Contributi originali
Co-assunzione di alcol ed amfetamina nei ratti giovani:
effetti cognitivi e neurodegenerativi
Laura Soverchia1,2, Serena Stopponi1,2, Massimo Ubaldi1, Maria Francesca Evaristi1, Roberto Ciccocioppo1
1 Scuola di Scienze del Farmaco e dei Prodotti della Salute, Unità di Farmacologia, Università di Camerino, Italia
2 Gli autori hanno collaborato in egual misura alla realizzazione del progetto)
O
biettivi: il consumo di elevati quantitativi di bevande alcoliche in poche ore (binge drinking) associato all’uso
di psicostimolanti, rappresenta ad oggi un fenomeno di enorme diffusione fra i giovani con possibili ripercussioni a livello anatomico e funzionale in un cervello in fase di sviluppo quale quello adolescenziale. Nel presente
lavoro abbiamo utilizzato un modello animale per analizzare gli effetti neurodegenerativi e cognitivi associati alla
co-assunzione di alcol e amfetamina.
Metodi: giovani ratti Wistar sono stati intossicati per quattro giorni con alcol, amfetamina o entrambi (binge treatment). I cervelli sono stati analizzati a vari livelli mediante colorazione con Fluoro-Jade B, un marker di degenerazione neuronale. Gli effetti dell’intossicazione con alcol sulla flessibilità cognitiva sono stati studiati mediante il
test del reversal learning.
Risultati-chiave: l’intossicazione con alcol ha indotto una diffusa degenerazione neuronale nel giro dentato dell’ippocampo (DG) e nella corteccia entorinale (Etc) e perinale (PRh). Gli animali intossicati con amfetamina hanno
mostrano una compromissione dell’area somatosensoriale (S1BF), e, seppur in misura inferiore, dell’area talamica
(VA). La co-somministrazione di alcol ed amfetamina ha indotto una marcata neurodegenerazione nella Etc, nella
DG e, nel bulbo olfattorio (OB), zona debolmente lesionata nel gruppo intossicato con solo alcol. Il test del reversal learning ha evidenziato una perdita di flessibilità cognitiva dovuta all’intossicazione con alcol.
Conclusioni: il binge treatment ha indotto una diffusa neurodegenerazione a livello di alcune aree coinvolte nella
memoria quali DG ed Etc. La vasta neurodegenerazione osservata a livello del bulbo olfattorio con la presumibile
perdita di cellule dopaminergiche suggerisce la possibilità che la co-somministrazione ripetuta di alcol ed amfetamina possa portare a disturbi affettivi fra cui la depressione.
Parole chiave: ratto,
alcol, binge drinking, amfetamina, Fluoro-Jade B, neurodegenerazione
Alcohol and amphetamines co-assumption in young rats: cognitive and neurodegenerative effects
Objectives: the consumption of large amounts of alcohol in a small period of time (binge drinking) associated with the use of psycho-stimulants, is nowadays a widespread phenomenon observed among adolescents. This can lead to possible repercussions in the correct
anatomical and in the functionality of the developing brain. In this study, we used animal models to analyze the neurodegenerative and
cognitive effects associated with the consumption of alcohol and methamphetamines.
Methods: young Wistar rats were administered high amounts of alcohol or amphetamines or both for four consecutive days (binge
treatment). Brain activity was analyzed during different stages, by using Fluoro-Jade B, a marker for neural degeneration. The effects
to the cognitive flexibility due to intoxication were studied by using the reversal learning test.
Key-Results.: alcohol intoxication resulted in a high neural degeneration in the Dentate Gyrus of the Hippocampus (DG) and in
the entorhinal and perineal cortex (Etc) and (PRh). The specimens that were administered methamphetamines showed damage to the
somatosensory system (S1BF) and slight damage to the thalamus area (VA). The specimens that consumed both alcohol and methLaura Soverchia
Scuola di Scienze del Farmaco e dei Prodotti della Salute, Unità di Farmacologia, Università di Camerino
Via Madonna delle Carceri, 62032 Camerino, Italia
tel: +39 0737403322 Fax: +39 0737403325
email: [email protected]
Contatti: Dott.ssa
Di partimento Politiche Antidroga
Presidenza del Consiglio dei Ministri
15
Italian Journal on Addiction
Vol. 3 Numero 4 2013
amphetamines showed a significant neural degeneration in the Etc, DG and in the olfactory bulb, which is an area that was weakly
damaged during the only-alcohol trials. The reversal learning test showed a loss of cognitive flexibility due to alcohol intoxication.
Conclusion: the binge treatment resulted in high neural degeneration of certain areas related to memory, such as DG and Etc. The vast
level of neural degeneration detected in the olfactory bulb along with the presumable loss of dopaminergic cells, suggests that repeated
use of both alcohol and methamphetamines could be responsible for the emergence of psychiatric and neurological diseases including
depression.
Keywords: rat,
alcol, binge drinking, amphetamine, Fluoro-Jade B, neurodegeneration
Obiettivi
S
ebbene l’uso eccessivo di bevande alcoliche sia una pratica storicamente diffusa tra gli adulti, negli adolescenti sta
crescendo vertiginosamente un fenomeno noto come binge
drinking, definito come il consumo di cinque o più bevande
alcoliche in una sola occasione1,2,3.
Dai dati dell’indagine europea “Special Eurobarometer-EU
citizens’ attitudes towards alcohol”, condotta nell’ottobre
2009, risulta che un cittadino europeo su tre, di età compresa
tra 15 e 24 anni (33%) pratica il binge drinking regolarmente
almeno una volta la settimana4.
Gli adolescenti sembrano essere predisposti al binge drinking
per una alterata sensibilità agli effetti dell’alcol5,6, con un aumento della sensazione di disinibizione sociale ed una riduzione della percezione delle conseguenze negative (sintomi
dell’astinenza come ansia, tremore, crisi epilettiche) rispetto
agli adulti6,7.
Il binge drinking rappresenta un passaggio iniziale ma assai
significativo verso la progressione tossicomanica e lo sviluppo della dipendenza alcolica8,9,10 causando gravi alterazioni sia
a livello funzionale che anatomico11,12. Studi preclinici hanno
dimostrato che l’esposizione ad elevate dosi di alcol, anche
per brevi periodi può determinare gravi alterazioni neurofunzionali, morte neuronale e conclamate alterazioni morfologiche a livello delle aree corticolimbiche come l’ippocampo,
la corteccia prefrontale e le strutture cortico-amigdaloidee13,
coinvolte nel controllo dei processi emozionali, dell’apprendimento e della memoria spaziale14.
Gli psicostimolanti, fra cui le amfetamine, sono un’altra classe
di sostanze d’abuso ad elevata azione neurotossica e al contempo di ampio utilizzo nei giovani. Queste sostanze causano iperstimolazione protratta del sistema dopaminergico e
serotonergico con conseguente danno neurologico delle vie
catecolaminergiche. L’ipotesi che il loro uso possa contribuire
all’ezio-patologia dei disturbi del movimento (come ad esempio il morbo di Parkinson) è stata spesso suggerita, tuttavia
questa possibile correlazione non è stata sufficientemente
studiata15.
Nel cervello di individui che hanno abusato di amfetamine da
giovani, è stata osservata un’alterata attività elettrica a livello
della corteccia frontale e una ridotta sensibilità del sistema
dopaminergico a livello dei nuclei della base con alterazioni
dei meccanismi della memoria, capacità decisionale ed impulsività15.
Negli animali da laboratorio concentrazioni ematiche di amfetamina (AMPH) e metamfetamina (METH) comprese tra
2.5 e 3.5 μM (raggiunte dopo 1 ora dall’iniezione) causano
una deplezione del 70% della dopamina striatale, una marcata
neurodegenerazione nella corteccia e nel talamo ed un improvviso aumento della temperatura corporea fino alla morte
per ipertermia16,17,18,19.
Sebbene AMPH e METH producano chiari segni di neurotossicità negli animali adulti, meno noto è l’effetto esercitato
16
Di partimento Politiche Antidroga
Presidenza del Consiglio dei Ministri
su un cervello giovane in fase di sviluppo. Alcuni dati suggeriscono la possibilità che, anche nell’animale giovane, tali
sostanze causino compromissione della funzione delle vie
talamo-corticali20.
Un fenomeno di enorme diffusione fra i giovani è la pratica
del co-abuso, dove più sostanze psicoattive legali e/o illegali
vengono combinate per esaltarne gli effetti di rinforzo positivi ed eventualmente attenuarne gli effetti secondari sgraditi.
Una delle combinazioni più frequenti tra i giovani consumatori consiste nel mix di alcol e stimolanti del sistema nervoso21; difatti i frequenti o assidui bevitori di alcol segnalano
livelli di consumo di amfetamine o di ecstasy molto più alti
rispetto alla media della popolazione22.
Lo scopo di questa combinazione è quello di potenziare gli
effetti euforizzanti delle due sostanze e contemporaneamente
ridurne gli effetti sgradevoli, quali la sedazione indotta dall’alcol e l’agitazione e l’ansia che possono essere provocate dagli
psicostimolanti21.
Sebbene l’alcol e gli amfetaminici siano tra le sostanze d’abuso più largamente diffuse, e nonostante numerose evidenze
sperimentali indichino che queste sostanze condividono diverse proprietà farmaco-tossicologiche, non sono presenti in
letteratura studi rivolti ad un’analisi sistematica degli effetti
risultanti dal loro uso concomitante durante lo sviluppo del
sistema nervoso centrale.
Nel presente studio si sono valutati gli effetti neurotossici
derivanti dall’uso di alcool, amfetamina e la combinazione
delle stesse utilizzando giovani ratti Wistar intossicati secondo il protocollo descritto da Majchrowicz23, universalmente
accettato come modello preclinico di binge drinking13-24. Per
l’intossicazione con amfetamina è stato seguito il protocollo
sperimentale modificato di Bowyer25.
Durante la fase di intossicazione (alcolica e/o amfetaminica),
sono stati monitorati alcuni parametri fisici (temperatura corporea, stato di idratazione) e psichici (eventuali segni dell’astinenza alcolica, stato di sedazione, irritabilità) come indice
dello stato di benessere generale dell’animale.
Allo scopo di valutare se l’esposizione a dosi intossicanti di
alcol potesse avere effetti protratti sulla sfera mnesica, a distanza di alcuni giorni dalla fase di intossicazione, un nuovo
gruppo di animali è stato testato per misurare la flessibilità
cognitiva utilizzando il test del reversal learning.
Materiali e metodi
Animali
Gli animali, ratti Wistar maschi di 8 settimane, (Harlan, Correzzana, Italia) sono stati stabulati due per gabbia in condizioni di temperatura (20-22°C) e di umidità controllate (45-55%),
con un ciclo giorno/notte invertito (luce spenta alle ore 8.00).
Durante la sperimentazione, gli animali hanno avuto libero
accesso all’acqua ed al cibo (4RF18, Mucedola, Settimo Mila-
Co-assunzione di alcol ed amfetamina nei ratti giovani
nese, Italia) per l’intero arco delle 24 ore. Tutte le procedure
sperimentali sono state effettuate nel rispetto delle norme etiche sulla sperimentazione animale dell’European Community
Council Directive for Care and Use of Laboratory Animals.
Intossicazione alcolica
L’intossicazione alcolica è stata eseguita esponendo gli animali ad una dieta liquida contenente il 79.3% di una soluzione
alcolica (etanolo al 20% v/v), latte in polvere (14.7 %) (Mellin
2, Mellin S.p.A.) e zucchero (6%). I ratti di controllo hanno
ricevuto la stessa soluzione (Veicolo Alcol) priva di etanolo.
Gli animali sono stati intossicati per 4 giorni consecutivi secondo il protocollo di Majchrowicz23. La soluzione alcolica è
stata somministrata oralmente (gavage) in tre dosi frazionate
nell’arco delle 24 ore ad intervalli di 8 ore per garantire livelli
ematici alcolici continui e costanti23. Il primo giorno di intossicazione, una dose iniziale di 5 g/kg è stata somministrata
a tutti gli animali. Le dosi successive (totale 10-13 g/kg/ di
alcol al giorno) sono state stabilite valutando lo stato di intossicazione dell’animale, secondo quanto riportato nella tabella
di Majchrowicz23. Sono stati analizzati i seguenti parametri:
atassia (livello 1, 2, 3, 4), sedazione, perdita del “righting reflex” ed eventuale stato di coma. La stessa procedura è stata
eseguita nel gruppo di controllo a cui è stato somministrato
latte (veicolo dell’alcol).
I campioni sono stati centrifugati per 10 minuti a 1500 rcf,
ottenendo cosi la frazione plasmatica. La misurazione della
concentrazione di etanolo è stata effettuata con 5µl di plasma
mediante un analizzatore ad ossigeno (Analox Instruments,
Lunenburg, MA).
Peso e temperatura corporea
Gli animali sono stati pesati giornalmente per monitorare le
eventuali variazioni di peso durante le intossicazioni. La misurazione della temperatura rettale è stata effettuata un’ora
dopo la prima e la terza somministrazione giornaliera di amfetamina.
Immunoistochimica
Fase di pretrattamento. Gli animali sono stati abituati alla
procedura sperimentale ricevendo iniezioni intraperitoneali
di soluzione salina e somministrazioni orali di acqua demineralizzata nei 3 giorni antecedenti l’inizio dell’esperimento.
Fase di intossicazione. Utilizzando il disegno sperimentale
between-subject, gli animali (n=32) sono stati suddivisi in 4
gruppi (n=8/gruppo). Le somministrazioni sono state eseguite secondo lo schema riportato nelle Tabelle 1 e 2.
Per la valutazione istologica del danno neuronale, il quinto
giorno, un’ora dopo l’ultima somministrazione di alcol (ore
9.00) e di amfetamina (ore 10.00), i ratti sono stati anestetizzati con isofluorano e perfusi (perfusione intracardiaca) con
salina, seguita da una soluzione di paraformaldeide (4% paraformaldeide in 0.4 M tampone sodio fosfato pH 7.4). I cervelli sono stati rimossi e sottoposti ad un trattamento di postfissazione per 16 ore in paraformaldeide al 4%, seguita da 24
ore in saccarosio al 30%. Successivamente sono state tagliate
al criostato (Leica CM- 1950) delle sezioni orizzontali da 20
µm. Le sezioni, raccolte in accordo alle coordinate riportate
nell’atlante del cervello di ratto di Paxinos and Watson26 rispetto al Bregma, sono: -3.1, -3.2, -3.3, -3.8, -4.2, -5.1, -5.8,
-6.1, -6.2 , -6.6, -6.8, -7.1, -7.6. Le sezioni sono state montate
in vetrini Superfrost Plus® (Fisher Scientific) e conservate a
-20°C fino al momento dell’uso. Le sezioni su vetrino sono
state marcate con Fuoro-Jade B (Histo-Chem Inc. P.O.box
183, Jefferson, AR 72079, USA), marker di neurodegenerazione secondo il protocollo di Schmued27.
In dettaglio: le sezioni di cervello sono state immerse in etanolo al 100% (Sigma) per 3 minuti e reidratate attraverso passaggi graduali di soluzioni di etanolo decrescenti. Le sezioni
sono state quindi incubate in leggera agitazione con 0.06 %
di permanganato di potassio per 15 minuti, risciacquate con
acqua distillata e quindi incubate in una soluzione allo 0.01%
di Fluoro-Jade B in 0.1% di acido acetico per 30 minuti. Le
sezioni sono state successivamente lavate in acqua distillata,
disidratate, chiarificate in xilene e montate con DPX (Sigma).
I vetrini sono stati analizzati con un microscopio fluorescente
(Olympus BX51) con una luce di eccitazione blu (450-490
nm).
Livelli ematici nel sangue (BALs)
Reversal learning
I campioni di sangue (~200 µl) sono stati ottenuti mediante
prelievo dalla vena caudale degli animali appartenenti al gruppo (3) Alcol/Veicolo dell’Amfetamina e (4) Amfetamina/Alcol, 90 minuti dopo la prima somministrazione giornaliera di
etanolo nel secondo, terzo e quarto giorno di intossicazione.
Un nuovo gruppo di animali (n=20) è stato testato per stu-
Intossicazione con amfetamina
La d-amfetamina HCl (Salars, Como, Italia) è stata sciolta in
salina (sodio cloruro 0.9%, S.A.L.F. S.p.A.) e somministrata
intraperitonealmente (IP) alla dose di 3 mg/kg (volume di
somministrazione 1 ml/kg) tre volte al giorno ad intervalli
di 3 ore (ore 9.00, 12.00, 15.00). La stessa procedura è stata
eseguita nel gruppo di controllo con la sola somministrazione
di soluzione salina (veicolo dell’amfetamina).
Protocollo di intossicazione
n
Alcol
Amfetamina
Gruppo 1
8
Latte (veicolo)
Salina (veicolo)
Gruppo 2
8
Latte (veicolo)
3 mg/Kg
Gruppo 3
8
3-5 mg/Kg
Salina (veicolo)
Gruppo 4
8
3-5 mg/Kg
3 mg/Kg
Tab.1: Schema di intossicazione con alcol ed amfetamina.
Tab. 2: Schema del regime terapeutico giornaliero.
Di partimento Politiche Antidroga
Presidenza del Consiglio dei Ministri
17
Italian Journal on Addiction
Vol. 3 Numero 4 2013
diare eventuali effetti sulla flessibilità cognitiva a seguito della
intossicazione alcolica. L’analisi è stata condotta utilizzando
una procedura di reversal learning che misura la capacità di
modificare il comportamento operante per raggiungere un
determinato obiettivo28. La procedura è iniziata con una fase
di training con sessioni giornaliere di 20 minuti durante le
quali gli animali, posti in condizioni di restrizione (16 g di
cibo al giorno) hanno imparato a premere la leva attiva per
ottenere un rinforzo, che corrispondeva al rilascio di un pellet
di cibo.
Nella fase successiva il modello ha previsto l’aggiunta di uno
stimolo visivo, rappresentato da una luce posizionata sopra
ciascuna leva, utilizzata come segue:
1. leva destra associata alla illuminazione della luce destra
(RR) e viceversa (LL)
2. leva destra associata alla illuminazione della luce sinistra
(RL) e viceversa (LR)
Un computer (MED-PC software, Med associated) ha registrato sia le premute alla leva attiva che a quella inattiva. La
leva attiva è variata ogni giorno secondo lo schema Gellerman randomizzato28. Tutti gli animali sono stati allenati alle
4 differenti associazioni leva/luce fino al raggiungimento del
90 % delle risposte corrette, che consistevano nel premere la
leva attiva ottenendo il rinforzo. Terminata la fase di training,
gli animali sono stati divisi in due gruppi: a) Veicolo dell’Alcol
(n=10), b) Alcol (n=10).
Gli animali hanno ricevuto il latte (gruppo veicolo dell’alcol)
o l’alcol (gruppo alcol) secondo le modalità descritte nella
sezione Intossicazione Alcolica. Cinque giorni dopo l’ultima
somministrazione, gli animali sono stati nuovamente sottoposti ai 4 differenti programmi sopra descritti (RR, LL, RL,
LR). L’esperimento è iniziato utilizzando lo stesso programma del giorno precedente, con una sessione che è terminata
o al raggiungimento di 25 premute sulla leva attiva o dopo 5
minuti. Trascorsi 2 minuti dalla fine del programma, la sessione è ricominciata invertendo la leva attiva e mantenendo la
luce nella stessa posizione. Sono state registrate sia le risposte
corrette che incorrette.
Peso e temperatura corporea
L’analisi statistica (ANOVA) ha evidenziato l’effetto significativo del trattamento sulla variazione del peso corporeo monitorato durante la fase di intossicazione [F(3,19) = 11.25,
p<0.01] (Figura 1). L’analisi post-hoc ha rivelato una generale
diminuzione del peso corporeo nei tre gruppi di animali trattati rispetto ai controlli, evidenziando un effetto più marcato
nel gruppo intossicato con alcol ed amfetamina (riduzione
del 20%).
La somministrazione di amfetamina non ha modificato la
temperatura corporea negli animali trattati, in contrasto con
quanto indicato in letteratura, dove per dosaggi di amfetamina superiori (4 mg/kg x 5 volte al dì, ad intervalli di 2 ore)25,
si evidenzia un aumento fino a 42◦C. Nel nostro studio, nel
gruppo di animali trattati contemporaneamente con alcol ed
amfetamina, il dosaggio è stato ridotto, in quanto gli animali,
dopo uno stato iniziale di malessere generalizzato, non sono
riusciti a tollerare la combinazione. Nel gruppo di ratti trattati con sola amfetamina, solo due animali hanno raggiunto la
temperatura di 40◦C.
Livelli ematici di alcol (BALs)
I livelli di alcol nel sangue sono stati analizzati nel secondo,
terzo e quarto giorno di intossicazione nei gruppi Alcol/Veicolo ed Alcol/Amfetamina. Come illustrato in tabella 3, gli
animali mostrano un graduale aumento dei BALs durante il
trattamento, a conferma del fatto che le somministrazioni di
alcol hanno permesso il raggiungimento di livelli ematici fra i
250 e i 400 mg/dl.
Gruppo Alcol/Veicolo
Gruppo Alcol/Amfetamina
Alcol
(g/kg/giorno)
BAL
(mg/dl)
Alcol
(g/kg/giorno)
BAL
(mg/dl)
Giorno 2
13.0 ± 0.0
265.7 ±
33.8
13.0 ± 0.0
250.4 ± 30.6
Analisi dei dati
Giorno 3
10.0± 0.8
330.3 ±
33.3
9.0± 1.2
302.7 ± 29.4
L’analisi dell’andamento del peso corporeo durante i giorni
dell’intossicazione è stata eseguita mediante ANOVA a due
vie ponendo il trattamento come fattore between ed i giorni
come fattore within. La successiva analisi post-hoc è stata eseguita per mezzo del test di Dunnett. L ‘analisi dei risultati del
reversal learning è stata eseguita mediante lo Student t-test.
Giorno 4
11.8± 0.2
428.5 ±
31.3
9.8± 0.7
390.8 ± 33.4
Peso corporeo (grammi)
280
Veicolo/Veicolo
Veicolo/Amfetamina
Alcol/Veicolo
Alcol/Amfetamina
260
240
*
220
*
200
180
0
1
2
3
4
Tempo (giorni)
**
5
6
Fig. 1: Effetto delle intossicazioni alcolica e/o amfetaminica sulla variazione
di peso corporeo nei quattro gruppi sperimentali. I dati rappresentano la
media ± SEM. *p<0.05, **p<0.01 vs. il gruppo Veicolo/Veicolo
Di partimento Politiche Antidroga
Presidenza del Consiglio dei Ministri
Tab. 3: Dosi di alcol somministrate e relativi BALs per i gruppi alcol ed
alcol + amfetamina.
Intossicazione da alcol e neurodegenerazione
Peso corporeo
18
Risultati
L’analisi istochimica delle sezioni cerebrali del gruppo di animali intossicato con alcol ha mostrato un elevato numero di
cellule neuronali positive al Fluoro-Jade B (neuroni in apoptosi), evidenziabili con una colorazione verde fluorescente su
uno sfondo di verde più scuro. I risultati mostrano una marcata neurodegenerazione nelle aree del giro dentato (DG),
della corteccia entorinale (Etc), della corteccia perinale (PRh)
e della corteccia laterale entorinale (LEnt). La conta dei neuroni, effettuata ad ingrandimento 20X di tutte le sezioni cerebrali del gruppo in esame (distanza -6.1 mm dal bregma), ha
dato un valore medio di 60 cellule degenerate nell’area della
corteccia entorinale e 40 nel giro dentato dell’ippocampo. I
risultati descritti si riferiscono al conteggio di tutti i neuroni
Co-assunzione di alcol ed amfetamina nei ratti giovani
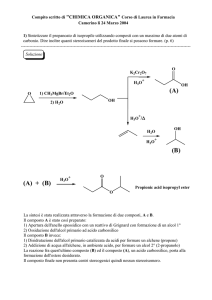
Fig. 2: Le immagini mostrano un esempio del risultato ottenuto mediante marcatura con Fluoro-Jade B delle sezioni cerebrali appartenenti al gruppo
sperimentale intossicato con alcol + amfetamina. In verde fluorescente i neuroni degenerati nel bulbo olfattorio (OB) ad ingrandimento 4X (A) e ad
ingrandimento 20X (B). Neurodegenerazione evidenziata nella regione del giro dentato (DG) ad ingrandimento 4X (C) e nelle aree della corteccia entorinale
(Ect), della corteccia perinale (PRh) e della corteccia entorinale laterale (LEnt) ad ingrandimento 4X (D). Le sezioni prese in esame si riferiscono al livello
bregma -6.1 mm
degenerati presenti sulla sezione analizzata e non è stato rapportato alle dimensioni dell’area.
Analizzando le sezioni a -5.6 mm dal bregma, una modesta
neurodegenerazione è stata evidenziata nell’area infralimbica
(IL), nella corteccia orbitale (VO) e nel bulbo olfattorio (OB).
Il quadro di neurodegenerazione è risultato identico per entrambi gli emisferi cerebrali.
Come ci si attendeva, il gruppo di animali di controllo non
ha presentato segni di neurodegenerazione sia nelle sezioni
bregma -6.1 mm che in tutte le altre aree analizzate.
Intossicazione da amfetamina e neurodegenerazione
Nei cervelli degli animali intossicati con amfetamina si evidenzia una chiara degenerazione nell’area cerebrale corrispondente alla corteccia somatosensioriale (S1BF). La media
dei neuroni marcati positivamente al Fluoro-Jade B di tutte
le sezioni cerebrali del gruppo in esame (distanza -3.1 mm
dal bregma) è di 14. Anche le aree del talamo (VA) e del periacquedotto (PAG) mostrano segni di una modesta neurodegenerazione con rispettivamente 5 e 3 neuroni marcati. Il
gruppo di animali di controllo per l’amfetamina non presenta
segni di neurodegenerazione nelle aree corrispondenti.
Intossicazione da alcol /amfetamina e neurodegenerazione
Come riportato in figura 2, nel gruppo sperimentale che ha
subito la co-somministrazione di alcol e di amfetamina, è stata osservata una chiara neurodegenerazione nelle stesse aree
risultate positive nel gruppo di ratti trattato con solo alcol,
ossia nel giro dentato (DG), nella corteccia entorinale (Etc),
nella corteccia perinale (PRh) e nella corteccia laterale entorinale (LEnt). La conta delle cellule marcate positivamente
Di partimento Politiche Antidroga
Presidenza del Consiglio dei Ministri
19
Italian Journal on Addiction
Vol. 3 Numero 4 2013
Fig. 3: Rappresentazione schematica in cui vengono riportate tutte le aree
cerebrali nelle quali è stata osservata neurodegenerazione dopo trattamento
con alcol, amfetamina o loro combinazione: bulbo olfattorio (OB), area
infralimbica (IL), corteccia orbitale (VO), corteccia somatosensoriale (S1BF),
giro dentato (DG), corteccia entorinale (Ect), corteccia perinale (PRh),
corteccia entorinale laterale (LEnt), periacquedotto (PAG)
dal Fluoro-Jade B, su tutte le sezioni del gruppo in esame, ha
definito un valore medio di 38 neuroni nell’area della corteccia entorinale e 55 nel giro dentato dell’ippocampo.
Questo risultato sembra indicare un’inversione di tendenza rispetto al gruppo di animali intossicati con solo alcol, nei quali
era maggiore il numero dei neuroni degenerati nella corteccia entorinale rispetto all’ippocampo. Il risultato più evidente
dell’effetto della co-somministrazione di alcol ed amfetamina
è la marcata neurodegenerazione a livello del bulbo olfattorio.
In questa regione una media di 30 cellule sono risultate positive al Fluoro-Jade B (Figura 2) contro la media di 3 cellule
riscontrate nell’intossicazione con solo alcol. Tutte le aree in
cui è stata osservata neurodegenerazione nei diversi gruppi
trattati sono schematizzate in Figura 3.
Reversal Learning
Durante la fase di training, tutti gli animali hanno raggiunto
il 90% di risposte corrette come previsto dal protocollo sperimentale. Terminata l’intossicazione alcolica, tutti gli animali
sono stati nuovamente esposti ai 4 differenti programmi (RR,
LL, RL, LR), raggiungendo la stessa percentuale di risposte
corrette come mostrato in Figura 4a. Nella fase di initial task,
entrambi i gruppi, controllo e ratti intossicati, hanno raggiunto la media delle percentuali di risposte corrette (80%) necessarie per raggiungere la soglia prestabilita Figura 4b. Nella
Figura 4c sono mostrate le medie percentuali delle premute
alla leva attiva nel reversal task nei due gruppi sperimentali.
I risultati mostrano che la percentuale delle risposte corrette
è stata più bassa nel gruppo degli animali intossicati rispetto
ai controlli. La differenza non è però statisticamente significativa. Questi risultati anche se preliminari suggeriscono una
20
Di partimento Politiche Antidroga
Presidenza del Consiglio dei Ministri
Figura 4. Reversal Learning Task. a) risultati ottenuti durante la fase di training
(8 giorni); viene riportata la media percentuale delle risposte rinforzanti
registrate ogni giorno. b) risultati della fase di initial task; viene riporata la
media percentuale delle risposte corrette in funzione del trattamento (veicolo
vs intossicazione alcolica). c) risultati della fase di reversal task; viene riportata
la media percentuale delle risposte corrette in funzione del trattamento
(veicolo vs intossicazione alcolica)
perdita della capacità di cambiare strategia per ottenere un
rinforzo dopo intossicazione alcolica. Ciò riflette una riduzione delle capacità cognitive conseguenti l’esposizione all’alcol.
Discussione
Per mimare la modalità di assunzione di alcolici più frequentemente adottata dai giovani è stato utilizzato il protocollo
di intossicazione sviluppato da Majchrowicz23, come modello
animale di “binge drinking”. I risultati dello studio condotto,
hanno dimostrato come la somministrazione intragastrica di
etanolo in ratti giovani, induce uno stato di costante e sostenuta intossicazione alcolica, con BALs compresi nell’intervallo
Co-assunzione di alcol ed amfetamina nei ratti giovani
250-400 mg/dl. Il livello di intossicazione è stato confermato
dall’osservazione di alcuni parametri comportamentali quali
atassia, sedazione, perdita del riflesso di equilibrio e coma.
Tra gli effetti dell’intossicazione alcolica, è stata rilevata una
marcata riduzione del peso corporeo negli animali intossicati
rispetto ai veicoli. È noto dalla letteratura che negli alcolisti,
un inadeguato apporto nutrizionale può contribuire allo sviluppo di malattie neurodegenerative celebrali29
Il test comportamentale di reversal learning ha confermato
che l’esposizione a dosi intossicanti di alcol altera la flessibilità
cognitiva degli animali, mostrando un trend alla diminuzione
della percentuale delle risposte corrette, negli animali intossicati rispetti ai veicoli.
Nei giovani, l’esposizione ad elevati quantitativi di bevande
alcoliche è caratterizzata da una maggiore suscettibilità ad
eventi neurodegenerativi, in quanto a questa età le attività
cerebrali sono fortemente plastiche e quindi sensibili alla tossicità di agenti xenobiotici30,31,32. A livello neuroanatomico, il
binge drinking è associato ad alterazioni strutturali sia dell’ippocampo33,34 che della corteccia prefrontale35,36. In accordo
con i dati di letteratura, i nostri studi, di immunoistochimica con Fluoro-Jade B, hanno evidenziato una marcata neurodegenerazione nelle aree del giro dentato, della corteccia
entorinale, della corteccia perinale e della corteccia laterale
entorinale. È stata inoltre osservata una moderata degenerazione a livello dei bulbi olfattori, della corteccia orbitale e
dell’area infralimbica. La regione ippocampale come, seppure
in misura minore, la corteccia entorinale e le aree dei bulbi
olfattori, sono conosciute per essere implicate nei processi
di apprendimento spaziale e nella memoria di lavoro37,38,39,40.
I nostri dati di neurodegenerazione suggeriscono che l’esposizione all’alcol e all’amfetamina potrebbero tradursi in una
significativa compromissione di questi processi cognitivi. In
parte questa osservazione è confermata dai dati comportamentali che, come discusso in precedenza, hanno dimostrato
che l’esposizione a dosi intossicanti di alcol riduce in maniera
significativa le capacità cognitive dell’animale.
Nei ratti giovani, il trattamento cronico con amfetamina, pur
non avendo provocato l’innalzamento della temperatura corporea rispetto ai veicoli, ha determinato una evidente degenerazione dei neuroni della corteccia somato sensoriale (S1BF).
La letteratura scientifica supporta i nostri risultati fornendo
una serie di evidenze sperimentali sul danno neurotossico
dell’amfetamina in questa area specifica41,42. D’altro canto
studi preclinici confermano che le alterazioni morfologiche
e funzionali indotte dall’ intossicazione da amfetamina nella
regione corticale somato-sensoriale non sono correlabili alla
sindrome serotoninergica e al relativo aumento della temperatura corporea16,43.
Nel nostro studio, segni di una modesta neuro degenerazione
sono stati osservati anche nelle aree del talamo (VA) e del periacquedotto (PAG). In linea con la nostra osservazione, data
precedentemente pubblicati hanno dimostrato che una singola iniezione di uno psicostimolante (cocaina o amfetamina) o
l’esposizione acuta ad uno stress sociale (social defeat stress),
determinano un aumento significativo dell’espressione del cfos, un marcatore tipico dell’attivita’ neuronale, in alcune aree
specifiche come il PAG, il locus ceruleus (LC) ed il dorsal
raphe44. La PAG potrebbe quindi essere coinvolta nella sensitizzazione da psicostimolanti (Klaus et al. 1999) e mentre
l’aumento dell’espressione del c-fos potrebbe riflettere una
attivazione neuronale associata agli effetti neurodegenerativi
osservati in questa area.
I risultati di immunoistochimica successivi alla co-somministrazione di alcol ed amfetamina hanno evidenziato una
fortissima neuro degenerazione a livello del bulbo olfattorio.
Livelli di neuro degenerazione significativi sono stati inoltre
osservati anche nel giro dentato e nella corteccia entorinale.
Il bulbo olfattorio è una regione del cervello ricca di neuroni
dopaminergici fortemente sviluppata nei roditori, coinvolta
nella regolazione dei processi olfattivi ma anche nel controllo
dell’umore. Esperimenti di lesione cerebrale hanno evidenziato, ad esempio, che nel ratto il danneggiamento dei bulbi
risulta in un marcato effetto simil-depressivo45. Queste osservazioni sono in linea con i dati clinici i quali dimostrano come
l’esposizione protratta ad amfetamina e ad altri psicostimolanti possa innescare crisi depressive nell’uomo46.
In conclusione, i dati raccolti nella nostra sperimentazione
preclinica evidenziano la pericolosità del co-abuso di alcol ed
amfetamina nell’indurre gravi danni neuronali in aree implicate nel controllo della sfera psichica e cognitiva. Questi danni
possono essere molto marcati se l’esposizione avviene in età
giovanile e cioè in una fase dello sviluppo caratterizzata da
elevata plasticità neuronale. I dati epidemiologici indicano che
l’uso degli psicostimolanti da soli e in associazione con bevande alcoliche è molto frequente proprio fra i giovani e ciò
rende la situazione particolarmente allarmante. L’attuazione
di politiche preventive e di strategie educative mirate alla riduzione dell’uso di queste sostanze dovrebbe essere percepita
come una priorità medica e sociale.
Ringraziamenti
Questo studio è stato condotto con il supporto del Fondo di
Ateneo per la Ricerca 2011 (a MU). Si ringraziano i Sign.ri
Marino Cucculelli e Rina Righi per la assistenza tecnica e la
cura degli animali.
Bibliografia
1. Miller, J. W., Naimi, T. S., Brewer, R. D., and Jones, S. E. Binge drinking
and associated health risk behaviors among high school students. Pediatrics
2007, 119: 76–85.
2. SAMHSA Substance Abuse and Mental Health Services Administration. Results from the 2007 National Survey on Drug Use and Health: National Findings. NSDUH Series H-34 2008. Rockville, MD: Office of Applied
Studies.
3. Zeigler, D.W.,Wang, C. C.,Yoast, R. A., Dickinson, B. D.,McCaffree, M.
A., Robinowitz, C. B. The neurocognitive effects of alcohol on adolescents
and college students. Prev. Med. 2005, 40: 23–32.
4. Special Eurobarometer 331-72.3. Special Eurobarometer 331. European Commission EU citizens’ attitudes towards alcohol. 2010.
5. Miller, M. W., and Spear, L. P. The alcoholism generator. Alcohol. Clin.
Exp. Res. 2006, 30: 1466–1469.
6. Spear, L. P. The adolescent brain and the college drinker: biological
basis of propensity to use and misuse alcohol. J. Stud. Alcohol. Suppl. 2002,
71–81.
7. Clark, D. B., Bukstein, O., and Cornelius, J. Alcohol use disorders in
adolescents: epidemiology, diagnosis, psychosocial interventions, and pharmacological treatment. Paediatr. Drugs 2002,4: 493–502.
8. Crews, F. T. Alcohol and neurodegeneration. CNS Drug Rev. 1999, 5:
379–394.
9. Hunt, W. A. Are binge drinkers more at risk of developing brain damage? Alcohol 1993, 10: 559–561.
10. Koob, G. F., and Le Moal, M. Drug abuse: hedonic homeostatic
dysregulation. Science 1997, 278: 52–58.
11. Tapert, S. F., Granholm, E., Leedy, N. G., and Brown, S. A. Substance
use and withdrawal: neuropsychological functioning over 8 years in youth. J.
Int. Neuropsychol. Soc. 2002, 8: 873–883.
12. Tapert SF, Schweinsburg AD, Barlett VC, Brown SA, Frank LR,
Brown GG, Meloy MJ .Blood oxygen level dependent response and spatial
working memory in adolescents with alcohol use disorders. Alcohol. Clin.
Di partimento Politiche Antidroga
Presidenza del Consiglio dei Ministri
21
Italian Journal on Addiction
Vol. 3 Numero 4 2013
Exp. Res. 2004, 28: 1577–1586.
13. Crews, F. T., Braun, C. J., Hoplight, B., Switzer, R. C. 3rd, and Knapp,
D. J. Binge ethanol consumption causes differential brain damage in young
adolescent rats compared with adult rats Alcohol Clin Exp Res. 2000,
24(11):1712-23.
14. Brown, S.A., Tapert, S.F., Granholm, E., & Delis, D.C. Neurocognitive functioning of adolescents: Effects of protracted alcohol use. Alcoholism: Clinical and Experimental Research 2000, 242:164-171.
15. Seiden LS, Sabol KE. Review. Methamphetamine and methylenedioxymethamphetamine neurotoxicity: possible mechanisms of cell destruction. NIDA Res Monogr. 1996; 163:251-76.
16. Bowyer JF, Peterson SL, Rountree RL, Tor-Agbidye J, Wang GJ.
Neuronal degeneration in rat forebrain resulting from D-amphetamineinduced convulsions is dependent on seizure severity and age. Brain Res
1998,809:77–90.
17. Bowyer JF, Ali S. High doses of methamphetamine that cause disruption of the blood– brain barrier in limbic regions produce extensive neuronal
degeneration in mouse hippocampus. Synapse 2006,60:521–32.
18. Hutchaleelaha A, Sukbuntherng J, Chow HH, Mayersohn M. Disposition kinetics of dand l-amphetamine following intravenous administration
of racemic amphetamine to rats. Drug Metab Dispos 1994a,22:406–11.
19. Hutchaleelaha A, Walters A, Chow HH, Mayersohn M. Sensitive
enantiomer-specific high-performance liquid chromatographic analysis of
methamphetamine and amphetamine from serum using precolumn fluorescent derivatization. J Chromatogr B Biomed Appl 1994b,658:103–12.
20. Adrianov O.S, Shimko I.A., Gershtein L.M., Popova N.S., Fokin V.F.
Neirofiziologicheskie I biokhimicheski mekhanizmy dozozavisimoi amfetaminovoi moduliatsii neironov sensomotornoi kory u razvivaiushchikhsia
zhivotnykh, Uspekhi Fiziologicheskikh Nauk 1997, 28: 78–97.
21. Brunell SC, Spear LP. Effect of stress on the voluntary intake of a
sweetened ethanol solution in pair-housed adolescent and adult rats. Alcohol, Clin Exp Res 2005,29:1641–53.
22. OEDT (2009), Polydrug use: patterns and responses, Questione specifica, Ufficio delle pubblicazioni dell’Unione europea, Lussemburgo.
23. Majchrowicz, E. Induction of physical dependence upon ethanol
and the associated behavioral changes in rats. Psychopharmacologia 1975,
43:245–254.
24. Schulteis G., Archer C., Tapert S.F., Frank L.R. Intermittent Binge
Alcohol Exposure during the Periadolescent Period Induces Spatial Working
Memory Deficits in Young Adult Rats. Alcohol, 2008: 42(6): 459–467.
25. Bowyer J.F. Neuronal degeneration in the limbic system of weanling
rats exposed to saline, hyperthermia or d-amphetamine. Brain Research
2000, 885:166–171.
26. Paxinos G, Watson C (1998) The Rat Brain in Stereotaxic Coordinates, 4th Edition. San Diego: Academic Press.
27. Schmued LC, Albertson C, Slikker W. Fluoro-Jade: a novel fluorochrome for the sensitive and reliable histochemical localization of neuronal
degeneration. Brain Res 1997,751(1):37-46.
28. Abdul-Monim Z, Reynolds GP, Neill JC. The atypical antipsychotic ziprasidone, but not haloperidol, improves phencyclidine-induced cognitive deficits in a reversal learning task in the rat. J Psychopharmacol
2003,17(1):57-65.
29. Crews, F.T.and Nixon Kim. Mechanisms of Neurodegeneration and
Regeneration in Alcoholism. Alcohol & Alcoholism 2009, 44(2): 115–127.
30. Nixon K, McClain JA Adolescence as a critical window for develo-
22
Di partimento Politiche Antidroga
Presidenza del Consiglio dei Ministri
ping an alcohol use disorder: current findings in neuroscience. Curr Opin
Psychiatry 2010, 23(3):227-32.
31. Spear LP. Assessment of adolescent neurotoxicity: rationale and methodological considerations. Neurotoxicol Teratol 2007, 29:1–9.
32. Nixon K, Morris SA, Liput DJ, Kelso ML. Roles of neural stem
cells and adult neurogenesis in adolescent alcohol use disorders. Alcohol.
2010,44(1):39-56.
33. De Bellis MD, Clark DB, Beers SR, Soloff PH, Boring AM, Hall J,
Kersh A, Keshavan MS. Hippocampal volume in adolescentonset alcohol
use disorders. Am J Psychiatry 2000, 157:737–744.
34. Medina KL, Schweinsburg AD, Cohen-Zion M, Nagel BJ, Tapert SF.
Effects of alcohol and combined marijuana and alcohol use during adolescence on hippocampal volume and asymmetry. Neurotoxicol Teratol 2007,
29:141–152.
35. De Bellis, M. D., Narasimhan, A., Thatcher, D. L., Keshavan, M. S.,
Soloff, P., and Clark, D. B. Prefrontal cortex, thalamus, and cerebellar volumes in adolescents and young adults with adolescent onset alcohol use
disorders and comorbid mental disorders. Alcohol. Clin. Exp. Res. 2005,
29:1590–1600.
36. Medina, K., McQueeny, T., Nagel, B., Hanson, K., Schweinsburg, A.,
and Sf, T. Prefrontal cortex volumes in adolescents with alcohol use disorders: unique gender effects. Alcohol. Clin. Exp. Res. 2008, 32:386–394.
37. Aggleton JP, Hunt PR, Rawlins JN. The effects of hippocampal lesions upon spatial and non-spatial tests of working memory. Behavioural
Brain Research 1986,19:133–146.
38. Johnson DL, Kesner RP. The effects of lesions of the entorhinal
cortex and the horizontal nucleus of the diagonal band of broca upon
performance of a spatial location recognition task. Behav Brain Res.
1994,31:61(1):1-8.
39. Nagahara AH, Otto T, Gallagher M. Entorhinal-perirhinal lesions
impair performance of rats on two versions of place learning in the Morris
water maze. Behav Neurosci. 1995,109(1):3-9.
40. Van Rijzingen IM, Gispen WH, Spruijt BM. Olfactory bulbectomy
temporarily impairs Morris maze performance: an ACTH(4-9) analog accellerates return of function. Physiol Behav. 1995,58(1):147-52.
41. Bowyer J.F., Holson R.R. Methamphetamine and amphetamine neurotoxicity: characteristics, interactions with body temperature and possible
mechanisms. Handbook of Neurotoxicology 1995, 2:845– 870.
42. Abekawa T., Ohmori T., Koyama T. Effects of repeated administration of a high dose of methamphetamine on dopamine and glutamate release in rat striatum and nucleus accumbens. Brain Res. 1994, 276–281.
43. Jakab R.L., Bowyer J.F. Parvalbumin neuron circuits and microglia in
three dopamine-poor cortical regions remain sensitive to amphetamine exposure in the absence of hyperthermia, seizure and stroke. Brain Res. 2002,
958:52–69.
44. Nikulina EM, Marchand JE, Kream RM, Miczek KA. Fos expression
in the murine brain stem after a brief social stress: behavioral sensitization to
cocaine. Brain Res. 1998, 9;810(1-2):200-10.
45. Deng X, Ladenheim B, Jayanthi S, Cadet JL. Methamphetamine administration causes death of dopaminergic neurons in the mouse olfactory
bulb. Biol Psychiatry. 2007;61(11):1235-43.
46. Negoias S, Croy I, Gerber J, Puschmann S, Petrowski K, Joraschky P,
Hummel T. Amphetamine causes dopamine depletion and cell death in the
mouse olfactory bulb Eur J Pharmacol. 2008, 589(1-3): 94–97.