caricato da
common.user2860
Membrana Plasmatica: Struttura e Funzione

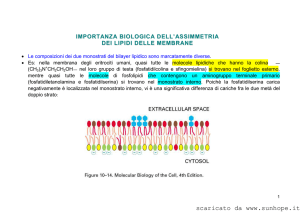
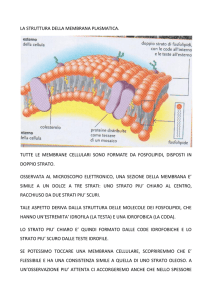
La membrana plasmatica: ASSICURA ALLA CELLULA UN IMPORTANTE ISOLAMENTO FISICO REGOLA GLI SCAMBI TRA L’AMBIENTE ESTERNO E L’AMBIENTE INTERNO DELLA CELLULA RENDE SENSIBILE LA SUPERFICIE CELLULARE ALL’ARRIVO DI DIVERSI STIMOLI RAPPRESENTA UNA BARRIERA CHE FORNISCE PUNTI DI ATTACCO AGLI ELEMENTI CITOSCHELETRICI SVOLGE UN AZIONE DI CONTENIMENTO PER LE DIVERSE COMPONENTI DEL CITOPLASMA Il modello organizzativo della membrana plasmatica venne ideato negli anni ’70 da Singer e Nicholson e prende il nome di “modello a mosaico fluido”1. Secondo questo modello, la membrana plasmatica è costituita da un doppio strato fosfolipidico (allo stato liquido-cristallino) in cui si trovano le proteine globulari di membrana. Queste proteine sono dislocate sulla superficie interna ed esterna della membrana o addirittura immerse nel doppio strato fosfolipidico. COMPOSIZIONE CHIMICA DELLA MEMBRANA PLASMATICA o I lipidi rappresentano circa il 40% del peso secco della membrana plasmatica. Tra questi vi sono fosfolipidi, glicolipidi (lipidi coniugati a catene oligosaccaridiche), colesterolo. o Le proteine rappresentano circa il 50% del peso secco della membrana plasmatica. Troviamo sia proteine semplici che proteine coniugate (prevalentemente glicoproteine). o I glucidi rappresentano circa il 10% del peso secco della membrana plasmatica. Si ritrovano prevalentemente legati a proteine (per formare glicoproteine) o legati a lipidi (per formare glicolipidi). 1 Questo modello è definito “a mosaico” proprio perché le proteine di membrana sono distribuite nel doppio strato fosfolipidico come le tessere di un mosaico. LIPIDI DELLA MEMBRANA PLASMATICA FOSFOLIPIDI sono sostanze anfipatiche, ovvero costituite da una porzione apolare e da una porzione polare; si suddividono in fosfogliceridi (o glicerofosfolipidi) e sfingofosfolipidi (o sfingolipidi). FOSFOGLICERIDI: contengono una molecola di glicerolo legata a 2 catene di acidi grassi e ad un gruppo fosfato. Il glicerolo è un alcool trivalente, quindi costituito da tre funzioni alcoliche. Due funzioni alcoliche interagiscono mediante legame estere con due catene di acidi grassi (possono essere saturi o insaturi2); la restante funzione alcolica interagisce sempre mediante legame estere con un gruppo fosfato, che a sua volta lega un gruppo polare. Il gruppo polare può essere un composto alcolico (come colina, etanolammina, glicerolo, inositolo) o un amminoacido (come serina o treonina). SFINGOFOSFOLIPIDI: contengono una molecola di sfingosina legata ad 1 catena di acido grasso e ad un ortofosfato che a sua volta lega altre molecole come la colina (in questo caso si forma la sfingomielina). Nello specifico…la sfingosina è un amminoalcol a lunga catena, contenente due gruppi chimici reattivi: un gruppo amminico e un gruppo ossidrilico. Il gruppo amminico si legherà mediante legame ammidico al gruppo carbossilico di un acido grasso, mentre il gruppo ossidrilico si legherà mediante legame estere ad un ortofosfato, che a sua volta legherà un’altra molecola (la colina nel caso della sfingomielina). 2 Gli acidi grassi saturi non presentano nella loro struttura chimica doppi legami, pertanto la loro catena carboniosa sarà lineare. Gli acidi grassi insaturi presentano nella loro struttura chimica dei doppi legami, che causeranno dei ripiegamenti nella loro catena carboniosa. GLICOLIPIDI possono contenere il glicerolo oppure la sfingosina. Un esempio di glicolipide costituito da glicerolo è il galattolipide, che contiene una molecola di glicerolo, che lega due catene di acidi grassi e una molecola di galattosio. Un esempio di glicolipide costituito da sfingosina è il cerebroside3, che contiene una molecola di sfingosina che lega una catena di acido grasso e una molecola di galattosio. COLESTEROLO è uno steroide, ovvero un lipide policiclico. Presenta un’estesa regione apolare e una piccola regione polare. La regione apolare è costituita da 4 anelli idrocarburici e da una catena laterale; mentre la piccola regione polare è costituita da un gruppo ossidrilico (si trova in corrispondenza dell’estremità della molecola). Il colesterolo è un importante elemento di stabilizzazione della membrana plasmatica (vedi dopo). Precisazione sui fosfolipidi… Come scritto sopra, i fosfolipidi (in generale) sono molecole anfipatiche, perché presentano una regione polare ed una regione apolare. La regione polare corrisponde al gruppo fosfato esterificato, mentre la regione apolare corrisponde alle due catene di acidi grassi. Proprio per queste caratteristiche, i fosfolipidi in un liquido acquoso tendono a formare spontaneamente un doppio strato nel quale le parti idrofile sono rivolte verso l’esterno e le code idrofobe verso l’interno. 3 Così chiamato perché lo ritroviamo nelle membrane plasmatiche delle cellule nervose. PROTEINE DELLA MEMBRANA PLASMATICA Le proteine globulari della membrana plasmatica si distinguono in: ESTRINSECHE contraggono rapporti superficiali con le teste idrofile dei fosfolipidi (non si affondano nel doppio strato fosfolipidico). Si distinguono in interne ed esterne a seconda se istituiscono rapporti con le teste idrofile del versante intracellulare o del versante extracellulare. INTRINSECHE una parte della loro struttura è immersa nel doppio strato fosfolipidico, mentre la restante parte protrude o dal versante intracellulare o dal versante extracellulare della membrana plasmatica e per questo classificabili in interne ed esterne. TRANSMEMBRANA attraversano da parte a parte la membrana. GLUCIDI DELLA MEMBRANA PLASMATICA Come scritto sopra, i glucidi della membrana plasmatica sono perlopiù associati a lipidi (per formare glicolipidi) e a proteine (per formare glicoproteine). Glicolipidi e glicoproteine svolgono diverse funzioni, ad esempio costituiscono il cosiddetto cell coat (o glicocalice) ovvero un rivestimento abbastanza sottile che ha il compito di proteggere dal punto di vista meccanico e chimico la membrana plasmatica. Poi, fungono da siti di riconoscimento e, infine, rivestono un ruolo molto importane nel processo di “adsorbimento” (processo che permette di trattenere adese alla superficie cellulare determinate sostanze). FLUIDITA’ DELLA MEMBRANA PLASMATICA E SUA REGOLAZIONE La membrana plasmatica presenta una certa fluidità, che è data dalle catene idro-carboniose dei lipidi. I fattori che influenzano questa fluidità sono diversi: TEMPERATURA all’aumentare della temperatura aumenta la fluidità. GRADO DI INSATURAZIONE DEGLI ACIDI GRASSI DEI FOSFOLIPIDI varia a seconda della specie lipidica considerata. Ad esempio: Fosfatidilserina e fosfatidiletanolammina sono fosfolipidi più insaturi rispetto alla sfingomielina, pertanto presenteranno catene insature, contenenti doppi legami, che ostacoleranno l’interazione tra fosfolipidi adiacenti e renderanno la membrana plasmatica più fluida. COLESTEROLO è un elemento di stabilizzazione della membrana plasmatica e lo si ritrova inframezzato tra i fosfolipidi. Ha il compito di: Diminuire la fluidità della membrana plasmatica, quando le temperature sono elevate. Questo avviene grazie al fatto che il colesterolo istituisce dei legami deboli (forze di Van der Waals, interazioni idrofobiche e ponti idrogeno) con i fosfolipidi. Aumentare la fluidità della membrana plasmatica, quando le temperature sono basse. Questo avviene grazie al fatto che il colesterolo va ad inserirsi tra i fosfolipidi ed evita che questi possano formare dei legami che renderebbero la membrana plasmatica più solida e meno fluida. ZATTERE LIPIDICHE DELLA MEMBRANA PLASMATICA Le zattere lipidiche (o raft lipidici) sono zone specializzate della membrana plasmatica che presentano grandi quantità di colesterolo e glicosfingolipidi; proprio per questo, risultano essere delle regioni a fluidità minore. È possibile distinguere due tipi di zattere lipidiche: le caveole ed i raft non invaginati. Dal punto di vista morfologico, le caveole appaiono come invaginazioni della membrana plasmatica strutturalmente stabilizzate dalla presenza di una proteina detta caveolina; mentre i raft non invaginati non sono distinguibili morfologicamente dal resto della membrana plasmatica. Dati sperimentali hanno dimostrato che la maggior parte delle proteine con funzione recettoriale sono localizzate sulle zattere lipidiche del monostrato esterno. Quest’ultime sembrano essere accoppiate alle zattere lipidiche del monostrato interno in cui sono localizzate proteine citoplasmatiche della trasduzione del segnale (ad esempio proteine G eterotrimeriche etc). Il legame funzionale tra le zattere lipidiche del monostrato esterno e le zattere lipidiche del monostrato interno permette l’interazione tra i recettori, attivati da un determinato segnale, e le proteine intracellulari. MOVIMENTI DEI FOSFOLIPIDI I fosfolipidi possono effettuare dei piccoli movimenti nell’ambito della membrana plasmatica. Si parla di: DIFFUSIONE LATERALE questo movimento permette ai fosfolipidi di uno stesso monostrato di scambiarsi di posto. ROTAZIONE movimento che permette al fosfolipide di ruotare su se stesso. DIFFUSIONE TRASVERSALE questo movimento, noto come flip-flop, permette ad un fosfolipide di passare da un monostrato ad un altro (quindi dal suo monostrato a quello controlaterale). È un processo estremamente regolato dal punto di vista biologico, avviene con dispendio energetico ed è mediato da una proteina transmembrana nota come flippasi. Quest’ultima è particolarmente presente nella membrana del REL (sede di sintesi dei fosfolipidi), mentre è scarsamente rappresentata nella membrana plasmatica. ASIMMETRIA DELLA MEMBRANA PLASMATICA La membrana plasmatica è asimmetrica, perché i due monostrati (interno ed esterno) che la costituiscono presentano notevoli differenze per quanto riguarda la componente lipidica, proteica e glucidica. Per quanto riguarda i glucidi: i glucidi nella membrana plasmatica si ritrovano perlopiù sotto forma di glicolipidi e glicoproteine e si riscontrano solo nel monostrato esterno (a contatto con l’ambiente extracellulare). Possono costituire il cosiddetto cell coat (o glicocalice) oppure svolgere funzioni recettoriali e tanto altro ancora. Per quanto riguarda i lipidi: la distribuzione dei lipidi nei due monostrati della membrana plasmatica è estremamente caratteristica. Esempio: quasi sempre è possibile trovare nel monostrato esterno fosfatidilcolina e sfingomielina, mentre nel monostrato interno fosfatidiletanolammina, fosfatidilserina e fosfatidilinositolo. fosfatidiletanolamina fosfatidilserina sfingomielina fosfatidilcolina glicolipidi Situazione particolare… La fosfatidilserina, in una cellula sana, è presente solo ed esclusivamente nel monostrato interno della membrana plasmatica. Quando una cellula avvia il processo di apoptosi (morte programmata), accade che viene attivata una flippasi specifica che trasloca la fosfatidilserina dal monostrato interno al monostrato esterno della membrana plasmatica. La presenza della fosfatidilserina sulla superficie cellulare risulta essere un segnale che induce i macrofagi, presenti nelle vicinanze, a fagocitare la cellula morta e a digerirla.