modo corretto o meno. Con l’aumento della pratica la conoscenza del risultato è sempre meno necessaria in quanto la traccia percettiva può formarsi esclusivamente sui feedback di tipo cinestesico. Nello stadio motorio la traccia percettiva si consolida e diviene una forma di rappresentazione interna del movimento; essa è molto consistente e resistente al decadimento. A questo livello l’abilità motoria è già appresa ed è per questo che la pratica comporta solo il miglioramento della prestazione. Un esempio può essere considerato l’imparare a portare la bicicletta. Come qualsiasi grande teoria, anche quella di Adams ha ricevuto numerose critiche, le quali si sono concentrate su 2 punti particolarmente: 1) il rapporto tra la traccia mnestica e i movimenti in essa contenuti; 2) il problema dell’esecuzione di movimenti mai effettuati in precedenza. Per quanto riguarda il primo punto, risulta difficile ammettere l’esistenza di tante tracce mnestiche x quanti sono i movimenti appresi, perché tale organizzazione comporterebbe una grande mole di lavoro sia x l’immagazzinamento di ogni singolo movimento, sia per il suo recupero. Per quanto riguarda il secondo punto la questione diventa più complessa in quanto se la traccia mnestica è necessaria per iniziare un movimento, ed essa si è formata dopo una serie di ripetizioni di quella stessa azione motoria, il processo descritto sembra seguire un criterio circolare di difficile comprensione. A ciò si aggiunge il fatto che 2 movimenti non possono mai essere uguali, pertanto la ripetizione di un gesto non è mai la copia esatta dell’atto motorio precedente e – di conseguenzail secondo è un movimento nuovo. Questo schema basato sul rapporto “uno ad uno” è stato modificato da Schmidt, il quale ha proposto un modello di apprendimento motorio basato su un rapporto di “uno a molti”. TEORIA DI SCHMIDT La teoria di Schmidt viene definita teoria dello schema ed essa è riuscita ad affermarsi in modo così pregnante per 3 motivi fondamentali: 1) innanzitutto perché ha fornito delle risposte a quegli interrogativi che la teoria di Adams aveva lasciato in sospeso; 2) in secondo luogo negli anni ’70 c’è stata una grande enfasi nei confronti delle teorie del controllo e dell’apprendimento motorio a circuito aperto; 3) infine perché tale teoria fece un passo avanti sia rispetto al modello a circuito chiuso, sia rispetto alle prime ipotesi sulla struttura del programma motorio appartenente al modello di controllo motorio a circuito aperto. Lo schema può essere descritto come una regola generale che rappresenta tutte le relazioni esistenti tra le diverse variabili oggetto del movimento; e la più grande differenza che c’è tra la teoria dello schema e quelle ad essa precedenti è che tale teoria è riuscita a rispondere a due problemi rimasti irrisolti: quello dell’immagazzinamento e quello della novità (ossia dell’esecuzione di un nuovo movimento). Un ipotesi che potrebbe fornire una spiegazione relativa all’immagazzinamento di una grande quantità di movimenti è quella che sostiene la presenza di un processo generativo che sia in grado di formare una regola che può essere applicata a tutti i movimenti simili e appartenenti ad una stessa categoria. Pertanto lo schema sarebbe formato da un insieme di principi che fungono da istruzione x la produzione di una serie di movimento. Secondo Schmidt la teoria dello schema si basa su 4 tipi di informazioni motorie che il soggetto immagazzina durante l’esecuzione del movimento: i parametri specifici, ossia i valori delle variabili intervenute; il risultato che si è ottenuto a seguito del movimento; le conseguenze sensoriali e le condizioni di partenza. Secondo la teoria, dopo che il movimento è stato completato, le varie informazioni relative ai 4 punti descritti vengono registrate e la forza tra i vari elementi che compongono l’azione motoria aumenta ad ogni ripetizione dello stesso movimento o di movimenti simili. In questo modo si forma lo schema, il quale sarà più completo e articolato quanto maggiore è stata la variabilità nella fase di formazione. Dopo che si è formato lo schema, le abilità motorie in esso contenuto possono essere trasferite a movimenti nuovi, favorendo – in questo modo- il processo di apprendimento. Oltre alle teorie di Adams e Schmid, molto importante è anche il pensiero di Kurt Meinel, il quale individua 3 tappe che delineano il percorso dell’apprendimento motorio: 1) fase della coordinazione grezza, la quale caratterizza il primo approccio all’esecuzione di un’abilità. In questa fase il movimento può avvenire grazie alla comprensione del compito da parte dell’allievo che: raccoglie, attraverso l’apparato senso-percettivo, tutte le info provenienti dall’esterno o dall’interno dell’organismo; si rappresenta mentalmente il movimento e lo esegue sotto il controllo dell’attenzione. Per le prime volte l’esecuzione è lenta, imprecisa, poco efficace ed evidenza una serie di caratteristiche, quali: ritmo esecutivo inadeguato, carenza di timing o tempi di intervento imprecisi o errati, scarsa fluidità e ampiezza, scarsa rapidità e precisione, frequenti tentativi e insuccessi. 2) Fase della coordinazione, la quale evidenzia il momento in cui l’abilità motoria è già a buon livello. Infatti l’esecuzione appare più fluida e facilitata e il movimento si presenta armonico, continuo, economico, efficace, parzialmente automatizzato e con timing precisi e puntuali, inoltre scompaiono i movimenti associati (ossia quei movimenti che il sogg. compie nelle prime fasi dell’apprendimento motorio), si raggiunge un buon grado di automatizzazione, ma la variazione della situazione o la comparsa di fattori di disturbo fanno ricomparire errori grossolani. 3) Fase della coordinazione fine. In questa fase il soggetto fa un ulteriore passo avanti per il completo controllo del movimento e giunge alla completa e definitiva acquisizione di abilità e competenze motorie. Questo stadio viene definito abilità di alto livello o disponibilità variabile e il movimento si presenta automatizzato ed eseguito con facilità e disinvoltura; adattabile a situazioni nuove; eseguibile anche in presenza di fattori di disturbo. IMAGERY MOTORIA E SIMULAZIONE Anche se queste teorie tradizionali hanno una grossa rilevanza, resta il fatto che alcuni elementi restano poco sviluppati. In particolare, negli ultimi anni, per spiegare l’apprendimento motorio sono stati sottolineati: l’importanza della “imagery motoria” e il ruolo del sistema di neuroni a specchio. Per imagery motoria si intende uno stato dinamico in cui le rappresentazioni di un atto motorio sono richiamate interamente nella memoria di lavoro, senza che questo porti ad un output motorio. Pertanto la differenza che c’è tra l’immagine mentale e l’imagery motoria è che nel primo caso ci si forma la rappresentazione mentale di un oggetto, di un individuo; nel secondo caso ci si forma la rappresentazione mentale dell’esecuzione di un movimento. Inoltre l’imagery motoria non riguarda solo il mondo che ci circonda, ma anche il sé e mentre prima si pensava che essa fa riferimento da una rappresentazione consapevole ed esplicita, oggi si è costatato che essa comprende anche aspetti impliciti e non consapevoli. Per dimostrare che esiste una differenza tra immagini mentali e immagini mentali motorie sono stati fatti una serie di esperimenti, tra i quali ricordiamo quello di Jeannerod e Franck che ha messo in evidenza l’esistenza di un processo di simulazione, ossia di una forma di riproduzione interna e inconsapevole del movimento che i soggetti si creano. Oltre a quest’esperimento ce ne sono stati molti altri in cui veniva chiesto ai soggetti di immaginare di eseguire un movimento. I risultati di questi esperimenti hanno messo in luce che il tempo impiegato per le azioni simulate è pari a quello impiegato per le azioni realmente eseguite; inoltre le azioni più sono complesse, più tempo richiedono. Le basi neurali dei processi di imagery motoria vanno ricercate nel sistema di neuroni a specchio, che nelle scimmie scaricano anche quando il compito non richiede esplicitamente di compiere un’azione. Alcuni studi hanno dimostrato che la visione o la denominazione di alcuni oggetti afferrabili attivano la corteccia premotoria anche quando non deve essere eseguita nessuna azione e in letteratura la questione relativa alla misura in cui la simulazione attivi la corteccia motoria primaria è molto dibattuta in quanto degli esami fatti con la PET hanno dimostrato che la corteccia motoria primaria non si attiva in modo significativo durante l’immaginazione; mentre degli studi condotti con la risonanza magnetica funzionale e con la TMS hanno indicato il contrario, anche se il livello di attivazione corticale è più basso nei processi immaginativi rispetto a quello che si ha durante l’effettiva esecuzione del movimento. Per quanto riguarda le implicazioni che l’imagery motoria e i neuroni mirror hanno sull’apprendimento motorio vediamo che la prima – insieme all’osservazione- rappresenta uno strumento utile a migliorare il funzionamento del sistema motorio; il convolgimento del sistema dei neuroni specchio è supportato da prove le quali dimostrano che l’osservazione porta alla formazione di memorie simili a quelle che sottostanno ai cambiamenti, all’interno della rappresentazione motoria, indotti dal movimento stesso. In uno studio recente con la TMS è stato dimostrato che la corteccia motoria primaria: - mette in luce attività legate al sistema dei neuroni specchio in risposta all’osservazione di un movimento; - è coinvolta nell’apprendimento motorio; - influenza la formazione di memorie motorie, la quale costituisce un passo importante per l’acquisizione di abilità motorie. Infine bisogna sottolineare che una serie di esperimenti hanno dimostrato che sia l’osservare sia l’immaginare di svolgere un movimento comporta dei miglioramenti nelle proprie capacità motorie e- di conseguenza- favoriscono l’apprendimento motorio. I FATTORI DELL’APPRENIDMENTO MOTORIO L’apprendimento motorio è un processo che tende a far crescere i livelli di competenza (intellettiva, motoria, sociale e comunicativa) di un soggetto, soprattutto grazie all’integrazione di 3 componenti: l’allievo, l’insegnante e l’ambiente. L’allievo è la componente primaria e quella più importante in quanto senza di lui le competenze dell’insegnante e la presenza di un ambiente favorevole possono incidere ben poco sull’apprendimento motorio. Una delle caratteristiche più importanti che l’allievo deve possedere è la motivazione ad apprendere, la quale aumenta volontariamente e spontaneamente le capacità di attenzione e di concentrazione sul compito, le situazioni di sopportazione della fatica e del superamento dell’errore. Il livello psicomotorio dell’allievo, ossia il grado di capacità e di abilità motorie presenti nell’allievo, è un fattore determinante x l’apprendimento durante il quale diventano fondamentali le ripetizioni del compito unite alla consapevolezza degli errori commessi ed alla conoscenza degli interventi di correzione. Infine va sottolineato che la ripetizione del movimento, sotto gli occhi attenti di un insegnante, è la metodica più efficace per perfezionarlo prima di automatizzarlo. L’insegnante ha un compito molto importante: quello di far crescere i propri allievi facilitando il loro processo di apprendimento. Per far questo egli deve possedere delle conoscenze culturali non solo sul movimento, ma anche sull’età evolutiva. Inoltre è indispensabile la conoscenza dell’allievo e della situazione di partenza, ossia la ricerca sistematica e oggettiva del suo livello psicomotorio, ed è per questo che è necessario prendere in considerazione le capacità (condizionali, coordinative, intellettive e sociocomunicative); le abilità; gli interessi e le motivazioni. L’ambiente in cui avviene l’apprendimento deve essere attrezzato in modo da facilitare lo sviluppo di abilità motorie e deve essere accogliente per chi apprende in quanto un allievo che si sente accettato e – nel contempo- si sente parte di un gruppo può sbagliare, correggersi e apprendere con maggiore serenità. Inoltre è importante evidenziare che la crescita sociale viene favorita da situazioni motorie di interazione con l’altro, le quali possono portare l’allievo a conoscere e a collaborare con esso, ad avere fiducia e senso di responsabilità nei suoi confronti, capirlo, accettarlo e aiutarlo. METODOLOGIE DELL’APPRENDIMENTO MOTORIO: l’apprendimento motorio può avvenire secondo diverse metodologie: per imitazione, per tentativi ed errori, per analisi e comprensione del compito. Apprendimento per imitazione: questa modalità di apprendimento è caratterizzato dalla spontaneità e della naturalezza e – in più- necessita della presenza di un modello da imitare, il quale può essere l’insegnante, un adulto o un compagno. Essa è una modalità che si adatta all’apprendimento di abilità globali o parziali, semplici, naturali e poco complesse e il risultato che l’allievo ottiene è estremamente individualizzato e dipendente dal suo grado di descrizione globale. Apprendimento per tentativi ed errori: l’aspetto principale di questa metodica di apprendimento è la situazione-problema. Infatti al soggetto viene dato in problema motorio da risolvere mediante una soluzione che per lui risulti appropriata. In questo tempo il tempo di apprendimento è più lungo rispetto al primo perché bisogna dare al soggetto la possibilità di trovare la soluzione più giusta; ma anche se è più lento esso è attivo e coinvolgente in quanto presenta il vantaggio di far ragionare l’allievo sollecitandone l’aspetto intellettivo-intuitivo. Tale metodica – infine- può essere distinta in 4 fasi: fase dell’esplorazione, fase dei tentativi e degli errori, fase della scelta più efficace e automatizzazione della risposta migliore. Analisi e comprensione del compito: in questa metodica di apprendimento l’insegnante è il vero protagonista in quanto è lui a scegliere le strategie di comunicazione più idonee, tra le quali ricordiamo: quella verbale, quella audio-visuale e quella espressivo-motoria. In questo caso all’allievo viene chiesto – prima- di capire o eseguire un movimento e – dopo- di correggerlo sotto l’attenzione vigile ed esperta dell’insegnante. Questa metodica può avvenire in 2 forme: quella analitica, durante la quale l’abilità viene scomposta in parti e l’allievo impara dapprima l’esecuzione corretta delle singole parti e, poi, le ricompone per automatizzare il gesto globale; quella globale in cui le correzioni vengono apportate direttamente sull’esecuzione completa. CAPITOLO VI MOVIMENTO E IMITAZIONE Il concetto di imitazione è stato ben chiarito da Tommasello, il quale ha esplicitato la differenza che c’è tra emulazione e imitazione. Con il termine emulazione si intende il fatto che gli indvidui, grazie all’osservazione di azioni, apprendono le proprietà degli oggetti (ossia dell’ambiente) senzanecessariamente – apprendere l’azione stessa. Con il termine imitazione si indica l’apprendimento delle azione e del modo in cui svolgerle. In senso ancora più stretto l’imitazione può essere intesa come la capacità di compiere azioni apprendendo il metodo necessario al loro svolgimento. Attenendosi a questa seconda definizione, l’imitazione prevede diverse fasi: 19 osservare qualcuno che compie un’azione o un movimento; 2) individuare un piano x agire; 3) implementare e specificare il piano; 4) eseguire l’atto motorio o l’azione. Da ciò si evince che l’imitazione coinvolge la visione, la pianificazione e il controllo motorio e il suo studio può vertere anche su aspetti sociali e comunicativi. IMITAZIONE NON INTENZIONALE-IMPLICITA: I NEURONI MIRROR Il fenomeno dell’imitazione implicita è molto discusso in letteratura; infatti basti pensare all’effetto camaleonte o all’effetto del contagio emozionale, nei quali si tende automaticamente e involontariamente a imitare la mimica facciale, i gesti, la postura e il tono di voce dell’interlocutore. Alcuni studi recenti hanno dimostrato che le basi neurali dell’imitazione motoria risiedono nell’attivazione dei neuroni a specchio (o mirror neurons) i quali sono una classe particolare di visuo-neuroni che sono coinvolti sia nell’imitazione non intenzionale-implicita, sia nella spiegazione di essa. Questi neuroni specchio sono stati trovati nell’area F5 della corteccia motoria dei macachi, accanto ai neuroni canonici, che scaricano alla comparsa di un oggetto. La caratteristica principale dei neuroni mirror è che essi scaricano sia quando l’animale compie un’azione sia quando vede di compiere un movimento da un suo simile o dall’uomo. Tuttavia questi neuroni non scaricano se è presente solo l’effettore, se i movimenti non sono rivolti ad oggetti e nemmeno in presenza del solo oggetto; essi scaricano solo se è visibile un’interazione tra l’effettore e l’oggetto. Inoltre è stato anche scoperto che esistono neuroni specchio che rispondono solo ad azioni identiche a quelle codificate dal punto di vista motorio e azioni che rispondono ad azioni simile ma non identica. Un’altra serie di studi recenti, condotti con macchine come la risonanza magnetica funzionale e la TMS, hanno dimostrato che i neuroni mirror esistono anche nell’uomo e che osservare un’azione porta ad attivare aree corticali che solitamente si attivano nel momento in cui avviene l’esecuzione del movimento. Un altro problema che molti studiosi si sono posti è quello relativo al come noi riusciamo ad imitare un’azione motoria in modo non-intenzionale; e su tale questioni sono nate diverse ipotesi. La prima prevede che l’imitazione non-intenzionale implicita facilita lo sviluppo di forme di imitazione esplicita, rendendo più facile la messa in atto di un comportamento imitativo il quale è importante per aumentare la coesione tra i gruppi e l’empatia tra i soggetti. Una seconda ipotesi lega l’imitazione implicita alla capacità di comprendere le azioni altrui. Di questa ipotesi esistono due versioni: una più moderata e una più forte. Secondo la formula più moderata, l’imitazione non intenzionale aiuterebbe a comprendere le azioni altrui attraverso un processo di simulazione mentale interna, il quale implica che un meccanismo cognitivo può essere utilizzato anche off-line, ricevendo informazioni dalla memoria piuttosto che dagli stimoli ambientali. Inoltre la comprensione delle azioni altrui avviene – secondo tale ipotesi- quando l’osservazione di azioni attiva un meccanismo di risonanza nel nostro sistema cognitivo. Secondo la formula più forte l’imitazione non-intenzionale contribuisce a comprendere non solo ciò che gli altri stanno facendo, ma anche le intenzioni e gli obiettivi che stanno dietro le azioni che vediamo; inoltre sembra che tale forma di imitazione possa costituire la base per la formazione di una teoria della mente che consente di comprendere gli stati mentali degli altri. In sintesi, secondo questa formula, il cervello dell’osservatore riproduce interamente – attraverso un processo di simulazione- non solo il pattern motorio ma anche gli effetti che l’azione comporta. Una terza ipotesi connette l’imitazione non intenzionale allo sviluppo del linguaggio in quanto pare che alcuni studi hanno dimostrato che l’area di Broca ( a cui è legata la funzione di comprensione del linguaggio) sia la corrispondente dell’area F5 della scimmia, nella quale si trovano i neuroni specchio. Una quarta ipotesi è stata formulata nel 2005 da Wilson e Knoblich, i quali interrogandosi sul ruolo svolto dall’imitazione non intenzionale hanno dedotto che questo tipo di imitazione svolge la funzione di aiutare a percepire il comportamento dei propri simili, funzionando come un sistema che gli autori definiscono “emulazione percettiva”. Questo sistema registrerebbe in tempo reale il comportamento di altri membri della propria specie e- in questo modo – avrebbe la funzione di colmare lacune e informazioni ambigue, dando la possibilità al soggetto di reagire velocemente agli stimoli. Questa ipotesi si differenzia dalla seconda, e in particolar modo dalla formula più forte di quell’ipotesi, perché non verte sulla comprensione dell’azione, ma sulla percezione di essa. Più in particolare, l’ipotesi di Wilson e di Knoblich si differenzia dalla seconda per due aspetti: 1) l’imitazione non intenzionale viene concepita come una sorta di emulatore, che svolge una funzione predittiva, ossia aiuta il processo percettivo mentre si svolge e ne anticipa gli effetti; 2) essa richiede una rappresentazione del corpo umano che sia indipendente dal punto di vista dell’osservatore. PRINCIPALI SPIEGAZIONI DEL COMPORTAMENTO IMITATIVO: LA TEORIA DEL MATCHING DIRETTO Le teorie che riguardano il comportamento imitativo si distinguono in due grandi classi: 1) le teorie che postulano l’esistenza di un unico processo, le quali comprendono la teoria del matching diretto e la teoria AIM; 2) le teorie duali, ossia quelle che postulano l’esistenza di due processi, come la teoria dell’imitazione basata su scopi. TEORIA DEL MATCHING DIRETTO La teoria del matching diretto afferma che il sistema motorio si attiva – in modo diretto- nel momento in cui il soggetto percepisce l’azione, pertanto la percezione del comportamento altrui costituisce l’input dal quale partire per eseguire l’azione imitativa. Questa teoria è nata dall’idea che esiste un codice comune tra l’azione percepita e quella da eseguire, e cioè tra percezione e azione, e le basi neurali di essa si ritrovano negli studi sui neuroni specchio. Secondo la teoria del matching diretto c’è un circuito neurale specializzato che è in grado di connettere in modo diretto le azioni osservate con i comandi motori, inducendo il soggetto a produrre azioni simili a quelle osservate. Detto in altre parole questa teoria propone l’idea che l’osservazione di azioni attiva un meccanismo di risonanza o di simulazione mentale, che costituisce la base cognitiva dell’imitazione. Un primo esempio di risonanza riguarda proprio i comportamenti imitativi, ossia la tendenza automatica a ripetere i movimenti di altri soggetti. Ciò è visibile soprattutto nei comportamenti imitativi dei neonati umani o di alcune specie di animali. Un secondo meccanismo di risonanza consiste nell’attivazione di neuroni motori durante l’osservazione di azioni motorie compiute da soggetti diversi da sé. Questo meccanismo – molto particolare- non si traduce in forma esplicita di imitazione, ma resta implicito dando –probabilmente- la possibilità all’individuo di capire i comportamenti altrui. Nel 1999 due studiosi hanno confermato la teoria del matching diretto, secondo la quale l’imitazione si basa su un meccanismo che permette all’azione osservata di essere “mappata” da un’azione prodotta internamente. Per verificare questa spiegazione è stato fatto un esperimento in cui i partecipanti venivano sottoposti a condizioni di osservazione, durante le quali i soggetti dovevano semplicemente osservare gli stimoli; e a condizioni di osservazione-esecuzione durante le quali dovevano compiere un’azione motoria in risposta allo stimolo presentato o imitare il movimento. I risultati dello studio, condotto con la risonanza magnetica funzionale, hanno messo in luce che sia la corteccia frontale inferiore sx, sia la regione rostrale del lobo parietale superiore dx si attivano registrando la risposta motoria, indipendentemente da come questo movimento veniva evocato. La cosa interessante è che queste aree si attivano specialmente durante l’imitazione, e cioè quando un soggetto esegue la stessa azione che sta osservando e che viene compiuta da altri. Dall’analisi di tutto ciò è possibile – quindi- affermare che tali risultati depongono a favore della teoria del matching, la quale prevede che le aree neurali in cui ha luogo il matching contengono neuroni che scaricano in risposta a delle azioni e neuroni che si attivano quando l’azione osservata coincide con quella eseguita. TEORIA AIM La teoria Aim prevede che già i neonati codificano le azioni con un sistema sovra modale (ossia indipendente da una modalità specifica come quella percettiva o motoria) innato, il quale unifica l’osservazione e l’esecuzione degli atti motori. Questa teoria è confermata dal fatto che i neonati di pochissimi giorni sono già in grado di imitare movimenti manuali e facciali e Meltzoff, con i suoi collaboratori, ha scoperto che essi sono capaci di imitare un modello anche in seguito, ossia anche quando non hanno imitato l’azione di un modello mentre lo osservavano. Pertanto, prendendo in considerazione tali esempi, è possibile dedurre che la capacità di imitare è innata. Meltzoff e i suoi collaboratori sostengono che l’imitazione costituisce la base per la rappresentazione del pensiero altrui, per l’empatia e per la comprensione dei sentimenti altrui e, in aggiunta a ciò, hanno affermato che la formazione della teoria della mente avviene mediante 3 stadi. In primo luogo, l’equivalenza tra gli stati propri e quelli altrui costituisce la base dalla quale partire per la costruzione della teoria della mente. In secondo luogo i neonati imparano ad associare a determinati comportamenti il loro stato d’animo interno e, infine, il terzo stadio coincide con quello che da l’avvio allo sviluppo della teoria della mente: ossia la capacità di comprendere che i comportamenti degli altri sono associati agli stessi stati mentali ai quali si associano quelli propri. Inoltre, secondo i sostenitori di questa teoria, l’imitazione motoria gioco un ruolo chiave perché costituisce l’anello mancante tra gli studi condotti sui neuroni mirror e quelli fatti sulla teoria della mente. Le tesi di Meltzoff e dei suoi collaboratori sono state oggetto di numerose critiche, sia sul piano teorico che su quello empirico. Sul piano teorico, alcuni autori hanno criticato la teoria AIM perché ritengono che non si può affermare con certezza che l’imitazione motoria aiuti a colmare la lacuna che c’è tra gli studi sui neuroni specchio e quelli sulla teoria della mente. Heyes ha poi affermato che Meltzoff e i suoi colleghi hanno proposto un ruolo troppo specifico dell’imitazione per lo sviluppo della mente; mentre Jacob e Jeannerod sostengono che per valutare la tesi di Meltzoff e dei suoi colleghi è opportuno chiarire- prima- cosa si intende per imitazione. Infatti se con il termine imitazione si indica l’atto di copiare i movimenti e i comportamenti altrui, tale teoria non vale perché i movimenti non sono copiabili. Se il termine imitazione include anche delle interpretazioni creative, la situazione sarebbe diversa perché – in questo caso- anche gli stati d’animo interni potrebbero essere copiati. Anche da un punto di vista empirico la teoria AIM non trova un grande sostegno: infatti gli studi di Anisfeld hanno dimostrato che gli unici movimenti che i neonati riescono ad imitare sono quelli di protrusione della lingua. Poi numerosi studi hanno messo in dubbio che l’imitazione sia innata, mettendo- così- in evidenza che essa è una capacità che il soggetto acquisisce con l’esperienza, e cioè osservando gli altri che eseguono un’azione. Anche se tali esperimenti sembrano dimostrare che l’imitazione è frutto dell’esperienza, ancora non hanno chiarito al 100% se l’automaticità del comportamento imitativo si sviluppa con il tempo, e quindi viene appresa, o se sia innata. Infine, un altro limite della teoria AIM consiste con il fatto che le evidenze non tengono conto che i comportamenti imitativi spesso non sono fedeli ai modelli osservati, ma presentano delle variazioni; e che i bambini hanno una capacità di memoria diversa x le azioni dotate di senso e quelle prive di senso. TEORIA DELL’IMITAZIONE BASATA SU SCOPI Prima di comprendere la teoria dell’imitazione basata su scopi è fondamentale introdurre una teoria che non si limiti alla spiegazioni del comportamento imitativo, ma che fa riferimento anche al rapporto che intercorre tra percezione e azione. Secondo le teorie tradizionali tra percezione e azione c’è una certa differenza, mentre una serie di studi recenti hanno dimostrato che queste 2 attività non possono essere distinte e – addirittura- che l’una influenza l’altra. Una teoria che sostiene quest’ultima testi è la teoria ideomotoria e la teoria del codice comune, la quale si sviluppa partendo proprio da questa. La teoria ideomotoria parte dal principio che alla base dell’agire c’è sempre uno scopo, un’intenzione, e cioè l’intenzione di modificare l’ambiente che ci circonda e gli eventi presenti in esso. In pratica, secondo la teoria del codice comune i contenuti percettivi e le intenzioni d’azione vengono generati all’interno di una stessa rappresentazione mentale ed è proprio in questa prospettiva che gli stimoli che percepiamo possono essere intesi come eventi che hanno luogo nell’ambiente e, nello stesso tempo, anche le azioni che compiamo rappresentano altri eventi che hanno luogo nell’ambiente. Sulla base di questa teoria sono state formulate diverse predizioni supportate da ricerche empiriche. Prima di tutto, dall’idea del codice comune deriva il principio secondo il quale maggiore è la similarità che c’è tra le azioni che osserviamo e quelle che possediamo nel nostro repertorio di azioni possibili, maggiore sarà la quantità di codici attivati. La conseguenza di ciò è che noi tendiamo a riconoscere più facilmente le nostre azioni rispetto a quelle degli altri in quanto la somiglianza tra gli eventi che percepiamo e quelli rappresentati è alta. Pertanto quando noi osserviamo le nostre azioni, percepiamo azioni che eseguiremmo nello stesso modo; mentre se osserviamo le azioni degli altri ciò non accade. Degli ulteriori studi basati sul rapporto tra percezione e azione hanno – ancora- dimostrato che un soggetto è in grado di predire meglio gli effetti futuri delle proprie azioni che quelli delle azioni altrui e ciò è molto importante non solo da un punto di vista motorio, ma anche da una prospettiva psico-sociale in quanto riconoscere se stessi come agenti, come causa di un’azione e come soggetti capaci di gestire e prevedere gli effetti della propria azione è basilare per la formazione del sé e della propria identità. Un altro ambito di studi, ossia quello legato alla teoria ideomotoria, si propone di capire come le nostre azioni possono essere influenzate da quelle degli altri. La teoria ideomotoria parte dal presupposto che le azioni altrui sono codificate nei termini degli eventi percettivi che essi determinano; di conseguenza si registra un effetto di priming quando un essere umano osserva un atto motorio congruente con il proprio repertorio di azioni motorie ed è per questo che maggiore è la somiglianza tra i movimenti osservati e quelli percepiti, più rapidamente si riconoscono le azioni. Ne consegue che nel momento in cui un soggetto osserva le azioni eseguite dagli altri si attivano le strutture rappresentative che vengono messe in atto nel momento in cui si pianificano queste stesse azioni. Inoltre l’imitazione è stata assimilata ad un caso di compatibilità stimolo-risposta, il quale mette in evidenza che si producono risposte più veloci ad uno stimolo nel momento in cui stimolo e risposta condividono dei tratti. Un'altra teoria importante è la teoria duale e basata su scopi, la quale fa riferimento alla teoria del codice comune. Con il termine “duale” si intende che un soggetto deve acquisire –prima di tuttoassociazioni bidirezionali che ci sono tra i movimenti e gli effetti da essi prodotti, in modo da ottenere un’integrazione tra codici percettivi e codici motori. In seguito, se si decide di eseguire l’azione si attivano i codici percettivi relativi all’obiettivo che si raggiunge con tale azione e l’attivazione di tali codici provoca l’attivazione automatica dell’azione corrispondente. Questa visione è in linea con la teoria basata su scopi, la quale è molto importante perché tiene conto anche dei limiti cognitivi dovuti ad un carico eccessivo in memoria che spinge i bambini a non imitare in modo passivo un’azione osservata, ma a decomporla e ricomporla secondo una gerarchia di scopi. Da ciò si evince che i soggetti, nel momento dell’imitazione, non si limitano ad analizzare le azioni nei loro aspetti fisici, ma si concentrano anche sugli scopi ad esse soggiacenti perché nel momento in cui più scopi sono in competizione, l’individuo deve avere la possibilità di scegliere di raggiungere quello più importante trascurando quello meno importante. Ciò è possibile perché gli scopi sono ordinati gerarchicamente. In base alla teoria basata su scopi l’imitazione non è un processo unitario, ma un processo che si compone in due fasi: una di decomposizione e una di ricomposizione del pattern motorio in una serie di costituenti. Questo modello a due vie e la teoria basata su scopi hanno trovato una conferma negli studi di Tessari e Rumiati, le quali con un loro esperimento, in cui chiedevano ai partecipanti di imitare azioni sensate e azioni prive di senso, hanno messo in luce che esistono due meccanismi imitativi: una forma di imitazione diretta, utilizzata per le azioni nuove e prive di senso; una forma di imitazione per cui si fa riferimento alla memoria a lungo termine, le quali non vengono utilizzate in modo fisso, ma in relazione alla situazione che si sta vivendo e affrontando. CAPITOLO VII MOVIMENTO, OGGETTI E CONCETTI Anche se apparentemente sembra non esserci un contatto tra concetti, oggetti e movimento, in realtà non è così in quanto i concetti che ognuno di noi si forma sugli oggetti si basano su processi sensomotori e attivano, in modo automatico, un’informazione motoria. I concetti, secondo Murphy, possono essere definiti come le unità minimali della nostra conoscenza ed essi sono dati dalle informazioni che abbiamo in memoria in relazione alle categorie. Nell’ambito della scienza cognitivi ci sono teorie che hanno posizioni diverse per quanto riguarda il rapporto percezione-azione. Da un lato ci sono coloro che considerano la percezione e l’azione come 2 attività separate tra loro e distinte dai processi cognitivi superiori come la cognizione e l’organizzazione delle conoscenze. Dall’altro lato ci sono coloro i quali sostengono che tra azione e percezione c’è una stretta interazione e che tra gli aspetti senso-motori e quelli cognitivi superiori c’è una forte vicinanza. In accordo a queste posizioni sono stati proposti vari modelli i quali sostengono che esiste un’interazione tra percezione, azione e conoscenza e che la cognizione non può prescindere né dal corpo, né dalle esperienze senso-motorie ad esso associate. In questo quadro sono stai proposti modelli che intendono i concetti come fondati sull’esperienza senso-motoria e che non parlano di una rappresentazione motoria che ha un formato diverso da una rappresentazione neurale, in questo senso i concetti consistono nella riattivazione del pattern di attivazione neurale che si ha nel momento in cui si percepiscono i loro referenti o si interagisce con essi. Ad esempio, il concetto di tazza consiste nella riattivazione del pattern neurale che si attiva quando un soggetto ha un’esperienza con una tazza. Questi modelli, che enfatizzano il ruolo dei processi motori nella formazione della nostra conoscenza, consentono di capire meglio qual è la funzione dei concetti: quella di fornire un sostegno per facilitare l’interazione con gli oggetti e le entità che circondano l’individuo. In questo senso si può dire che i concetti sono fondati sulla percezione e sull’azione e –inoltre- incorporano informazione motoria. L’ipotesi che i concetti sono fondati sull’azione è sostenuta da due punti di vista diversi, ma non incompatibili. Il primo punto di vista (quello di Glenberg) sostiene che i concetti degli oggetti sono direttamente finalizzati all’azione ed è per questo che vanno intesi come pattern di azione possibile. Il significato degli oggetti è determinato dalle loro stesse proprietà ed è condizionato dalle restrizioni e dai vincoli imposti dal nostro corpo o dalle esperienze precedenti avutesi con quel tipo di oggetto. Da ciò si deduce che un oggetto come la tazza sono rappresentati –direttamente- in funzione dell’azione che può essere compiuta con essi e da ciò ne consegue che i concetti incorporano e attivano direttamente un’informazione motoria, la quale è importante perché induce un soggetto a rispondere più velocemente ad uno stimolo. Infine è importante sottolineare che i concetti, in questa prospettiva, sono fondati sull’azione e svolgono una funzione di supporto x l’azione stessa e, nello stesso tempo, permettono al soggetto di riattingere alle esperienze passate con gli stessi oggetti o con oggetti simili, facilitando – così- l’interazione con nuovi esmplari appartenenti alla stessa categoria. Il secondo punto di vista (quello di Barsalou) mette in evidenza che i concetti sono rappresentati tramite simboli percettivi, i quali rimandano direttamente alle esperienze sensomotorie con gli oggetti. Ad esempio, il concetto di tazza sarà dato dalla riattivazione di di tutte le sue proprietà percettive, quali: il colore, la forma e l’orientamento; e in funzione della situazione in cui un individuo si trova saranno selezionate le proprietà percettive utili x svolgere l’azione. Ad esempio, se la tazza serve per nasconderci qualcosa sotto io andrò ad attivare delle proprietà percettive diverse a quelle che attiverei se la tazza mi servisse per rompere una noce. CONCETTI ASTRATTI E MOVIMENTO: IL TEMPO L’esperienza del movimento e del muoversi nello spazio è molto importante per la comprensione di concetti astratti come il tempo. Ciò è stato indicato da alcuni studi recenti, i cui risultati sostengono le teorie “embolied” della conoscenza, secondo le quali i concetti astratti vengono compresi per analogia a partire da esperienze dirette e concrete, da schemi immaginativi e motori. Questo è stato dimostrato con alcuni studi condotti da diversi autori, nei quali si chiedeva ai soggetti di risponder ad una domanda ambigua: l’incontro di mercoledì è stato spostato di 2 giorni. Quando si terrà? La risposta non poteva essere una, ma dipendeva da come si concettualizza il tempo e dalla prospettiva che si assume nel rapportarsi al tempo: chi pensa al proprio movimento nel tempo, cioè chi assume una prospettiva ego-moving, tende a rispondere venerdì; chi – invece- pensa al movimento del tempo verso di sé assume una prospettiva time-moving e risponderà lunedì. In condizioni normali i partecipanti hanno uguale probabilità di adottare una delle due risposte; in condizioni sperimentali meno. Infatti se viene richiesto di sedersi su una sedia a rotelle e muoversi verso un punto nello spazio (prospettiva ego-moving) la max parte delle risposte sarà venerdì; se –invece- viene richiesto di tirare una sedia verso di sé (prospettiva time-moving) la max parte delle risposte sarà lunedi. Dall’analisi di questi risultati gli studiosi hanno dedotto che il nostro modo di concepire il tempo in termini di movimento può essere determinato dall’attività che si sta svolgendo. Oltre a questo esperimento ne sono stati svolti molti altri in contesti ecologici (come la fila in attesa di una partenza o un viaggio in treno), i quali hanno dimostrato- ancora una volta- che il nostro sistema di conoscenza temporale è costruito sulla base della nostra esperienza di movimento nello spazio. Pertanto si può concludere che anche la nostra conoscenza astratta si fonda sull’esperienza. CONCETTI DI OGGETTI E AFFORDANCES Per mettere in luce la relazione che c’è tra processi percettivi e processi motori è stata rivalutata – recentemente- la nozione di “affordance”: termine inglese che deriva dal verbo to affar (offrire) il quale mette in evidenza che gli oggetti presenti nell’ambiente offrono delle indicazioni che ci permettono di agire con essi, e cioè ci invitano a compiere determinati movimenti o azioni. Ad esempio, la maniglia della porta ci invita ad essere impugnata, la porta ci suggerisce di essere spinta o tirata per essere aperta. Nonostante ciò è importante sottolineare che le affrodances non sono caratteristiche degli oggetti in sé, ma variano a seguito dell’interazione con il nostro sistema percettivo, con il nostro corpo e con l’ambiente che ci circonda. Ad esempio, una fessura di un distributore di bibite può rappresentare un affordance di inserimento di una moneta x un adulto che consoce la funzione del distributore, mentre può rappresentare un affordance diverso x un bambino. In letteratura ci sono alcuni autori i quali sostengono che le proprietà visive di un oggetto attivano direttamente il sistema motorio senza passare attraverso la mediazione della conoscenza concettuale, ossia senza riconoscere gli oggetti. Contro questa ipotesi ci sono altri studiosi che si chiedono che se la visione degli oggetti attivasse direttamente l’informazione motoria, che ruolo avrebbero i concetti? L’autore maggiormente convinto dell’assenza della mediazione concettuale è Gibson, secondo il quale il nostro sistema visivo informa direttamente il sistema motorio, che risponde automaticamente. La conseguenza di ciò è che nel momento in cui un soggetto interagisce con gli oggetti che lo circondano non è necessario riconoscerli o essere consapevole di ciò che ha di fronte. Tuttavia la posizione di Gibson non è stata completamente accettata in letteratura, in quanto ad esso si sono opposti diversi studiosi i quali si sono chiesti che se la visione degli oggetti attiva direttamente l’informazione motoria, che ruolo hanno i concetti? E, in più, come si potrebbe continuare a pensare che i concetti rappresentano un supporto per l’interazione tra l’uomo e gli oggetti presenti nell’ambiente? VIA DEL WHAT E VIA DEL WHERE Prima di soffermarsi su quelle posizioni che sostengono l’utilità e la centralità dei concetti è importante far presenti- prima- gli studi fatti sull’elaborazione degli stimoli visivi, in riferimento alla quale sono stati distinti due sistemi diversi: uno deputato al riconoscimento degli oggetti, l’altro finalizzato alla loro specializzazione spaziale. In linea con questa prospettiva, più di 20 anni fa, due studiosi hanno distinto due vie neurali che partivano dalla corteccia visiva primaria: 1) la via visiva ventrale (comunemente riconosciuta come via del what) responsabile del riconoscimento degli oggetti; 2) la via visiva dorsale (via del where) dedicata alla posizione degli oggetti nello spazio. Intorno al 1995 altri due studiosi (Milner e Goodale) hanno rivisto tale distinzione e hanno elaborato una teoria secondo la quale esistono 2 sistemi separati che analizzano le caratteristiche visive degli oggetti per obiettivi distinti: la via ventrale (o via del what) deputata al riconoscimento degli oggetti; la via dorsale (o via dell’how) che giunge le interazioni motorie con gli oggetti. Gli studi di Milner e Goodale sono molto particolari perché questi due autori più che focalizzarsi sul diverso tipo di informazione prodotta dai sistemi, si sono concentrati sul modo in cui l’informazione viene trasformata in funzione dell’output e hanno affermato che: se la rappresentazione degli oggetti deve essere stabile nel tempo è attiva la via ventrale; se la rappresentazione avviene in modo immediato si attiva la via dorsale. Infine un’altra particolare distinzione è quella di Jeannerod, il quale ha distinto la rappresentazione degli oggetti pragmatica da quella semantica perché la prima è puramente motoria, la seconda è legata al significato. DALLA PERCEZIONE ALL’AZIONE: UNA VIA DIRETTA O INDIRETTA? Molti studiosi si sono chiesti come avviene il passaggio dalla percezione visiva all’azione e a tale quesito si è risposto affermando che esistono due vie che portano dalla percezione all’azione. Esse sono: la via diretta percezione-azione, mediata dal sistema dorsale; la via indiretta che passa – prima- dal lobo temporale, presupponendo il riconoscimento degli oggetti e l’accesso all’informazione concettuale; poi dal lobo parietale. L’esistenza della via diretta percezione-azione sembra essere confermata oltre che da evidenze comportamentali, anche da studi condotti sui pazienti; ma – nello stesso tempo- c’è una teoria la quale dimostra che questa via diretta (non semantica) svolga funzioni limitate: a tal proposito alcune prove recenti mettono in luce che questa via si attiva per oggetti nuovi o molto semplici, mentre per gli oggetti complessi è l’accesso al sistema semantico che garantisce l’interazione tra essi e i soggetti. In questo contesto è opportuno far riferimento ad un esperimento del 2001 di Creem e Proffit in cui i partecipanti sono stati divisi in 3 gruppi: uno di controllo e due gruppi sperimentali, i quali avevano tutti un compito principale: quello di afferrare e spostare oggetti impugnandoli con la mano. Ai 2 gruppi sperimentali, oltre a questo compito, ne venne affidato un secondo: al primo gruppo un compito di natura spaziale; al secondo un compito di natura semantica. Il risultato di quest’esperimento è stato: che coloro che svolgevano il compito secondario di natura semantica compromettevano la corretta esecuzione del compito primario, mentre coloro che svolgevano un compito secondario di natura spaziale non presentarono nessuna differenza con il gruppo di controllo, il quale ha effettuato il compito principale in modo appropriata. L’assenza di peggioramento della performance del primo gruppo ha portato gli autori a pensare che mentre la capacità di afferrare gli oggetti non implica necessariamente l’accesso al sistema semantico; la capacità di interagire con essi in modo appropriato e tenendo conto della loro funzione lo presuppone. Di conseguenza, per afferrare un oggetto in modo appropriato, un soggetto ha bisogno sia di una rappresentazione che indichi dove afferrarlo sia di una rappresentazione che, basandosi sulla comprensione dell’identità dell’oggetto, indichi come afferrarlo. Infine è possibile dire che questi studi suggeriscono che l’indipendenza del sistema visuo-motorio è limitata e che c’è una stretta relazione tra sistema visivo, motorio e semantico. TECNICHE DI REGISTRAZIONE DELL’ATTIVITA’ CEREBRALE Degli studi molto recenti effettuati con tecniche di scansione computerizzata e tecniche di registrazione dell’attività cerebrale hanno messo in evidenza che la visione degli oggetti attiva informazioni motorie. Tra questi studi molto importanti sono stati quelli di Martin e dei suoi colleghi i quali, utilizzando la Pet, si sono proposti di verificare quali aree neurali si attivano durante compiti di denominazione di utensili e di animali. I risultati di questi studi hanno messo in evidenza che quando i soggetti devono denominare utensili si attivano l’area che – di solito- si attiva in compiti di produzione di nomi e azioni e l’area che si attiva quando si immagina di afferrare oggetti con la mano dominante. Altri studi importanti sono stati quelli di Chao e Martin che, impiegando la risonanza magnetica funzionale, hanno dimostrato che la corteccia premotoria sx si attiva nel momento in cui il soggetto vede immagini di utensili e non di animali, volti o case. Una questione molto dibattuta in letteratura si riferisce alla specificità o meno di concettualizzare gli oggetti, la quale può essere spiegata con una domanda: solo i concetti di utensili e di artefatti attivano informazione motoria, o il legame tra i concetti e l’azione fa riferimento anche a concetti generali? In risposta a tale questione è nata un’ipotesi secondo la quale l’attivazione di un’informazione motoria è generata da concetti di oggetti manipolabili, indipendentemente dal fatto che si tratti di artefatti o di oggetti naturali. Ciò è importante perché dimostra che le info – a livello neurale- non sono codificate in funzione di categorie rigide, ma in funzione di quello che con gli oggetti viene fatto. Quest’ipotesi è stata confermata da una serie di studi i quali hanno dimostrato che l’informazione motoria si attiva quando un soggetto vede un oggetto manipolabile perché, in questo caso, è come se egli simulasse mentalmente l’azione che si svolge con quell’oggetto. INFORMAZIONE MOTORIA: con il termine informazione motoria si può intendere sia il modo con cui interagiamo con gli oggetti, sia il modo in cui li usiamo x raggiungere degli scopi specifici. Siccome non sempre c’è una corrispondenza tra sapere come interagire con un oggetto e sapere a che cosa serve un oggetto e come va utilizzato, la letteratura suggerisce che la visione di un oggetto attiva un’informazione relativa a come raggiungerlo e manipolarlo, ossia a come interagire con esso; mentre l’informazione funzionale viene attivata solo nel momento in cui l’informazione relativa alla forma e alla funzione evoca la stessa risposta motoria. La presenza di una doppia dissociazione tra l’informazione relativa all’azione e a quella relativa alla funzione sembra essere confermata anche da recenti studi neurofisiologici, i quali hanno messo in evidenza che il cervello preferisce rispondere meglio all’informazione relativa al come manipolare gli oggetti, piuttosto che a quella relativa alla loro funzione. STUDI COMPORTAMENTALI L’ipotesi secondo la quale la visione degli oggetti provoca l’attivazione del sistema motorio è stata confermata da una serie di studi comportamentali, i quali hanno messo in luce l’influenza del modo in cui un soggetto interagisce con gli oggetti e sul modo in cui li categorizza. Tra questi studi, molto importanti sono quelli che hanno indagato la preferenza costante x la forma, ossia la tendenza ad attribuire lo stesso nome ad oggetti che hanno una forma simile. Questo studio concorda con la teoria di Biederman, secondo la quale gli oggetti sono riconosciuti in virtù delle parti che li compongono, e cioè per la loro forma, e in seguito per le loro proprietà. La centralità della forma è, poi, strettamente legata alla sua rilevanza per l’azione/interazione con gli oggetti; infatti uno studio recente ha dimostrato che il modo in cui ci formiamo le categorie è influenzato dal tipo di azione che compiamo con un determinato oggetto. Infine, altri studi comportamentali hanno confermato e integrato le prove ottenute con le tecniche di registrazione dell’attività cerebrale, le quali hanno dimostrato una sensibilità nei confronti della distinzione che c’è tra oggetti manipolabili e oggetti non manipolabili, legata al fatto che i primi attivano un’informazione motoria. Quest’ultimo punto è stato sottolineato dagli esperimenti di Saffran e colleghi, i quali hanno trovato che con oggetti manipolabili si producono più verbi che con oggetti non manipolabili. STUDI CON PARADIGMI DI COMPATIBILITA’ La relazione tra percezione e azione è stata studiata anche con l’impiego di paradigmi di compatibilità, e in questo settore, molto importanti sono stati gli studi di Tucker ed Ellis , i quali con i loro esperimenti hanno messo in evidenza che la visione degli oggetti attiva e potenzia le affordances ad essi associati. Questi due studiosi più che di affordances, hanno parlato di microaffordaces, le quali si attivano automaticamente e indipendentemente dagli scopi e – nello stesso tempo- attivano azioni specifiche per un dato tipo di oggetto. Ciò significa che la grandezza di un oggetto non provoca un comportamento di afferra mento generale, ma un movimento di afferra mento adatto a quel particolare tipo di oggetto. In un loro esperimento Ellis e Tucker hanno chiesto ai partecipanti di classificare una serie di oggetti di forma e grandezza diversa in oggetti naturali e artefatti; e il compito di questi soggetti era quello di fare pressione su un dispositivo simile ad un joystick, pressione che poteva simulare una presa di forza o una presa di precisione. Il risultato di tale esperimento ha dimostrato un effetto di compatibilità tra il tipo di presa e la grandezza dell’oggetto, anche se quest’ultimo era collocato in uno spazio che non dava al soggetto la possibilità di afferrarlo. Tale effetto suggerisce con chiarezza che vedere un oggetto porta ad attivare un movimento e a simulare uno specifico tipo di prensione. Oltre a tali esperimenti sono stati effettuati un’altra serie di studi i quali hanno dimostrato che le microaffordances non attivano solo comportamenti di prensione, ma anche movimenti di raggiungimento, sollecitati dalla forma e dall’orientamento degli oggetti. Ciò è stato dimostrato empiricamente da altri esperimenti condotti da Ellis e Tucker i quali hanno messo in evidenza che la visione di un oggetto attiva il movimento di raggiungimento e un lato di risposta. STUDI CON PARADIGMI DI PRIMING La relazione che intercorre tra sistema percettivo e sistema motorio è stata ulterioremente confermata da studi che hanno impiegato il “priming” motorio-visivo, tra i quali molto importanti sono stati quelli di Craighero e colleghi che hanno dimostrato che un compito di preparazione motoria può favorire la rilevazione e la discriminazione di forme visive. In questo esperimento i partecipanti dovevano afferrare una barra che poteva essere orientata in senso orario o antiorario . prima di quest’azione veniva presentato - ai partecipanti – l’immagine di un oggetto orientato in senso orario o antiorario e il loro compito era quello di afferrare la barra dopo la presentazione dello stimolo. Il risultato di tale esperimento ha messo in luce che un effetto di compatibilità tra l’orientamento dell’oggetto presentato visivamente e quello della barra da afferrare, a condizione che l’oggetto avesse caratteristiche percettive simili a quelle della barra. Se così non era, nel senso che l’oggetto presentato evocava un tipo di prensione diversa perché aveva caratteristiche differenti dalla barra, l’effetto non compariva in quanto esso è generato dalle affordance estratte dall’oggetto e non dal suo orientamento. Questa stretta relazione tra stimoli visivi e azioni motorie è stata ulteriormente confermata da studi molto recenti e anche con l’impiego di un paradigma di priming visuo-motorio. In quest’ultimo caso, veniva chiesto ai partecipanti di fissare un punto posto al centro dello schermo di un pc e di afferrare uno o due oggetti con la mano dx nel momento in cui il punto cambiava colore. Prima, simultaneamente o dopo il cambiamento di colore, appariva sullo schermo la figura di un oggetto che poteva essere congruente o meno con quello da afferrare. Il risultato di tale esperimento ha dimostrato che nel caso in cui c’era congruenza tra l’oggetto presentato sullo schermo e quello reale, i tempi di risposta erano molto più veloci rispetto al caso di incongruenza. Tuttavia è importante sottolineare che anche se questi studi sono molto importanti perché hanno confermato l’esistenza di tale relazione, essi non hanno preso in considerazione un problema molto sentito: quello relativo al fatto se lo stimolo visivo è sufficiente per estrarre una risposta motoria. Infatti, a tal proposito, molti sostengono che uno stimolo visivo può dar vita ad una risposta motoria, ma solo nel caso in cui c’è una preattivazione del sistema motorio o se l’esperimento è preceduto da una preparazione motoria. Tra i diversi studi che sostengono tale ipotesi è fondamentale ricordare quello di Bub, Masson e Bukach, dove ai partecipanti venivano mostrate delle fotografie che rappresentavano mani con diverse posture, seguite da oggetti associati a quella postura. Ad esempio, una mano raffigurata con una presa di precisione è seguita da un oggetto che si afferra con quel tipo di presa o con una presa di tipo diverso; il compito dei partecipanti è quello di scegliere tra l’una e l’altra presa in risposta al diverso colore dell’oggetto. La congruenza tra postura raffigurata e caratteristiche dell’oggetto si manifesta solo quando si utilizza un paradigma in cui un precedente segnale indica ai partecipanti se devono rispondere al colore o all’oggetto. Con questo esperimento gli studiosi hanno messo in evidenza che la semplice visione di un oggetto non attiva automaticamente la conoscenza gestuale-motoria. Un altro esperimento molto importante è stato quello di Borghi, durante il quale si presentavano ai partecipanti un presti molo, che coincideva con l’immagine di una mano la quale poteva assumere una postura presa di forza o presa di precisione, o una postura a mano aperta. L’immagine della mano era sempre seguita da quella di un oggetto e il compito dei partecipanti era quello di decidere se l’oggetto che seguiva l’immagine della mano era un oggetto naturale o un artefatto; mentre se veniva presentata la figura in cui la mano assumeva una posizione neutra essi non dovevano rispondere allo stimolo. Dal risultato dell’esperimento non è emerso un effetto di congruenza tra la postura della mano e l’oggetto raffigurato, dimostrando – in questo modo- che lo stimolo visivo non evoca, in modo diretto e automatico, l’informazione motoria relativa alla prensione dell’oggetto. L’effetto di congruenza tra la postura di precisione e oggetti piccoli – invece- si è presentato nel momento in cui l’esperimento è stato preceduto da una fase di addestramento. Da ciò si può dedurre che, in concomitanza con la presentazione dello stimolo visivo, è necessaria una fase di preattivazione del sistema motorio per suscitare un effetto di congruenza. Contro questi esperimenti ci sono stati gli studi di Vogt, Taylor e Hopkins i quali hanno dimostrato che l’effetto del priming visivo sul comportamento motorio c’è; ma nonostante ciò i dubbi restano in quanto non è chiaro se lo stimolo visivo da solo, ossia senza una preattivazione del sistema motorio, sia in grado di estrarre dal soggetto comportamenti motori. CAPITOLO 8 MOVIMENTO E COMPRENSIONE DEL LINGUAGGIO Come i concetti, anche le parole (le quali mediano questi ultimi) si fondano sulla percezione e sull’azione, nel senso che riattivano l’esperienza senso-motoria con gli oggetti a cui si riferiscono. L’ipotesi secondo la quale i concetti mediati dalle parole rimandano all’interazione senso-motoria con gli oggetti stessi è confermata da una serie di studi sperimentali; e ciò che vale per le parole, vale anche per le frasi e i testi: quando comprendiamo una frase che ci rimanda ad un’azione o ad un movimento, ci formiamo una simulazione interna di quell’azione o di quel movimento. PAROLE E FIGURE: DUE DIVERSE VIE DELL’AZIONE Per giungere all’azione è possibile percorrere due vie: una diretta dalla visione dell’azione (mediata dal sistema dorsale); una indiretta visione-semantica-azione (mediata dal sistema ventrale). Secondo la prima, se l’attivazione dell’informazione motoria è legata alle affordances, ossia alle proprietà degli oggetti, gli effetti che si ottengono con le figure non dovrebbero presentarsi con le parole. Secondo la seconda, le immagini attivano un’informazione motoria più velocemente rispetto alle parole, il che non esclude che le parole possono attivare un’informazione motoria. Ciò è stato dimostrato da una serie di studi, tra i quali ricordiamo: quello di Boronat e colleghi in cui i partecipanti all’esperimento (sottoposti alla Frm) dovevano decidere, in risposta a coppie di figure o di oggetti che denotavano oggetti, se si trattava di oggetti che possono essere manipolati nello stesso modo o che svolgono una stessa funzione. I risultati di questo studio hanno messo in evidenza che le aree di attivazione x gli stimoli verbali e quelli figurali erano molto simili: infatti x entrambe si attivavano vaste regioni della corteccia frontale e prefrontale sx dorso e ventrolaterale, oltre al lobo parietale inferiore sx, i quali sono tutti associati all’elaborazione senso-motoria. L’ipotesi per cui anche le parole rimandano ai processi senso-motori ed attivano informazione legata all’azione e l’ipotesi secondo la quale le figure attivano informazione motoria più direttamente delle parole, sono confermate da uno studio comportamentale recente il quale si basava sull’associazione di figure e parole che mandavano a oggetti manipolabili e non. Con questo studio gli autori hanno notato che con le figure viene prodotta una percentuale di verbi più elevata che con le parole; e che con le parole che rimandano ad oggetti manipolabili vengono prodotti più verbi rispetto alle parole che rimandano ad oggetti non manipolabili. Le parole attivano un’informazione multimodale, ossia relativa a diverse modalità sensoriali. Su questo punto sono stati effettuati una serie di studi i quali hanno messo in evidenza che il passaggio da una modalità sensoriale all’altra richiede dei costi al soggetto. In un esperimento, durante il quale venivano presentate ai partecipanti 2 figure e il loro compito era quello di scegliere se la seconda denotava una proprietà della prima, è stato dimostrato che x verificare una proprietà si è più lenti se si è appena verificata una proprietà di modalità diversa; più veloci se si è verificata una proprietà di modalità analoga a quella precedente. Ciò dimostra che cambiare modalità comporta un costo anche con le parole. Facendo un’analisi di tutti questi studi compiuti si può dedurre che l’attivazione automatica dell’informazione motoria non è dovuta solo all’azione delle affordances, ma anche dall’influenza che ha l’informazione semantica. Infine si può dire che, in questo modo, vengono confermate entrambe le ipotesi esposte in precedenza: sia quella secondo cui le figure attivano informazione motoria in modo più veloce rispetto le parole; sia quella secondo la quale le parole attivano direttamente un’informazione motoria. NOMI E VERBI Una vasta letteratura sulle basi neurali della conoscenza indaga i correlati neurali dei nomi e dei verbi. Su tale questione ci sono dei punti di vista divergenti: infatti, da un lato ci sono una serie di studiosi di neuropsicologia i quali hanno messo in luce che lesioni ad aree diverse provocano l’incapacità di utilizzare l’una o l’altra categoria grammaticale. Dall’altro lato ci sono coloro che hanno effettuato degli studi con tecniche di scansione cerebrale, i quali hanno messo in dubbio il fatto che diverse aree neurali siano dedicate a diverse categorie grammaticali e hanno affermato he le conoscenze concettuali sono distribuite in diverse aree neurali, ma non sono differenziate in funzione della classe grammaticale. Uno studio molto interessante è quello di Pulvermuller, Harle e Hummel, i quali sostengono l’ipotesi secondo la quale le parole sono organizzate in reti che riflettono i loro referenti. Se così fosse, nel momento in cui si pronuncia la parola “calciare” dovrebbero attivarsi, oltre che i neuroni delle aree del linguaggio anche i neuroni coinvolti nella coordinazione del movimento del calciare. Nell’esperimento condotto da questi studiosi veniva chiesto ai partecipanti di eseguire compiti di decisione lessicale, ossia di decidere se le parole presentate appartenevano o meno alla loro lingua. Dopo aver confrontato sia i tempi di reazione, sia il pattern (ossia modello regolare) di attivazione corticale relativo ai verbi legati alle azioni, è stato messo in evidenza che le parti del corpo cui questi verbi rimandavano erano riflesse nel pattern di attivazione corticale provocato da querste parole. Tale risultato mette in luce che le sotto-categorie dei verbi di azioni eseguite con parti del corpo diversi, sono distinte anche da un punto di vista neurofisiologico. EFFETTI DI COMPATIBILITA’ CON LE PAROLE Nel momento in cui un soggetto interagisce con un oggetto, le sue azioni dipendono solo dalle caratteristiche visive dell’oggetto o anche dalla riattivazione di esperienze visuo-motorie avute precedentemente con oggetti simili? Per dare una risposta a tale quesito sono state formulate 2 ipotesi: la prima secondo cui i risultati di studi con paradigmi di compatibilità si spiegano mediante una via diretta, x cui l’informazione diretta viene attivata direttamente dagli stimoli visivi; la seconda che fa ricorso ad una via indiretta, che prevede il coinvolgimento della memoria e delle conoscenze a lungo termine. Una serie di studi recenti hanno dimostrato che nel momento in cui bisogna riconoscere oggetti nuovi viene attivata la via dorsale; mentre nel momento in cui c’è un’interazione con gli oggetti conosciuti viene attivata la via ventrale. Gli studi sull’elaborazione di parole che rimandano ad oggetti hanno fornito un importante contributo x sciogliere la controversia che esiste tra chi sostiene che effetti di compatibilità sono attribuibili all’attivazione del solo sistema dorsale e chi – invece- sostiene che tali effetti sono spiegabili grazie alla mediazione del sistema ventrale. Tali studi, infatti, hanno messo in evidenza che se gli effetti di compatibilità si manifestano anche con le parole non è possibile escludere l’intervento del sistema ventrale e, quindi, della MLT. Ciò è quello che ammettono Tucker ed Ellis, i quali – in un loro esperimentohanno trovato effetti di compatibilità tra la grandezza di un oggetto e il tipo di presa, pronunciando solo parole che rimandano a quegli oggetti e senza utilizzare figure. Da ciò si è dedotto che la MLT relativa ad associazioni tra oggetti e azioni sembra spiegare gli effetti di compatibilità, almeno per la prensione. I risultati di Tucke ed Ellis sono importanti perché spiegano la presenza di effetti di compatibilità per quello che riguarda la prensione; ma lasciano in sospeso la questione legata al fatto se x le proprietà estrinseche, come l’orientamento degli oggetti, l’informazione motoria può essere attivata in modo diretto e automatico o attraverso la mediazione della conoscenza a lungo termine. Su questo punto si trova una risposta in uno studio recente (effettuato da Borghi e colleghi), il quale dimostra che anche per l’orientamento degli oggetti, la MLT legata alle interazioni tra oggetto e azione è fondamentale x spiegare l’effetto delle affordances. Dall’analisi di tutte queste ricerche si può dedurre che sia x quanto riguarda la postura, sia per quanto riguarda il raggiungimento, gli effetti di compatibilità sono spiegati da associazioni presenti nella MLT - tra gli oggetti e le azioni che, generalmente, si compiono con essi. Questo conferma la teoria senso-motoria, secondo la quale i concetti non sono simboli arbitrari, ma riattivano riattivano la varietà delle esperienze e di modalità con cui i concetti sono stati acquisiti; e un discorso simile vale x le parole, il cui significato non è spiegabile in base a reti arbitrarie dio associazioni con altre parole, ma rimanda ad un referente e all’esperienza senso-motoria con quel referente. Pertanto la presenza di effetti di compatibilità non è dovuta al solo coinvolgimento del sistema dorsale, ma anche del sistema ventrale. Tuttavia a questa affermazione se ne affianca una alternativa (quella di Gentilucci) la quale mette in evidenza che nel sistema dorsale può avere luogo una qualche forma di rappresentazione dell’oggetto, che codifica le modalità di interazione più comuni. Qualunque delle due ipotesi sia corretta è importante sottolineare che questi effetti di compatibilità trovano una spiegazione sulla base di associazioni visuo-motorie a lungo termine che si instaurano tra gli oggetti e le azioni che vengono effettuate con essi. Ciò può avvenire attraverso 2 modalità: la prima, che coincide con l’ipotesi indessicale, le parole attivano info percettive e visive sugli oggetti, le quali – a loro volta- attivano informazioni motorie. Secondo l’altra ipotesi le parole attivano direttamente associazioni visuo-motorie. In conclusione si può dire che le parole attivano informazioni motorie, anche se resta aperta la possibilità che gli effetti di compatibilità prodotti dalle parole sono meno forti di quelli prodotti con le figure. EFFETTI DI COMPATIBILITA’ E CINEMATICA Gli studi descritti sino ad ora non hanno mai preso in considerazione la risposta motoria fornita dai partecipanti ed è per questo che è importante far riferimento a degli studi recenti i quali si sono proposti di indagare il rapporto che c’è tra il significato delle parole e le caratteristiche cinematiche della risposta motoria. Più precisamente, lo scopo di questi studi era quello di analizzare in che misura il significato delle parole influenza la cinematica relativa al raggiungimento o alla prensione degli oggetti e in quale momento del movimento avviene tale influenza. In quest’ambito è necessario far riferimento ad un esperimento di Gentilucci, il quale ha dimostrato che le parole hanno un’influenza sul raggiungimento e la prensione di un oggetto e ha specificato che tale influenza avviene soprattutto nelle fasi iniziali. Inoltre, in uno studio successivo, Gentilucci ha dimostrato che i verbi influenzano il controllo dell’azione in modo più forte rispetto agli aggettivi. Pertanto, compiendo un’analisi generale su questi studi, si deduce che essi provano che il significato delle parole influenza il sistema motorio e precisano in quale fase dell’esecuzione del movimento si ha l’influenza della semantica sull’attività motoria. PAROLE CON VALENZA EMOTIVA E MOVIMENTO Un’altra vasta gamma di ricerche hanno messo in evidenza che eseguire un movimento mentre si apprende una parola può portarci a valutarla positivamente o negativamente. Tra questi esperimenti importanti sono stati quelli di Caccioppo, Priester e Berntson, iu quali hanno dimostrato che gli individui avevano un atteggiamento positivo nei confronti di ideogrammi cinesi a loro sconosciuti se disponevano le braccia in una posizione che – di solito- indica accoglienza; e un atteggiamento più negativo se estendevano le braccia eseguendo un movimento di allontanamento dal propriio corpo. Gli effetti della flessione e dell’estensione delle braccia fungevano da stimoli per le parole nuove o x le non parole, ma non erano efficaci per le parole familiari, in quanto queste erano già connotate emotivamente sulla base di esperienze passate. In un altro studio ai partecipanti venivano presentate parole con valenza positiva o negativa e il loro compito era quello di determinare la valenza tirando, verso di sé, o allontanando una leva. Nel momento in cui la parola e il gesto erano congruenti, i tempi di risposta erano più veloci; in caso contrario i tempri di risposta erano più lenti. Lo stesso risultato si è ottenuto anche quando il compito dei partecipanti non era quello di determinare la valenza della parola, ma solo nel rispondere ad quando esse apparivano sullo schermo di un Pc. Un'altra serie di studi simili sono stati effettuati nel tempo per dimostrare l’ipotesi iniziale: ossia che un movimento può influenzare la valutazione (positiva o negativa) che un soggetto da ad una parola. COMPRENSIONE DI FRASI E TEORIA DELLA SIMULAZIONE La teoria della simulazione ha avuto successo negli ultimi anni, durante i quali ci si è resi conto che molti processi fondati sulla simulazione sottostanno alla comprensione delle azioni, intenzioni ed emozioni altrui e all’attribuzione ad altri stati. Quando si parla di linguaggio non si fa riferimento alla simulazione in generale, ma ad una particolare accezione della nozione di simulazione: quella applicata agli studi della comprensione del linguaggio. In accordo alla teoria della simulazione, quando comprendiamo una frase riproduciamo mentalmente la situazione descritta; e cioè si riattivano i pattern di attivazione neurale che si attivano durante la percezione e l’interazione con gli oggetti descritti dalla frase. Ciò è stato dimostrato da un esperimento di Stanfield e Zwann, i quali hanno presentato ai partecipanti delle frasi in cui l’immagine principale era un chiodo che poteva assumere un orientamento verticale o orizzontale. In seguito, veniva presentata sullo schermo del Pc l’immagine di un oggetto e il compito dei partecipanti era quello di decidere se l’oggetto rappresentato era quello della frase. L’aspetto manipolato era l’orientamento degli item critici, i quali non avrebbe dovuto avere nessuna influenza sui risultati. Invece alla fine dell’esperimento si è notato che nel momento in cui l’orientamento dell’oggetto sia nella frase che nell’immagine collimavano, i tempi di risposta erano più veloci rispetto a quando tali orientamenti non erano uguali. Tale risultato suggerisce che c’è una stretta relazione tra informazione linguistica e informazione percettivo-motoria e che la comprensione di una frase comporta la simulazione mentale di come gli oggetti possono essere disposti, in modo da interagire meglio con essi. Utilizzando un paradigma sperimentale simile a quest’ultimo, questi stessi studiosi hanno dimostrato che noi non ci rappresentiamo solo l’orientamento degli oggetti, ma anche la loro forma. Un ulteriore conferma alla teoria della simulazione è stata data dagli studi di Spivey e Geng, i quali hanno registrato i movimenti oculari di partecipanti che, posti di fronte ad uno schermo bianco, ascoltavano il racconto di una scena. Le scene erano costruite in modo da raggiungere una direzionalità e i partecipanti tendevano a muovere gli occhi nella direzione che veniva descritta. La cosa interessante è stata quella che tale fenomeno si è verificato anche quando i partecipanti tenevano gli occhi chiusi, quando non sapevano che si stavano registrando i loro movimenti oculari e quando non veniva chiesta loro di immaginare la situazione descritta. Ciò dimostra che i movimenti oculare non sono azioni utili a raccogliere una serie di info, ma rappresentano il legame tra le rappresentazioni interne e i loro referenti, i luoghi esterni, le entità esterne. In questo settore una questione molto dibattuta è quella relativa a quali meccanismi entrano in gioco nel momento in cui si comprende una frase, in quanto molti hanno ipotizzato che la comprensione di una frase sul movimento scaturisce l’attivazione di quegli stessi neuroni deputati alla percezione di un movimento. Tutto ciò sembra essere stato dimostrato da un esperimento di Kaschack, durante il quale venivano presentate ai partecipanti delle frasi che dovevano essere giudicate sensate o meno. Le frasi sensate seguivano una specifica direzione (di avvicinamento, di allontanamento, o un movimento verso il basso, l’alto o di lato) e contemporaneamente ad esse venivano mostrati degli stimoli che riproducevano un movimento nella stessa direzione o in quella opposta. Il risultato di questo esperimento ha messo in evidenza che i tempi di risposta erano più lenti nel giudicare sensata una frase quando c’era una concomitanza tra la direzione dell’immagine mostrata e quello dello stimolo. Questo risultato ha messo in luce il fatto che nel simulare la frase vengono implicati gli stessi neuroni coinvolti nel percepire il movimento in un data direzione; e ciò sembra entrare in contraddizione con altri studi i quali hanno messo in evidenza che la lettura di frasi sul movimento attiva il sistema motorio, facilitando la risposta piuttosto che creare un’interferenza. Una spiegazione a ciò è stata data proproio da Kaschak, il quale ha affermato che quando una frase e lo stimolo vengono presentati contemporaneamente, si ha unm’interferenza; se la frase viene presentata prima dello stimolo si ha una facilitazione. Un altro fattore che può determinare interferenza o facilitazione coincide con la facilità di integrare gli stimoli nella simulazione creatasi leggendo la frase. In linea con questi risultati, un lavoro importante è quello di Buccino e dei suoi colleghi, il quale mette in evidenza che la lettura di frasi che descrivono azioni modula il sistema motorio. In questo caso gli autori hanno utilizzato uno studio comportamentale associato ad uno studio in cui, tramite la Tms, si è registrata l’ampiezza dei potenziali evocati muscolari. In questo esperimento i partecipanti dovevano ascoltare frasi di 3 tipi: astratte, frasi relative ad azioni eseguite con i piedi e frasi relative ad azioni eseguite con le mani; il loro compito era quello di fornire una risposta motoria (o con la mano o con il piede) nel momento in cui ascoltavano una frase concreta, e di astenersi dal rispondere nel caso ascoltavano una frase astratta. I risultati hanno dimostrato che ascoltare frasi relative ad azioni eseguite con le mani provoca un calo dell’ampiezza dei potenziali evocati dei muscoli dei piedi; mentre ascoltare frasi relative ad azioni eseguite con le mani provoca un calo dell’ampiezza dei potenziali evocati dei muscoli delle mani. Inoltre i risultati dimostrano che risposte date con lo stesso effettore a cui fa riferimento la frase sono più lente di quelle date con un effettore diverso. Alla fine si può dire che questi studi hanno indicato che il significato delle frasi comporta una modulazione dell’attività del sistema motorio e che questa modulazione è specifica, cioè differisce in funzione dell’effettore coinvolto. COMPRENSIONE DI FRASI E AFFORDANCES La teoria della simulazione prevede che quando un soggetto comprende una frase relativa ad azioni compiute con oggetti, si riattivano le affordances di quegli stessi oggetti. Secondo la teoria indessicale ciò avviene perché le parole o le frasi rimandano agli oggetti, ai loro referenti o alle loro rappresentazioni. Dagli oggetti derivano delle affordances, che possono essere diverse anche per un solo stimolo, ed è per questo che si è giunti ad affermare che non sono le parole stesse a vincolare il modo in cui le idee possono essere combinate; bensì le affordances degli oggetti a cui le parole rimandano. Per dimostrare questo gli studiosi che hanno elaborato la teoria indessicale hanno fatto un esperimento in cui i partecipanti dovevano valutare se le frasi presentate al pc erano sensate o meno, premendo uno o l’altro tasto i quali erano associati a 2 oggetti che potevano completare la frase anche se non erano collegati semanticamente all’oggetto che andavano a sostituire. Il risultato di questo esperimento (pag.183) ha evidenziato che le frasi che rimandano ad azioni con degli oggetti comportano una simulazione interna dell’azione descritta, la quale è vincolata dalle caratteristiche percettive dell’oggetto cui rimanda. Un altro grande lavoro è stato quello di Borghi e colleghi, in cui veniva chiesto ai partecipanti di giudicare se la parola che seguiva una frase costituiva una parte dell’oggetto descritto nella frase. Il risultato di tale esperimento ha dimostrato che le parti che attivano un’azione congruente all’azione descritta nella frase portano a tempi di risposta più veloci, indipendentemente dall’associazione semantica tra la parola e la frase che la precede. Tale risultato suggerisce che i concetti sono variabili, nel senso che vengono attivate caratteristiche percettive diverse a seconda dell’azione che si compie e dell’obiettivo che si vuole raggiungere; la conseguenza è che mentre per un dato tipo di azione, una parte può essere una buona affordances, per un altro tipo di azione ciò non vale. COMPRENSIONE DI FRASI E RISPOSTE MOTORIE Mentre la comprensione delle frasi che descrivono azioni con oggetti porta a simulare internamente l’azione descritta, la lettura di frasi relativa ad azioni, attiva direttamente un tipo di movimento. Ciò è dimostrato dagli studi di Glenberg e Kaschak, i quali chiedevano ai partecipanti di valutare la sensatezza o meno delle frasi e di rispondere si o no premendo un pulsante. Per premere il pulsante essi dovevano allontanare o avvicinare la mano dal corpo e, confermando la teoria della simulazione, sono stati trovati effetti di compatibilità tra azioni descritte e i movimenti dei partecipanti. Tuttavia va sottolineato che quando una frase implicava un’azione che seguiva una direzione opposta a quella del movimento i soggetti avevano difficoltà a fornire un giudizio di esattezza; mentre era più facile rispondere se le due direzioni erano uguali. Lo stesso effetto di compatibilità si verificava anche con le frasi astratte. Anche in uno studio molto simile a questo, in cui si chiedeva ai partecipanti di valutare la sensatezza di coppie composte da verbo e nome, i risultati hanno confermato la teoria della simulazione: leggere frasi che descrivono situazioni in cui si afferrano oggetti, comporta una preattivazione dell’arto con cui gli oggetti vengono afferrati generalmente. COMPRENSIONE DI FRASI E MOVIMENTO DEGLI OGGETTI Una serie di ricerche recenti hanno dimostrato che nel momento in cui leggiamo o comprendiamo una frase legata ad un’azione simuliamo mentalmente quell’azione; pertanto la comprensione di frasi attiva rappresentazioni visive dinamiche le quali sono molto importanti per le azioni che bisogna compiere. Uno studio molto importante è quello di Zwann e dei suoi colleghi durante il quale si chiedeva ai partecipanti di ascoltare frasi che descrivevano un movimento o di avvicinamento di un oggetto. Dopo la presentazione della frase i partecipanti vedevano due immagini dell’oggetto presente in essa in sequenza e, premendo un tasto, dovevano indicare se le due figure rappresentavano lo stesso oggetto o meno. Nella prima delle immagini l’oggetto poteva essere più piccolo rispetto alla seconda, suggerendo un movimento di avvicinamento; o più grande, suggerendo un movimento di allontanamento. A conferma della teoria della simulazione si è trovato un effetto di congruenza tra il tipo di frase presentata e la sequenza di presentazione, ma quando la frase suggeriva un movimento di allontanamento, i tempi di reazione erano più veloci rispetto a quando la frase suggeriva un movimento di allontanamento. E’ interessante notare che la teoria della simulazione vale anche quando si prende in considerazione un movimento fittizzio; e a tal proposito, importanti sono stati gli studi di Matlock durante i quali si leggeva, ai partecipanti, una storia che descriveva un particolare scenario e, dopo, si presentavano frasi che potevano o meno contenere verbi relativi al movimento fittizzio. l’autrice ha notato che il tipo di scenario rappresentato influenzava l’elaborazione della frase che conteneva il verbo indicante il movimento fittizzio. Inoltre nel momento in cui lo scenari era ricco di ostacoli, i tempi di risposta erano più lenti rispetto al momento in cui lo scenario indicava una situazione priva di ostacoli. Questi risultati sono giunti ad affermare la tesi di Langacker secondo la quale x formarsi un concetto occorre assumere una prospettiva dalla quale partire, dalla quale si analizza la scena e si simula mentalmente il movimento. In uno studio successivo, in cui i partecipanti dovevano fare dei disegni che rappresentavano scene descritte da verbi fittizzi o da verbi che non indicavano movimento, Matlock ha dimostrato che i soggetti tendevano a disegnare figure più allungate con i verbi di movimento fittizzio e, in particolare, con quelli che indicavano movimenti veloci piuttosto che con quelli che indicavano movimenti lenti. Infine questo studioso ha messo in evidenza che i verbi fittizzi influenzano anche il tipo di prospettica che si adotta nel pensare al tempo. COMPRENSIONE DI FRASI E PROSPETTIVE RILEVANTI PER L’AZIONE Le parole che rimandano ai concetti attivano un’informazione motoria anche un modo indiretto. Infatti, tra le diverse caratteristiche percettive di un oggetto noi selezioniamo quelle che rimandano ai referenti percepiti attraverso una data prospettiva che, generalmente, è quella canonica, la quale è più rilevante per le azioni situate nei contesti. Tuttavia, oltre alla prospettiva canonica, la quale permette di rimandare i concetti a referenti percepiti da vicino, frontalmente, dall’esterno e visivamente; c’ anche la prospettiva dipendente dal contesto, che – in alcuni casi- può superare quella canonica. In una serie di esperimenti Borghi e i suoi colleghi hanno cercato di capire se i concetti rimandano a referenti rappresentati secondo diverse prospettive e se tali prospettive sono quella canonica e quella dipendente dal contesto. I risultati di tale esperimento hanno dimostrato che noi ci rappresentiamo i concetti secondo queste due prospettive in quanto tendiamo a rappresentarci gli oggetti secondo la modalità visiva, ma siamo in grado di simulare anche situazioni in cui l’oggetto si caratterizza per proprietà uditive. Infine va sottolineato che le prospettive con cui gli oggetti sono percepiti possono essere modulate dallo scenario rappresentato da una frase. Ciò è stato ulteriormente dimostrato da un esperimento di Borghi e dei suoi colleghi, il cui risultato indica che nell’elaborare frasi ci raffiguriamo mentalmente le situazioni in modo puntuale e selezioniamo informazioni rilevanti a partire da una data prospettiva. L’ATTIVITA’ SPORTIVA: MOVIMENTO E MEMORIA La memoria di movimenti può essere considerata uno specifico settore all’interno degli studi sulla memoria in generale e, in più, essa può essere vista come il punto di contatto tra due ambiti di ricerca differenti: nel primo caso essa si connette con gli studi fatti sulla memoria di movimenti semplici; nel secondo caso esiste un legame molto stretto tra il ricordo del movimento in attività sportive e gli studi sulle modalità di memorizzazione utilizzate dagli esperti. Il fattore che accomuna questi due settori è il livello di abilità/conoscenza che gli atleti professionisti possiedono nei confronti della disciplina che praticano. Alcuni studi i quali hanno messo a confronto le modalità di ricordo degli atleti professionisti con quelle di esperti in altri campi, hanno messo in evidenza che i giocatori di tennis, hockey su prato e di pallacanestro impiegano strategie di memorizzazione simili a quelle impiegate dai giocatori di scacchi. Dato che la caratteristica comune a entrambe gli sport è quella della presenza di due squadre che si scontrano, alcuni autori sostengono che il fattore che assimila il tipo di memorizzazione coincide sia la strategia di attacco o difesa impiegata nella partita. Per rispondere a questo interrogativo, la memoria di movimenti è stata studiata su attività sportive che non comportano la contrapposizione tra squadre. A tal proposito, uno studio importante è stato quello di Deakin e Allard, i quali hanno messo a confronto un gruppo di pattinatori su ghiaccio di prima categoria con un gruppo di pattinatori principianti. Il loro compito era quello di ricordare ( sia verbalmente che con il corpo) dei movimenti relativi al pattinaggio, dopo che questi erano stati presentati in modo corretto da professionisti. Il risultato di questo esperimento ha messo in evidenza che i pattinatori esperti hanno fornito percentuali di ricordo maggiori in entrambe le condizioni rispetto ai pattinatori non esperti. Tale risultato è stato spiegato affermando che gli esperti riescono ad accedere più facilmente alla “memoria semantica” in cui sono conservati i vari movimenti (relativi ad una disciplina sportiva) attraverso un codice di tipo proposizionale. Due studiosi, Bach e Tipper, hanno affermato che durante l’identificazione di persone tendiamo ad utilizzare il sistema motorio in quanto associamo ad esse delle specifiche abilità motorie. Per dimostrare ciò hanno fatto un esperimento in cui hanno presentato ai partecipanti le immagini di 4 atleti famosi: 2 giocatori di calcio e 2 tennisti, e – dopo- gli hanno chiesto di premere un tasto con la mano o un pedale con il piede. I risultati di quest’esperimento hanno dimostrato che le risposte date dai partecipanti erano più veloci nel momento in cui c’era congruenza tra l’immagine e l’azione; mentre erano più lente in caso di incongruenza. Ciò ha confermato l’ipotesi secondo la quale la visione di una persona che ha elevate capacità motorie influenza il sistema motorio e le azioni dell’osservatore. Negli ultimi anni le caratteristiche dei giocatori o degli atleti esperti hanno catturato l’attenzione di molti studiosi, i quali si sono concentrati su diversi aspetti: da un lato ci sono coloro i quali sostengono che i grandi esperti possiedono abilità-domini specifiche; dall’altro ci sono coloro che si sono soffermati sulle caratteristiche peculiari della memoria degli atleti e, a tal proposito, è stato suggerito che praticando – nel tempo- una disciplina sportiva, gli atleti acquisiscono automaticità nell’eseguire determinati pattern motori. Tuttavia è importante sottolineare che tali abilità non sono relative ad un singolo dominio o ad una singola disciplina, ma possono essere trasferiti da un ambito all’altro. L’ATTIVITA’ SPORTIVA: RISORSE ATTENTIVE Nell’uomo le risorse attentive disponibili sono limitate ed è per questo che nel momento in cui un soggetto compie più di un compito nello stesso tempo, possono verificarsi fenomeni di interferenza. Nell’ambito di attività agonistiche tali fenomeni possono essere provocate nel momento in cui l’atleta comincia ad elaborare delle informazioni che stanno al di fuori del campo di gara o che, comunque, non sono legate all’azione che sta svolgendo. Se ciò accade si verificherà un decadimento qualitativo della prestazione e un conseguente danno dell’attività in corso. Il consiglio che spesso viene dato agli atleti per evitare la presentazione di fenomeni di interferenza è quello di mantenere o aumentare la concentrazione; e cioè di concentrare le proprie risorse attentive sul compito rilevante in quel determinato momento e luogo. Tuttavia – per ottenere un effettivo vantaggio dalla focalizzazione delle risorse sul compito- occorre prima individuare le parti del compito stesso. Alcune caratteristiche del sistema attentivo dell’uomo sono state valutate in alcune specifiche attività agonistiche, come la pallavolo e l’atletica. Castiello e Umiltà hanno misurato lo sforzo attentivo richiesto nella pallavolo, partendo dal presupposto che maggiori sono le risorse impiegate nell’esecuzione di un compito principale, minori sono quelle disponibili per un compito secondario. Nell’esperimento i due studiosi hanno preso in analisi il momento della ricezione e gli atleti sottoposti ad esso avevano un caschetto dal quale gli è stato mandato un input sonoro in tre momenti della battuta, al quale essi dovevano rispondere con un segnale vocale. Per verificare i risultati di tale esperimento sono stati misurati i TR degli atleti sia nella condizione sperimentale (dove i compiti erano 2) sia nella condizione di controllo, in cui essi erano chiamati a svolgere un solo compito. I risultati di questo studio hanno messo in evidenza – prima di tutto- che i TR nella situazione di doppio compito rallentavano rispetto alla condizione di controllo e, in secondo luogo, che lo sforzo attentivo si modifica nel corso della prestazione per raggiungere la massima intensità nel momento in cui devono essere selezionati i programmi relativi alla risposta (nel caso della pallavolo). La stessa tecnica del doppio compito è stata adoperata anche nell’atletica e, in particolare, nelle gare dei 100 metri piani e nei 100 ad ostacoli le quali rappresentano delle attività sportive in cui la richiesta delle risorse non dipende dal comportamento dell’avversario, ma dalle difficoltà dell’attività motoria. Dai risultati emersi dagli esperimenti condotti nell’ambito di tali sport si può dedurre che la richiesta delle risorse attentive non è determinata solo dal comportamento dell’avversario ma, in primo luogo, dal compito motorio in sé e che le richieste di risorse variano durante la prestazione, aumentando nelle fasi iniziali e finali della corsa. Una delle caratteristiche principali degli atleti di varie discipline è quella di essere in grado di concentrarsi per lunghi periodi di tempo. Tale concentrazione può essere distolta da stimoli esterni o interni: gli stimoli esterni sono, per esempio, il rumore di sottofondo, il pubblico; gli stimoli interni sono la tendenza a pensare ad eventi passati o futuri, a pensare ad altre competizioni o a prestare attenzione a troppi elementi contemporaneamente. Tuttavia nella focalizzazione dell’attenzione esistono delle differenze individuali, ed è per questo che uno studioso – Mehrabian – ha distinto gli atleti in screenser e non screenser. I screenser sono meno ansiosi, più capaci di scansionare l’ambiente circostante e selezionare l’informazione di rilievo. Per i giocatori di tennis la focalizzazione dell’attenzione può migliorare attraverso una serie di strategie, come quella di imparare a concentrarsi sulla situazione presente e focalizzarsi sul compito senza lasciarsi distrarre da riflessioni sul possibile esito della propria prestazione. Gli sport all’aperto – invece- richiedono una particolare flessione nell’uso dell’attenzione perché gli stimoli presenti nell’ambiente sono moltissimi e, inoltre, i giocatori di calcio, di rugby, di pallavolo e di box devono avere un fuoco dell’attenzione più ampio in quanto devono rispondere contemporaneamente a più stimoli. Infine, i giocatori di cricket devono possedere un fuoco dell’attenzione ristretto perché in questo sport bisogna rispondere a pochi stimoli contemporaneamente. Un altro modo per misurare le risorse attentive è quello di misurare la possibilità e il tempo necessario all’orientamento dell’attenzione verso diverse posizioni spaziali. La capacità di spostare l’attenzione da un punto all’altro è una caratteristica fondamentale in molti sport e, di conseguenza, gli atleti esperti sono molto abili nell’effettuare questo tipo di processo. Per confermare questa ipotesi una serie di autori hanno sottoposto 6 arcieri, 6 pugili, 6 atleti impegnati nelle gare del pentathlon e un gruppo di controllo ad un esperimento di TR a stimoli visivi, presentati con il paradigma di Posner. Il compito dei partecipanti era quello di indirizzare la propria attenzione verso la posizione (indicata da una cifra o da un punto interrogativo) in cui sarebbe comparso lo stimolo e, all’apparire di esso, premere il pulsante di risposta. Le condizioni sperimentali che si sono presentate in questo compito furono 3: una neutra, quando lo stimolo era preceduto dal punto interrogativo; una valida, quando lo stimolo era preceduto dalla cifra che indicava la posizione corretta; una invalida, quando lo stimolo era preceduto dalla cifra che indicava la posizione non corretta. Dal confronto tra le posizioni valide e quelle invalide è risultato che si possono ottenere il beneficio o il costo temporale conseguenti alle diverse condizioni sperimentali. Inoltre i risultati di questo esperimento hanno messo in evidenza che a differenza del gruppo di controllo, i tre gruppi di atleti sembra non abbiano utilizzato l’informazione che precedeva lo stimolo per dare la risposta, non ottenendo- così- né una facilitazione temporale della condizione valida né un corrispettivo costo a seguito della condizione invalida. Questo dato è stato spiegato affermando che gli atleti esperti hanno una maggiore flessibilità nello spostamento dell’attenzione o hanno la capacità di mantenere tutto il campo visivo sotto il costante controllo dell’attenzione. Dato che questo secondo tipo di attenzione porta ad un risparmio del numero di spostamenti attenzionali da un lato, ma ad un generale ritardo delle risposte dall’altro, una serie di autori hanno affermato che i TR degli atleti esperti non sono più veloci di quelli del gruppo di controllo.