Il trattamento del dolore
neuropatico in base ai
meccanismi fisiopatologici
Alberto Manassero
Brà, 22 ottobre 2005
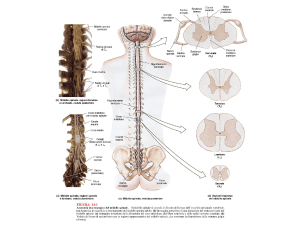
Da un punto di vista patogenetico si distingue il dolore
neuropatico in base alla sede della lesione nel sistema
nervoso:
Dolore neuropatico periferico
A) dolore da persistente ipereccitabilità dei nocicettori
B) dolore da dismielinosi
C) dolore da neuroma ovvero interruzione dei neuriti
distalmente al ganglio
D) dolore da deafferentazione ovvero interruzione dei
neuriti prossimalmente al ganglio.
Dolore neuropatico centrale
Dolore da persistente ipereccitabilità dei nocicettori.
(danno prevalentemente a carico delle fibre C)
In qualche situazione, o forse in taluni individui, lo stimolo che
dovrebbe indurre una condizione (fisiologica) di transitoria
ipereccitabilità dei nocicettori provoca invece una modifica
permanente, o comunque di durata assai più lunga del normale,
del meccanismo di trasduzione dello stimolo (sensibilizzazione
periferica). Si ha allora il permanere del dolore dopo la
guarigione del tessuto.
Questo dolore costituisce una categoria intermedia fra il dolore
tessutale e quello neuropatico.
Le fibre C sensibilizzate aumentano la loro risposta alla
stimolazione simpatica e noradrenergica (adrenosensibilità)
quindi l’aumento dei ligandi α-adrenergici nei nocicettori
collegati alle fibre C, oltre quello dei canali del Na, fa si che il
normale tono simpatico e le catecolamine circolanti partecipino
alla produzione e mantenimento del dolore aggiungendo un
meccanismo di rinforzo (SMP).
Dolore da dismielinosi
(dolore da neuropatia demielinizzante)
- danno a carico delle fibre ß e δ Se distalmente o prossimalmente al ganglio si ha un danno della
mielina senza interruzione delle fibre, si pongono le premesse
del dolore da dismielinosi, che può essere:
•da demielinizzazione
•da ipermielinizzazione
Dolore da demielinizzazione
Nei tratti di fibre nervosa demielinizzata si ha un
rallentamento della conduzione nervosa; nel tentativo di
ripristinare la normale conducibilità del nervo vengono
incorporati in queste sedi nuovi canali ionici per il Na.
Questo meccanismo di compenso, se eccessivo, fa si che la
fibra nervosa diventi patologicamente eccitabile; inoltre
poiché l’attivazione dell’assone o del dentrite avviene lungo
il suo decorso, il PTA che si produce ha modo di propagarsi
antidromicamente perché il tratto di fibra a valle non è in
periodo refrattario:
si ha quindi la riverberazione del PTA.
Dolore da ipermielinizzazione
Nei tratti di fibra ipermielinizzati, con l’aumento del
calibro dell’assone, si prolunga la durata del PTA. Se il
PTA dura più del periodo refrattario nel tratto di fibra a
valle, ha il tempo d’eccitare, oltre il tratto successivo della
fibra, anche quello che ha superato non appena questo esce
dal periodo refrattario.
Si produce quindi un extra-spike in grado di procedere
ortodromicamente e antidromicamente lungo l’assone, vale
a dire la riverberazione del PTA.
quando la riverberazione del PTA si verifica nelle fibre Aδ
si ha nocicezione patologica.
quando la riverberazione del PTA si verifica nelle fibre Aβ,
affinchè questi impulsi diventino dolorosi occorre il
reclutamento delle fibre Aδ e C.
L’attività nelle grosse afferenze mieliniche produce l’inibizione
dei terminali centrali Aδ e C secondo la teoria dell’inibizione
pre-sinaptica asso-assonica;
depolarizzazione dell’afferente primario =
inibizione pre-sinaptica
Se l’attività delle grosse afferenze mieliniche è eccessiva,
come nel caso della riverberazione del PTA, anziché il grado
di depolarizzazione sufficiente a ridurre la quantità di
neurotrasmettitore liberato dal primo neurone nocicettivo, si
ha un tale grado di depolarizzazione pre-sinaptica dei
terminali centrali Aδ e C da superare la loro soglia di
eccitazione e farli scaricare;
quello che doveva essere un meccanismo per ridurre la
nocicezione si trasforma in una causa di nocicezione
patologica.
La scarica ectopica del terminale nocicettivo così indotta
provoca da un lato la liberazione del mediatore chimico nella
sinapsi avviando un impulso nel secondo neurone e,
dall’altro, ha modo di propagarsi antidromicamente perché il
tratto di neurone che precede il terminale centrale non è in
periodo refrattario.
Si produce quindi anche nelle fibre Aδ e C la riverberazione
del PTA che amplifica e prolunga la trasmissione nocicettiva
e provoca il parossismo algico.
Dolore da neuroma
(dolore da neuropatia assonale)
- danno a carico delle fibre ß, δ e C Si ha il dolore da neuroma a seguito della sezione dei neuriti
distalmente al ganglio. La lesione nervosa include
l’interruzione di fibre afferenti sensitive di tutti i tipi (Aβ, Aδ
e C ), di fibre efferenti motorie e simpatiche.Il tentativo di
ripristinare l’integrità della fibre produce a livello del
moncone prossimale delle gemmazioni (peripheral
sprouting) che producono il neuroma d’amputazione dove
vengono incorporati i canali ionici per il sodio ed il cloro ed i
recettori adrenergici normalmente destinati all’estrema
periferia dell’assone.
Oltre all’incorporazione dei neorecettori, nel neuroma si
verifica un altro fenomeno: la produzione delle efapsi:
Le efapsi sono corto-circuiti fra fibre vicine nel contesto degli
sprouts; a causa di quelle tra le fibre afferenti integre e quelle
danneggiate in rigenerazione i neorecettori degli sprouts
possono essere indotti a scaricare dall’attività delle fibre
afferenti limitrofe ed a causa di quelle tra le fibre efferenti
simpatiche e le afferenti somatiche si ha il trasferimento
dell’efferenza simpatica sulle afferenza nocicettive somatiche
che giustifica nel dolore da neuroma il SMP.
Con la formazione del neuroma, quel che doveva essere un
processo di riparazione della fibra danneggiata si trasforma
in una causa di nocicezione ectopica.
I terminali del neuroma dove sono stati incorporati i nuovi
canali ionici diventano, infatti, dei neorecettori
spontaneamente attivi e/o ipersensibili agli stimoli
meccanici, alla distorsione della fibra, agli ioni potassio
extracellulari, all’ischemia, gli agonisti adrenergici e alla
stimolazione simpatica.
Oltre alla produzione del neuroma, a seguito dell’interruzione
delle fibre nervose “distalmente” al ganglio possono aversi
profonde modificazioni dei terminali centrali verosimilmente
secondarie ai cambiamenti dei fattori trofici trasportati
centripetamente. Queste modifiche consistono nella scomparsa
dei neuropeptidi sostanza P, somatostatina e calcitonina gene
related peptide, normalmente trasportati dalle cellule del
ganglio fino al terminale centrale.
La scomparsa dei peptidi corrisponde alla degenerazione
transganglionare dei terminali centrali delle fibre C e alla
conseguente parallela gemmazione dei terminali delle Aβ.
( central sprouting ).
Questa trasformazione della DREZ fa sì che le afferenze
tattili e propriocettive che percorrone le fibre Aβ siano in
grado di attivare i neuroni centrali che normalmente sono
attivati dalle fibre C; da questa situazione deriva l’allodinia
(secondaria).
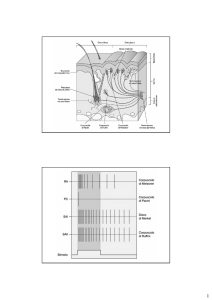
Ipereccitabilità centrale
( sensibilizzazione spinale )
Consiste in una modificazione dei neuroni nocicettivi centrali che
sviluppano condizioni di ipersensibilità quali:
•Aumentata risposta agli stimoli periferici
•Allargamento dei campi recettoriali
•Aumento della scarica spontanea
Può essere fisiologica o patologica, transitoria o permanente.
L’IPERECCITABILITA’ CENTRALE è fisiologica quando rappresenta
una normale risposta del SNC non danneggiato dalla lesione ed è
normalmente reversibile
L’IPERECCITABILITA’ CENTRALE è patologica, e potenzialmente
permanente quando, pur indotta dall’input nocicettivo, dura dopo il
suo esaurimento, diventando uno dei possibili responsabili della
mancata remissione del dolore dopo la guarigione della noxa e
quando concomita con un aumentato imput patologico
proveniente, per esempio, dai neuromi. Da questo risulta che
l’ipereccitabilità centrale può essere un meccanismo patogenetico
che si aggiunge agli altri o l’unica causa del dolore che, in questo
caso, è un’entità a sé stante che riconosce come lesione algogena
questa situazione.
L’ipereccitabilità centrale consegue all’attivazione intensa e
prolungata di fibre C nocicettive
sia di tipo ectopico
(a partenza dalla fibra)
sia di tipo normotopico (a partenza dalle terminazioni recettoriali)
Per questo motivo l’ipereccitabilità centrale si sviluppa in seguito
sia a lesioni tissutali che nervose
Neurotrasmettitori eccitatori:
Glutammato Aspartato
AMPA (ionotropo: Na-K)
Kainato (ionotropo: Na-K)
NMDA (ionotropo: Na-K-Ca)
Quisqualato-B (metabotropo)
Sostanza P
NK 1
CGRP-VIP-CCK-PG
Neurotrasmettitori inibitori:
GABA
GABA-A (ionotropo:Cl)
GABA-B (metabotropo)
Glicina - Adenosina
Oppioidi endogeni
Metabotropi (proteina G)
L’ipereccitabilità spinale
è dovuta alla ridotta inibizione dei neuroni centrali per:
- ipofunzione GABA-ergica
- deficit dei circuiti inibitori segmentali e discendenti
- attivazione dei NMDA-recettori
L’ipereccitabilità spinale può spiegare come un processo di
apprendimento mantiene per anni il dolore dopo la
guarigione della lesione.
Ridotta inibizione dei neuroni centrali per ipofunzione
GABA-ergica:
gli WDR-n contengono GABA-recettori con funzione
inibitoria. La loro attivazione è quindi frenata dal GABA per
cui soltanto gli stimoli più intensi (nocicettivi) possono
indurla. L’ipofunzione GABA-ergica conduce
all’ipereccitabilità degli WDR-n mettendoli in condizioni di
scaricare massimalmente anche all’arrivo degli impulsi nelle
fibre tattili. In queste condizioni gli WDR-n perdono la
capacità di discriminare gli stimoli nocicettivi e non
nocicettivi e producono l’allodinia Aβ-mediata.
Ridotta inibizione dei neuroni centrali per deficit dei
circuiti inibitori segmentali e/o discendenti:
Questo tipo d’ipereccitabilità centrale consegue al deficit della
funzione endorfinergica, serotoninergica e noradrenergica o al
danno anatomico del funicolo dorso-laterale.
Attivazione degli NMDA-recettori
Dolore da deafferentazione
E’ dovuto ad una lesione del primo neurone in una sede
prossimale al ganglio. Non dipende dalla trasmissione degli
impulsi nocicettivi in quanto il SNC non riceve più afferenze
dalla fibra nervosa corrispondente e l’eventuale dolore è
totalmente non nocicettivo.
Nelle lesioni di questo tipo “non si forma il neuroma” nel
moncone prossimale della fibra recisa perché il centro trofico
è distale alla lesione.
La degenerazione dei terminali centrali del primo neurone è
l’ovvia conseguenza della loro deconnessione dal centro
trofico.
La lesione nervosa nel ganglio o prossimale al ganglio
comporta la degenerazione dei terminali centrali, la
disinibizione, l’ipersensibilità da denervazione dei neuroni
centrali deafferentati ed il central sprouting dei terminali
centrali indenni.
Quel che differenzia il dolore da deafferentazione dal dolore
da neuroma è che nel primo si ha la degenerazione di tutti i
terminali centrali ( A e C ) e la deconnessione del secondo
neurone dal primo: il tentativo di riafferentazione è compiuto
dai terminali centrali dei neuroni provenienti dai metameri
vicini illesi che provano a colonizzare i secondi neuroni
deafferentati producendo l’allodinia nelle zone di confine fra
la cute ipoestesica e quella normoestesica.
Dolore centrale
Origina dall’anomala attività dei neuroni centrali per un loro
danno anatomico o una loro disfunzione primaria.
Si noti che la causa dev’essere primaria nel SNC, per cui il
dolore prodotto da una lesione nervosa periferica che ha
promosso secondariamente un meccanismo patogenetico
centrale non è centrale, anche se quel meccanismo è
d’importanza preminente.
In questo caso si ha il rinforzo del dolore da ipereccitabilità
centrale, non il dolore centrale.
La trasduzione degli stimoli nei nocicettori
Ipereccitabilità dei nocicettori
FANS/steroidi
Prostaglandine
Clonidina
Ligandi α adrenergici
Oppioidi periferici
Canali del Na
Dolore da persistente ipereccitabilità dei nocicettori
La conduzione assonale
La dismilelinosi – Il neuroma
Carbamazepina
Lidocaina
Mexiletina
Oxcarbazepina
Canali Na
Clonidina
Ligandi α adrenergici
Triciclici
Peripheral sprouting
Anestetico topico
Central sprouting
Dolore da ipereccitabilità della fibra
La trasmissione sinaptica
Gabapentin
Ca nella membrana pre-sinaptica
Pregabalin
Liberazione di glutammato e sostanza P
Oppioidi
Na-Ca tramite recettori AMPA e Neurochininici
Ketamina
Rimozione del Mg dai NMDA-recettori
Destrometorfano
Ca nella membrana post-sinaptica
Lamotrigina
Proteina kinasi-C
Topiramato
Ossido nitroso (NO)
Triciclici
Fosforilazione dei NMDA-recettori
Clonazepam
GABA sui WDR-n
Baclofene
Endorfine-serotonina-noradrenalina
Acido valproico
L’ipereccitabilità spinale
La deafferentazione
Triciclici
Disinibizione dei neuroni centrali
Clonazepam
Ipersensibilità da denervazione
Oppioidi (spinale)
Central sprouting dei terminali centrali indenni
Clonidina (spinale)
Il dolore da deafferentazione