UNIVERSITÀ POLITECNICA DELLE MARCHE
FACOLTÀ DI MEDICINA E CHIRURGIA
_______________________________________
Scuola di dottorato di ricerca in Medicina e Chirurgia
Biotecnologie Biomediche
XI ciclo nuova serie
Studio degli intermedi deamidati
del NAD e del loro metabolismo
Docente guida
Tesi di Laurea di:
Prof. Adolfo Amici
Maria Rita Carradori
A.A. 2009/10 – 2013/14
INDICE:
ABBREVIAZIONI: .............................................................................. 4
1 INTRODUZIONE.............................................................................. 5
1.1 La vitamina B3 .............................................................................. 5
1.2 Metabolismo del NAD ................................................................. 10
1.3 Via de novo ................................................................................. 11
1.4 Vie di recupero ............................................................................ 12
1.5 Nicotinamide deamidasi .............................................................. 13
2 MATERIALI E METODI ................................................................. 17
2.1 Attività enzimatica ....................................................................... 17
2.2 Saggio spettrofotometrico continuo per la determinazione
dell’attività enzimatica della carbossilesterasi ................................... 17
2.3 Saggio in HPLC per la determinazione dell’attività enzimatica della
NDasi ................................................................................................ 18
2.4 TSK-DEAE-5PW ......................................................................... 20
2.5 Elettroforesi in condizioni denaturanti su gel di poliacrilammide
(SDS-PAGE) ..................................................................................... 20
2.6 Western Blot ............................................................................... 21
2.7 Proteolisi limitata ......................................................................... 22
2.8 Clonaggio ed espressione della carbossilesterasi umana (hCE1b)
in sistemi eucariotici .......................................................................... 24
2.9 Amplificazione mediante pcr del cDNA codificante hCE1b .......... 26
2.10 Clonaggio di hCE1b in pcDNA3.1/MYC-HIS A .......................... 26
2.11 Estrazione del DNA codificante hCE1b ..................................... 28
2.12 Mutagenesi sito-specifica .......................................................... 28
2.13 Trasfezione delle cellule COS-7 ................................................ 29
2.14 La resina di affinità Ni-NTA ....................................................... 31
2.15 Immunoblotting ......................................................................... 32
2
2.16 Estrazione dei nucleotidi con etanolo bollente da pellet di cellule
Hep G2 ............................................................................................. 32
2.17 Estrazione acida dei nucleotidi da pellet di cellule HepG2 ......... 32
2.18 Preparazione degli enzimi ancillari NadD, PncC, NadE e
determinazione delle loro attività. ...................................................... 33
2.18.1 NaMN adeniltrasferasi (NadD) ............................................... 33
2.18.2 NMN deamidasi (PncC).......................................................... 35
2.18.3 NAD sintetasi (NadE) ............................................................. 37
2.19 Rilevazione e quantificazione dei mononucleotidi mediante
saggio in HPLC ................................................................................. 38
2.20 Saggio ciclico fluorimetrico ........................................................ 40
3 RISULTATI (parte I) ...................................................................... 41
3.1 CE e NDasi di fegato di coniglio e maiale ................................... 41
3.2 Determinazione della Km su RLE e PLE ..................................... 44
3.3 Mutagenesi pcDNA 3.1/hCE-1b .................................................. 49
4 RISULTATI (parte II) ..................................................................... 54
4.1 Studio sulle vie biosintetiche del NAD ......................................... 54
4.2 Conversione dell’NaMN e NMN in NaAD e quantificazione
mediante HPLC ................................................................................ 55
4.3 Conversione dell’NaAD a NAD e quantificazione mediante saggio
ciclico fluorimetrico............................................................................ 58
5 CONCLUSIONI (parte I) ................................................................ 60
6 CONCLUSIONI (parte II) ............................................................... 63
Bibliografia ...................................................................................... 65
3
ABBREVIAZIONI:
Na: Acido Nicotinico
Nam: Nicotinamide
NAD: β-Nicotinamide adenin dinucleotide
NADP: β-Nicotinamide adenine dinucleotide fosfato
NMN: Nicotinamide mononucleotide
NamR: Nicotinamide riboside
PRPP: Fosforibosil pirofosfato
QA: Acido chinolinico
NaMN: Nicotinato mononucleotide
NaAD: Nicotinato adenin dinucleotide
NamR: Nicotinamide Riboside
NDasi: Nicotinamide deamidasi
p-NPA: p-Nitrofenilacetato
p-NPN: p-Nitrofenilnicotinato
CE: Carbossilesterasi
hCE1a, hCE1b, hCE1c: Carbossilesterasi umana classe 1, isoforme 1a, 1b, 1c
hiCE: Carbossilesterasi intestinale umana
hCE2: Carbossilesterasi umana, classe 2
hCE3: Carbossilesterasi umana, classe 3 (cervello-specifica)
SDS: Sodio dodecil solfato
RLE: Carbossilesterasi di fegato di coniglio
PLE: Carbossilesterasi di fegato di maiale
4
1 INTRODUZIONE
1.1 La vitamina B3
Con il termine vitamina B3 (o niacina, o vitamina PP) ci si riferisce all’Na (acido
piridin-3-carbossilico) e alla sua forma amidata, la Nam (Fig. 1) che possiedono
attività biologica simile.
Figura 1: Struttura molecolare delle due forme della vitamina B3. Acido Nicotinico (sx) – Nicotinamide
(dx)
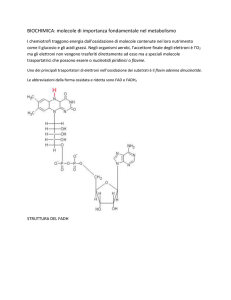
La Nam, è il precursore per il coenzima β-NAD (fig.2) e NADP, che funziona in
molte reazioni di ossidazione e riduzione catalizzate dagli enzimi. Perciò,
l’omeostasi della Nam è di vitale importanza per il corpo [1].
Figura 2: Struttura molecolare del NAD
La niacina è abbondante in molti tipi di carne, fegato, legumi, latticini, fagioli,
vegetali a foglia verde, caffè e tè. Anche il mais contiene elevate quantità di Na e
Nam, ma in forme legate e quindi non biodisponibili. Per incrementare la
5
biodisponibilità di niacina è necessario sottoporre questo cereale ad un
trattamento alcalinizzante. Tale pratica era utilizzata dalle popolazioni native del
Sud America, le quali infatti erano protette dalla deficienza da vitamina. Nella
carne, la vitamina si trova principalmente sotto forma di NAD e NADP, mentre
relativamente bassi sono i livelli di Na e Nam [2]. La Nam viene liberata dal
NAD(P) dagli enzimi della mucosa intestinale [2], mentre l’Na viene formato per
deamidazione della Nam ad opera della nicotinamidasi dei batteri presenti nel
lume intestinale [3]. Le due forme di niacina, sia introdotte tal quali, sia prodotte
nell’intestino come descritto, dopo essere state assorbite, entrano nel flusso
sanguigno per essere distribuite nei vari tessuti [2] [4] [5]. E’ stato riportato che
l’assorbimento di Nam è maggiore rispetto a quello di Na quando le fonti di
vitamina nella dieta sono il NAD e il NADP [6]. Infine, il NAD introdotto come tale
con la dieta può essere degradato nel lume intestinale a NMN e questo
defosforilato a NamR, il quale viene assorbito dalle cellule, e attraverso una serie
di pathway metabolici, viene riconvertito a NAD.
L’uomo è in grado di sintetizzare la niacina a partire dal triptofano [7]; si ritiene che
60 mg di triptofano possano dare origine a livello epatico a 1 mg di niacina (1 mg
di “equivalente di niacina” o NE) [8]. Le vitamine riboflavina (B2) e piridossina (B6),
insieme al ferro, sono necessarie per la conversione del triptofano a niacina [8] [9].
La capacità di convertire il triptofano a niacina, varia da individuo a individuo, ed è
regolata dalla dieta; l’efficienza di conversione, infatti, aumenta in condizioni di
deficienza di proteine e di triptofano, mentre diminuisce in presenza di
un’assunzione eccessiva di leucina [8]. La dose giornaliera raccomandata
espressa in equivalenti di niacina è di 16 NE per l’uomo e di 14 NE per la donna,
18 NE e 17 NE per le donne gravide e donne che allattano [10]. Nell’uomo, la Nam
e Na assunti con la dieta sono assorbiti dallo stomaco e dall’intestino sia
attraverso una diffusione passiva, sia attraverso una diffusione sodio-dipendente
[8].
Sempre nell’uomo, l’eccesso di Nam è metilato, ossidato o idrolizzato a N1metilnicotinamide o 6-idrossinicotinamide; l’N1-metilinicotinamide viene poi
ossidato a piridone N1-metil-2-piridone-5-carbossiamide (2Py) e N1-metil-4-
6
piridone-3-carbosamide dall’aldeide ossidasi (AOX; EC 1.2.3.1) e tutti questi
metaboliti vengono poi escreti con le urine [11].
La deficienza di niacina causa la pellagra, i cui sintomi principali sono
rappresentati da sfoghi cutanei foto-indotti, dermatiti, disturbi gastrointestinali e
neuropsichiatrici. Circa un centinaio di anni fa la pellagra era una malattia molto
diffusa tra la popolazione rurale povera di molti paesi, compresa l’Italia, e si
credeva fosse una malattia di tipo infettivo. Tuttavia nel 1914, Joseph Goldberger
verificò l’ipotesi per cui la pellagra potesse essere causata da una deficienza
alimentare, e scoprì che sostituendo una dieta a base di frumento con una dieta
con uova, latte e carne, la malattia veniva curata e prevenuta [12]. Ventitré anni
più tardi, Conrad Elvehjem dopo aver ottenuto da un estratto deproteinizzato di
fegato una frazione di Na e Nam, dimostrò che queste molecole avevano la
proprietà di guarire una malattia dei cani nota per la sua sintomatologia come
"black tongue" (lingua nera), conosciuta da tempo come l’equivalente della
pellagra umana [13]. I successivi studi biochimici identificarono la Nam come
componente fondamentale del NAD e del NADP e dimostrarono che gli animali
affetti da pellagra mostravano una significativa diminuzione dei livelli di NAD e
NADP nei muscoli e nel fegato [14].
Oggi la deficienza di vitamina B3 è un evento molto raro nei paesi industrializzati
[15]; tuttavia diete povere, anoressia, alcolismo, AIDS e altre malattie, possono
causare sintomi pellagra-simili. Inoltre, disturbi pellagra-simili (dermatite, disturbi
psichici) sono tipici della sindrome metabolica, nota come malattia di Hartnup,
causata da un’anomalia nel trasporto gastrointestinale e renale di alcuni
amminoacidi, tra cui il triptofano. Tali sintomi vengono alleviati e in alcuni casi
addirittura prevenuti dalla somministrazione della vitamina B3 [16].
Per molto tempo si è pensato che le manifestazioni cliniche della pellagra, fossero
dovute alla diminuzione dei livelli di NAD e NADP a concentrazioni non più
sufficienti a garantire l’energia necessaria per le varie funzioni cellulari [17]. In
effetti, queste molecole svolgono un ruolo centrale nel metabolismo cellulare di
tutti gli organismi viventi, essendo cofattori universali e ubiquitari di gran parte
delle deidrogenasi impegnate nelle vie metaboliche. Tuttavia, la comprensione
della grande varietà e complessità dei sintomi della pellagra è progredita in
7
seguito alla scoperta che la massima parte del pool cellulare del NAD, non è
utilizzato come coenzima nelle reazioni metaboliche di ossidoriduzione, ma viene
consumato dalle poli (ADP-riboso)polimerasi (PARPs) [18]. Questi enzimi
idrolizzano il legame N-glicosidico del dinucleotide, liberando la Nam e trasferendo
l’ADP-riboso
a
proteine
accettrici
di
varia
natura,
ADP-ribosilandole
e
modificandone così la funzione biologica.
La poli-ADP-ribosilazione svolge ruoli molteplici nella riparazione del DNA
danneggiato, nel mantenimento della stabilità genomica, nella regolazione
trascrizionale, nell’apoptosi e in molte altre funzioni cellulari [19]. Per quanto
riguarda ad esempio il coinvolgimento della PARP-1 nel riparo del DNA, è noto
che la sua attivazione può stimolare tre diversi pathway cellulari, a seconda
dell’intensità del danno. Nel caso in cui il danno al DNA è lieve, l’attivazione della
PARP-1 stimola il riparo del DNA e determina quindi la sopravvivenza della
cellula; se il danno non è riparabile, l’attivazione dell’enzima induce la morte
cellulare per apoptosi; infine, se il danno è severo, la PARP-1 viene iperattivata,
con conseguente diminuzione dei livelli cellulari di NAD, le cellule consumano ATP
cercando di risintetizzare il NAD e questo porta ad una significativa diminuzione
dell’energia disponibile con conseguente necrosi (fig. 3).
Figura 3: Schema del metabolismo della vitamina B3 e dell’enzima PARP-1
Studi effettuati sia in vivo che in vitro hanno dimostrato che la diminuzione dei
livelli di NAD nelle cellule determina un rallentamento dei meccanismi di poliADPribosilazione, causando così l’aumento dell’ instabilità genomica conseguente
a stress ossidativi o genotossici. La Nam assunta con la dieta è in grado di
8
rifornire le PARPs del substrato NAD, e in alcuni studi è stato osservato che
mantenere la vitamina ad un livello ottimale è indispensabile nei malati di cancro e
negli individui a rischio di esposizione ad agenti genotossici. Studi su modelli
animali hanno tuttavia dimostrato che la somministrazione di Nam può avere effetti
diversi sulla carcinogenesi a seconda del carcinogeno e dell’organo target. Ciò
probabilmente riflette la differente sensibilità di ogni organo agli agenti che
danneggiano il DNA, al fatto che la distribuzione della PARP-1 è tessuto- e cellulaspecifica, e all’osservazione che il NAD viene sintetizzato a partire da precursori
vitaminici diversi (Na, Nam, NamR) a seconda dell’organo [20].
Sia la Nam che l’Na sono proposti come agenti farmacologici nei confronti di altre
patologie: la Nam, ad esempio, ha azione protettiva nei confronti della
neurodegenerazione indotta dall’assunzione di etanolo durante lo sviluppo del feto
[21] e la somministrazione di alte dosi di Na è in grado di abbassare i livelli di
colesterolo e di trigliceridi nel sangue [22] [23].
9
1.2 Metabolismo del NAD
L’anello piridinico della Nam, introdotta con la dieta, viene incorporato nella
molecola del NAD. La Nam infatti è considerata il principale precursore del NAD
nei mammiferi [24].
Mentre nei procarioti, la molecola del NAD viene costruita anche de novo a partire
da quattro diversi composti: l’L-aspartato (L-asp), il didrossiacetone fosfato
(DHAP), il fosforibosil pirofosfato (PRPP) e l’ATP, metaboliti di pathways
essenziali quali il ciclo dell’acido citrico, la glicolisi e la via dei pentoso fosfati
(fig.4).
Figura 4: Precursori metabolici del NAD
La biosintesi del NAD(P) può avvenire sia a partire da precursori semplici (via de
novo), sia attraverso il riciclo dei prodotti di degradazione dei nucleotidi piridinici
(via di recupero, o salvage pathway) [25].
10
1.3 Via de novo
Negli eubatteri la via de novo inizia con la sintesi del QA da parte del complesso
enzimatico NadA-NadB [26]. Il primo enzima coinvolto in questo processo, l’Laspartato ossidasi (EC 1.4.3.16), codificato dal gene nadB, produce acido
imminoaspartico da L-aspartato. Questo intermedio imminico instabile, viene
convertito dalla chinolinato sintetasi (EC 2.5.1.72), il prodotto del gene nadA, in
QA (fig. 5).
Figura 5: Metabolismo del NAD in Eubatteri
Negli eucarioti, e recentemente in alcuni procarioti, si è visto come la sintesi del
QA può avvenire a partire dall’amminoacido triptofano [27], attraverso una serie di
reazioni enzimatiche, che complessivamente prende il nome di via delle
chinurenine (kynurenine pathway).
In entrambi i gruppi di organismi, il QA viene convertito a NaMN dalla chinolinato
fosforibosiltrasferasi (EC 2.4.2.19), enzima codificato dal gene nadC. Esso viene
adenilato a NaAD dalla nicotinato mononucleotide adenililtrasferasi (EC 2.7.7.18),
codificata dal gene nadD nei batteri e dagli enzimi NMN adeniltrasferasi nei
mammiferi (NMNAT, EC 2.7.7.18). Questi ultimi sono in grado di sintetizzare NAD
a partire anche dal suo immediato precursore NMN.
11
Infine, l’enzima NAD sintetasi (EC 6.3.5.1), codificato dal gene nadE nei batteri,
catalizza la reazione di amidazione del NaAD, con produzione di NAD.
Il NADP è sintetizzato tramite la fosforilazione del NAD, catalizzata dall’enzima
NAD chinasi (EC 2.7.1.23), codificato dal gene nadF nei batteri [28].
1.4 Vie di recupero
Il NAD viene degradato e riutilizzato attraverso diverse vie di recupero (PNC –
pyridine nucleotide cycle) che utlizzano come precursori alcuni derivati piridinici
preformati, tra i quali la Nam, l’Na e l’NMN.
L’Na e la Nam possono essere assimilati dall’ambiente, ma sono anche prodotti
attraverso la degradazione intracellulare del NAD. Il NAD viene degradato in
diverse forme da enzimi differenti, quali la NAD glicoidrolasi (EC 3.2.2.5), la DNA
ligasi (EC 6.5.1.1), la NAD(P) nucleosidasi (EC 3.2.2.5), la poli(ADP-riboso)
polimerasi (EC 2.4.2.30), le mono-ADP-ribosiltrasferasi (EC 2.4.2.31) e la NAD
pirofosfatasi (EC 3.6.1.22) [29]. Per i patogeni, questa via di recupero offre la
possibilità di ottenere questo cofattore direttamente dal loro ospite.
La Nam può essere riciclata a NAD attraverso l’azione dell’enzima NDasi (pncA,
EC 3.5.1.19) che converte la Nam in Na, e della nicotinato fosforibosiltrasferasi
(pncB, EC 2.4.2.11) che fosforibosila l’Na prodotto dalla prima reazione a NaMN.
L’NaMN così prodotto può essere riciclato a NAD attraverso l’attività dei due
enzimi comuni sia alla via de novo che alle vie di recupero sopra descritti: la
nicotinato mononucleotide adenililtrasferasi (NadD nei batteri, NMNAT nei
mammiferi) e la NAD sintetasi. La via biosintetica che porta alla formazione di
NAD dall’Na è detta via di Preiss-Handler. Tale via si ritrova in numerosi procarioti
ed eucarioti, inclusi i mammiferi, e ciò lascia dedurre la natura universale del
pathway metabolico (fig. 6).
12
Figura 6: Via di recupero del NAD
1.5 Nicotinamide deamidasi
Un ruolo chiave nel controllo dei livelli intracellulari di Nam può essere svolto
dall’enzima nicotinamide deamidasi (NDasi) che deamida l’anello piridinico della
Nam con formazione di Na (fig.7).
Figura 7: Reazione catalizzata dalla Nicotinamide deamidasi (NDasi)
Nei mammiferi tale attività enzimatica è stata rilevata più precisamente nell’uomo,
nel ratto e nel coniglio. La NDasi è stata purificata e parzialmente caratterizzata
dai microsomi del fegato di coniglio [30]. Si tratta di una glicoproteina contenente il
2,4% di mannosio e lo 0,57% di glucosamina; è costituita da 4 subunità di 64000
13
Da, per un peso molecolare di circa 218000 [31]. Il pH ottimale della proteina
purificata è fortemente dipendente dalla concentrazione della Nam [30]. Il valore
della Km per la Nam è compreso tra 20 e 250 mM [32]. E’ stato dimostrato che
nell’omogenato di fegato è presente un inibitore dell’attività NDasica che è stato
poi identificato come acidi grassi polinsaturi quali acido oleico, linoleico e
arachidonico, che causano la distruzione della struttura enzimatica [31]. Il
trattamento dell’enzima invece con reagenti come diisopropilfluorofosfato (DFP) e
carbobenzossiamide-2-fenietilclorometilchetone (ZPCK) che agiscono in maniera
specifica sui residui di serina e di istidina, ha messo in evidenza che questi residui
sono implicati nel meccanismo catalitico dell’enzima [33]. La NDasi, purificata da
fegato di coniglio, possiede anche attività esterasica nei confronti di composti
come il p-NPA e il p-NPN e la Km per tali composti è di qualche ordine di
grandezza più bassa di quella per la Nam [30].
Uno studio relativamente più recente ha individuato una NDasi citoplasmatica
presente nel terreno di crescita di neuroblastomi murini [34]. L’enzima è stato
purificato e caratterizzato. Le sue proprietà molecolari sono simili a quelle
dell’enzima microsomiale. Si tratta anche in questo caso di una glicoproteina con
1,7% di mannosio e 1,5% di N-acetilglucosamina, formata da 2 subunità di 65000
e 50000 Da, per un peso molecolare complessivo di 230000.
Nei batteri la NDasi è stata purificata e caratterizzata da E. Coli [35]. L’enzima ha
un peso molecolare di 30000 Da, un pH ottimale di 7,2 e una Km nei confronti
della Nam di 70 μM. Il gene pncA , che codifica per tale proteina è stato
individuato e localizzato [36]. La NDasi da lievito mostra caratteristiche simili alla
deamidasi batterica [37]. Essa, infatti, è una proteina monomerica di 34000 Da,
con un pH ottimale tra 5,5 e 7 e una Km per la nicotinamide di 33 μM. Il gene che
codifica l’enzima NDasi è stato identificato solo nei procarioti e negli eucarioti
mono e pluricellulari (Saccharomyces C. e C. Elegans). Sebbene dati
bioinformatici dimostrano che non esiste un gene omologo nei mammiferi, non si
può escludere che tale attività sia a carico di un altro enzima.
In un precedente lavoro di tesi, svolto presso il nostro laboratorio, si è dimostrato
come lo stesso enzima, purificandolo da fegato di coniglio, deamida la Nam ad
Na, ed ha attività e sequenza aminoacidica della carbossilesterasi.
14
Pertanto questa prima parte del lavoro di ricerca è volto alla parziale purificazione
e caratterizzazione dell’enzima estratto da fegato di maiale e di coniglio; valutare
se
in
entrambi
gli
organismi
le
due
attività
enzimatiche
(NDasica
e
Carbossilesterasica) sono a carico dello stesso enzima o di enzimi diversi. Una
volta stabilito che è lo stesso enzima che catalizza entrambe le reazioni
enzimatiche, si è proseguito nella seconda parte del lavoro, alla trasformazione di
cellule di mammifero contenenti il gene che codifica per la NDasi umana, volta alla
caratterizzazione dello stesso.
Le carbossilesterasi (CEs) sono proteine ubiquitarie e identificate in un ampio
numero di specie dai batteri all’uomo. Come suggerisce il loro nome, sono membri
di proteine appartenenti alla classe delle esterasi (EC 3.1.1.1 per CEs) [38] [39].
Le CEs tagliano l’estere carbossilico nel corrispondente alcool e acido carbossilico
(fig.8).
Figura 8: Reazione catalizzata dalla CE
Le CEs di mammifero sono importanti nel metabolismo e nella detossificazione di
numerosi composti endogeni e xenobiotici [40]. Esse giocano anche un ruolo
cruciale nell’attivazione dei profarmaci nell’uomo. I profarmaci contengono legami
esteri che possono aumentare la solubilità e bio-disponibilità di agenti terapeutici
[41]. Le CEs di mammifero agiscono su un’ampia varietà di esteri, amidi e tioesteri
[40] e metabolizzano numerosi composti analgesici e narcotici, inclusa l’aspirina
[42], cocaina [43], eroina [44], e la procaina [42]. L’enzima utilizza un meccanismo
di idrolisi in due step, nel quale la serina catalitica è attivata dall’attacco nucleofilo
dei residui di istidina e glutammato presenti nella triade catalitica [45]. L’acilenzima intermedio formatosi tra la serina e una porzione di substrato è rilasciato
nel secondo step della reazione o attaccando una molecola di acqua (idrolisi) o un
alcol (trans esterificazione) [45].
15
E’ stato rilevato inoltre che possono svolgere anche un’attività deamidasica nei
confronti di ammidi aromatiche.
Nell’uomo ci sono due classi di CEs, la hCE1, che è principalmente prodotta nel
fegato ma può anche essere isolata dai macrofagi e dall’ epitelio intestinale umano
[46] e la hCE2 umana, che è espressa più ampiamente nell’intestino tenue, fegato,
reni, cuore e muscolo scheletrico [47]. Una terza CE cervello-specifica, è stata
isolata nel 1999 e chiamata isoforma 3 (hCE3) [48].
Nell’uomo la proteina hCE1 è codificata dal gene CES-1, può dare origine a 3
isoforme (a,b,c) che differiscono tra loro per un amminoacido e per la loro diversa
espressione tissutale. L’enzima hCE1 è un trimero (circa 180 KDa) a
localizzazione prevalentemente epatica e si differenzia dalla forma intestinale
hCE2, codificata dal gene CES-2, con la quale condivide un 48% d’identità
aminoacidica. La proteina hCE-1b è localizzata nel reticolo endoplasmatico e il
segnale di localizzazione risiede nei 18 aminoacidi presenti all’estremità NH2terminale; mentre gli aminoacidi presenti all’estremità COOH-terminale, HIEL,
impediscono la secrezione della proteina dalla cellula. Poiché è stato dimostrato
che la proteina priva dell’estremità NH2-terminale è inattiva, è ragionevole
ipotizzare che la glicosilazione, che avviene sul reticolo endoplasmatico, è
necessaria per terminare la maturazione della proteina e renderla attiva.
L’eliminazione invece dell’estremità COOH-terminale non inattiva la proteina [49].
16
2 MATERIALI E METODI
Per dimostrare che la carbossilesterasi possiede anche attività deamidasica, dalla
Sigma, abbiamo acquistato due preparazioni enzimatiche commerciali di Rabbit
Liver Esterase (Sigma E0887) e Porcine Liver Esterase (Sigma E2884) e dissolte
in TFK 100 mM, pH 7.5 e su queste due preparazioni commerciali abbiamo poi
determinato sia l’attività carbossilesterasica, che quella deamidasica.
2.1 Attività enzimatica
L’attività enzimatica sia della CEs, che della NDasi è stata determinata
rispettivamente con il metodo spettrofotometrico in continuo e con il saggio in
HPLC discontinuo; e viene definita come la quantità di enzima che catalizza la
formazione di 1 μmole di prodotto, o la scomparsa di 1 μmole di substrato, in un
minuto nelle condizioni ottimali del saggio.
2.2 Saggio spettrofotometrico continuo per la determinazione
dell’attività enzimatica della carbossilesterasi
I metodi spettrofotometrici derivano dalla necessità di avere saggi rapidi ed
affidabili per la determinazione dell’attività in campioni provenienti da fonti diverse
e a differenti gradi di purificazione.
L’attività enzimatica della carbossilesterasi di coniglio e maiale, sono state
determinate utilizzando il p-nitrophenyl acetate (p-NPA) 0.5 mM come substrato,
seguendo l’andamento dell’assorbanza ad una lunghezza d’onda di 405 nm ed
utilizzando un coefficiente di estinzione molare di 16,4 mM-1*cm-1 per i calcoli.
Brevemente, la miscela di reazione per la misurazione dell’idrolisi del p-NPA è
costituita da una soluzione contenente: 100 mM Tris/HCl (pH 7.5), 0,4 mM p-NPA
e 0,01 mg/ml di enzima in un volume finale di 1,0 ml. L’attività enzimatica,
espressa in U/mL, è calcolata misurando l’incremento lineare di assorbanza della
miscela a 405 nm nell’unità di tempo (ΔE/ΔT) con la seguente formula:
17
ATTIVITÀ (U/mL) = (ΔE/Δt) . Vmix . 1000/ ε . Venz
dove:
ΔE = aumento dell’assorbanza a 405 nm nell’intervallo di tempo Δt;
Δt = intervallo di tempo nel quale avviene l’incremento lineare di assorbanza ΔE;
Vmix = volume totale della miscela di saggio, espressa in mL;
ε = coefficiente di estinzione millimolare del p-nitrofenolo;
Venz = volume di preparazione enzimatica nella miscela di saggio, espresso in μL.
Tutti i dati sono stati corretti sottraendo l’incremento l’assorbanza della miscela
d’incubazione senza l’enzima.
2.3 Saggio in hplc per la determinazione dell’attività enzimatica
della NDasi
L’attività della NDasi è stata determinata mediante saggio in HPLC discontinuo. La
miscela di reazione standard per la determinazione dell’attività della NDasi
contiene: 100 mM Tris/HCl (pH 7.5), 100 mM Nam, 0,1 mg/ml di enzima in 1,0 ml
finale.
La reazione inizia con l’aggiunta di substrato, mentre viene bloccata a tempi
definiti, aggiungendo un volume di HClO4 1,2 M pari ad ½ del volume della
miscela di reazione (concentrazione finale HClO4 0,4 M). Dopo 10 minuti in
ghiaccio, la miscela viene centrifugata per 3 minuti a 13000 g per allontanare le
proteine precipitate, e neutralizzata con K2CO3 0,8 M fino al raggiungimento di un
pH = 6.0. Una ulteriore centrifugazione a 13000 g x 3 minuti serve ad allontanare il
precipitato salino dal sovranatante, il quale verrà sottoposto a cromatografia a fase
inversa in HPLC. A tale scopo viene utilizzata una colonna contenente la resina
KINETEX C-18-S (Phenomenex 0,46 cm x 15 cm x 2,6 μm, 100 Å) equilibrata in
tampone formiato d’ammonio 5 mM. L’eluizione viene effettuata applicando un
gradiente continuo di acetonitrile fino al 50% nel tampone di equilibrazione. La
separazione ha una durata di 15 minuti e il flusso viene mantenuto costante a 0,75
18
mL/min. La temperatura è mantenuta costante a 18°C e la lunghezza d’onda del
rivelatore è impostata a 260 nm. Le caratteristiche del gradiente sono riportate in
Tab1:
Tempo (min)
Flusso (ml/min)
%A
%B
0
0,75
100
0
2
0,75
100
0
9
0,75
50
50
10
0,75
50
50
11
0,75
100
0
15
0,75
100
0
Tabella 1: Gradiente per la separazione dei nucleotidi piridinici mediante cromatografia a fase inversa
in HPLC. A = Tampone formiato d’ammonio 5 mM. B = acetonitrile al 50% nel tampone A.
Confrontando l’area dell’Na ottenuto iniettando in colonna il campione con l’area
del Na a concentrazione nota (standard) è possibile risalire alle μmoli di Na
iniettate. Tenendo conto delle diluizioni a cui viene sottoposta la miscela durante i
passaggi di deproteinizzazione e neutralizzazione, si risale all’attività enzimatica
espressa in Unità/mL (U/mL) utilizzando la seguente formula:
ATTIVITÀ (U/mL) = N · D · 1000 / Venz · t
dove:
N = μmoli di Na iniettate;
D = fattore di diluizione della miscela di reazione, espresso in minuti;
Venz = volume di preparazione enzimatica aggiunto alla miscela di reazione,
espresso in μL;
t = tempo di incubazione, espresso in minuti.
19
2.4 TSK-DEAE-5PW
Le preparazioni commerciali della RLE e PLE, sono state caricate su una colonna
a scambio anionico TSK-DEAE 5PW utilizzata in FPLC (dimensioni 8 mm x 75
mm), ed equilibrata con tampone 10 mM Tris/HCl pH 7.5 (tampone A). L’eluizione
è stata effettuata con gradiente lineare di NaCl da 0 M a 0,5 M in tampone A. Le
frazioni eluite sono state poi saggiate per verificare l’attività CE e NDasica. Le
caratteristiche del gradiente sono riportate in Tabella 2:
ml
%B
Flusso (ml/min)
0-5
0
(0.5)
5-60
0-100
(0.5)
60-62.5
100
(0.5)
62.5-65 100-0
(0.5)
65-67.5
(0.5)
0
Tabella 2: Caratteristiche del gradiente utilizzato per l’analisi in HPLC
Sono state raccolte frazioni di 0,5 ml e saggiate per le attività sopramenzionate.
2.5
Elettroforesi
in
condizioni
denaturanti
su
gel
di
poliacrilammide (SDS-PAGE)
La visualizzazione del contenuto proteico nelle diverse frazioni eluite dalla
purificazione è stata ottenuta mediante elettroforesi in condizioni denaturanti su
gel di poliacrilammide (SDS-PAGE) al 12% [50].
Dopo la corsa elettroforetica a 150 V per circa 1 h, il gel è stato colorato per 45
minuti con la soluzione di Comassie Brilliant Blue R-250 0,1% (W/V), acido acetico
10% (V/V) e metanolo 50% (V/V). La decolorazione è stata ottenuta mantenendo il
gel in agitazione in una soluzione di metanolo 50% (V/V) e acido acetico 10%
(V/V) fino ad ottenere l’ottimale visualizzazione delle bande proteiche.
20
2.6 Western Blot
Proteine separate mediante elettroforesi, possono essere trasferite dal gel ed
immobilizzate su una matrice, detta matrice di trasferimento.
Il trasferimento dal gel alla matrice può essere compiuto mediante semplice
diffusione, capillarità, aspirazione o elettroforesi. Il trasferimento mediante
elettroforesi è più rapido, efficiente e riproducibile quando il gel da cui trasferire le
proteine è l’acrilammide. Il tampone di trasferimento utilizzato è il Transfer Buffer
contenente 10 mM CAPS, pH= 11 (impedisce il blocco dell’N-terminale) e
Metanolo al 10%.
Dopo la corsa elettroforetica i campioni sono stati trasferiti su una membrana di
polivinildenfluoruro (PVDF, Millipore) precedentemente equilibrata per 3 secondi in
metanolo (100%), 3 minuti in acqua e 30 secondi nel tampone di trasferimento
costituito da 10 mM CAPS, pH= 11.0, metanolo 10% v/v. Il gel, equilibrato nel
tampone di trasferimento per 10 minuti, è stato messo a contatto con la membrana
PVDF procedendo all’assemblaggio dei componenti come mostrato in Fig 9:
Figura 9: Preparazione del sandwich per il trasferimento delle proteine dal gel al PVDF.
Il sandwich così composto viene immerso nel tampone di trasferimento all’interno
di una cameretta elettroforetica. Il trasferimento avviene a 4°C, in lieve agitazione,
per 3 ore a 250 mA.
21
Lo standard di proteine per la determinazione dei pesi molecolari trasferito su
membrana di PVDF è stato tagliato e colorato con una soluzione costituita da
metanolo 50%, acido acetico 10% e Coomassie Blue Brilliant R-250 0.7%, e
successivamente posta in una soluzione decolorante costituita da metanolo 50% e
acido acetico 10%. Il resto della membrana invece, dopo un abbondante lavaggio
in tampone 10 mM Tris, pH= 7.5, 150 mM NaCl (TBS), è stato incubato per tutta la
notte a 4°C in tampone composto da TBS e albumina sierica bovina (BSA) per
bloccare i siti idrofobici della membrana di PVDF rimasti liberi dopo il
trasferimento. Dopo un esteso lavaggio con tampone 20 mM Tris, pH= 7.5, 500
mM NaCl, Tween 20 0.05% (v/v), Triton X-100 0.2% (v/v) (TBS-Tween/Triton) la
membrana è stata incubata con l’anticorpo da coniglio primario con anti-His alla
diluizione di 1:1000 a temperatura ambiente per un’ora. Previo lavaggio con TBSTween/Triton e poi TBS, la membrana è stata infine incubata con l’anticorpo
secondario da capra coniugato con perossidasi di rafano, diluito 1:1000 nella
soluzione di TBS-Buffer addizionata di latte liofilizzato 10% per 1 ora a
temperatura ambiente. I segnali sono stati rilevati mediante chemioluminescenza
attraverso il software Quantity One (ChemiDocTM XRS, Biorad).
2.7 Proteolisi limitata
La proteolisi limitata è responsabile dell’attivazione di un’ampia gamma di proteine
immature ed è quindi implicata in un numero di sistemi biologicamente importanti
[51] [52] [53].
Basti pensare alla conversione del tripsinogeno in tripsina che avvia l’attivazione di
tutti gli altri enzimi precursori del succo pancreatico.
Le proteasi sono state ampiamente utilizzate in molte aree della biochimica, nel
sequenziamento, nell’(in) attivazione enzimatica, nella degradazione [54] [55].
Tutte queste reazioni idrolitiche, si verificano in condizioni denaturanti così che il
peptide verrà tagliato, lungo la sequenza aminoacidica locale, in corrispondenza di
quelle sequenze che soddisfano le caratteristiche della proteasi in questione [56].
Questa applicazione è utile per la determinazione della sequenza aminoacidica, in
22
caso di digestione completa i cui frammenti possono poi essere analizzati con la
spettrofotometria di massa.
L’utilizzo della proteolisi in condizioni denaturanti preclude però ogni possibilità di
ottenere informazioni sulla struttura tridimensionale delle proteine. Per questo tipo
di studi è necessario operare in condizioni native, inoltre le condizioni di reazione
per la proteolisi limitata sono scelte in modo tale da garantire che la proteina non
venga degradata completamente. Per la digestione sono utilizzati vari metodi.
In genere il rapporto tra proteasi e proteina si aggira tra 1:50 e 1:100 in questo
modo la proteolisi è incompleta e gli intermedi possono essere osservati man
mano che vengono formati.
Molte delle reazioni di proteolisi, sono osservate attraverso SDS-PAGE,
comunque, l’elettroforesi di proteine digerite, raramente permette di identificare in
maniera precisa il sito di taglio; per determinare il punto esatto dove la proteasi
taglia la proteina è buona norma effettuare una degradazione di Edman [57] [58].
Il sito iniziale di taglio, può alterare la stabilità della proteina, la quale
successivamente si sfolda, esponendo tutto il peptide alla completa degradazione
da parte della proteasi.
Nella proteolisi limitata, i siti tagliati dalle proteasi sono quelli più superficiali, quindi
esposti al solvente, come i loop o i segmenti di connessione tra domini.
Questa assunzione, confermata da osservazioni sperimentali, è stata ampiamente
utilizzata per studiare l’accessibilità dei siti, nelle proteine sconosciute.
Affinché la proteolisi abbia luogo è richiesto un unfolding locale della proteina.
Modelli plausibili derivati sulla base di strutture note proteolizzate hanno mostrato
che è necessario che almeno un tratto di 10 aminoacidi lungo la catena
polipeptidica dia luogo ad unfolding locale.
In maniera simile è stato dimostrato che i foglietti β sono inadatti ad essere tagliati
dalla proteasi. Nell’α-elica invece, sono presenti minori restrizioni geometriche al
local unfolding, sebbene sia probabile che le costrizioni energetiche sfavoriscano il
taglio all’interno dell’elica [57].
Il sito in cui taglia la proteasi, si è visto che forma dei deboli legami idrogeno con il
resto della proteina. Per cui il sito più suscettibile al taglio è stato caratterizzato
23
essere quello che instaura le più deboli interazioni con il resto della proteina e
perciò è la zona più favorevole a svolgersi.
2.8 Clonaggio ed espressione della carbossilesterasi umana
(hCE1b) in sistemi eucariotici
Il vettore plasmidico usato per l’espressione in cellule di mammifero è il
pcDNA3.1/myc-His A 5.5 kb della INVITROGEN (V855-20) (fig. 10).
In questo vettore, l’espressione della proteina ricombinante è sotto il controllo del
Figura 10: Mappa del vettore d’espressione pcDNA3.1/myc-His A
promotore del citomegalovirus, localizzato a monte di un polilinker costituito da
diversi siti di restrizione e dentro il quale viene inserito il gene che deve essere
clonato.
In
questo
modo
la
proteina
viene
prodotta
fusa
all’estremità
carbossiterminale con l’epitopo myc, che ne permette la rivelazione in
immunoblotting, e con una coda di 6 residui di Istidina (6 x His-Tag) che ne
permette, invece, la purificazione mediante resina di affinità Ni-NTA. Il vettore
contiene anche un cDNA che codifica per il gene che fornisce resistenza
all’ampicillina, il quale può essere usato per la selezione delle cellule trasfettate.
24
I costrutti ricombinanti vengono propagati e selezionati in cellule di E.Coli TOP 10
F’ fatte crescere in presenza di ampicillina. Il plasmide infatti presenta il gene
AmpR che codifica per l’enzima -lattamasi che idrolizza l’ampicillina.
Il vettore è in grado, inoltre, di replicare ad alto numero di copie, grazie alla
presenza dell’origine di replicazione SV40, che viene stimolata dalla porzione
large dell’antigene T di SV40, espresso in determinati tipi di cellule di mammifero
quali, ad esempio, le COS-7 ( fibroblasti renali di scimmia, fig.11).
Figura 11:Cellule COS-7 fibroblasti di Cercopithecus aethiops ( monkey, African green)
Da glicerinati di Top10 F’ contenenti il pcDNA3.1 che possiede la sequenza della
carbossilesterasi umana hCES1b è stato estratto il DNA plasmidico con il kit:
NucleoSpin Plasmid (MN); con questo kit, il pellet batterico, contenente il DNA
plasmidico viene risospeso in un buffer di lisi alcalino in modo da liberare il DNA
plasmidico dalle cellule ospiti di E.coli. Un’ulteriore buffer neutralizza il lisato e
crea le condizioni appropriate per il legame del DNA plasmidico alla membrana del
NucleoSpin Plasmid QuickPure Column. Le proteine precipitate, il DNA genomico,
ed i detriti cellulari, sono pellettati mediante centrifugazione. Il DNA plasmidico
puro viene in fine eluito con un buffer contenente 5 mM Tris/HCl, pH= 8.5.
Una volta estratto il DNA plasmidico, questo dapprima viene tagliato con l’enzima
di restrizione EcoRI al fine di linearizzare il plasmide, successivamente viene
tagliato con altri due enzimi di restrizione (EcoRI e XhoI) per vedere se è presente
l’inserto. Gli enzimi utilizzati sono entrambi della Fermentas.
25
2.9 Amplificazione mediante pcr del cDNA codificante hCE1b
La reazione di PCR è stata condotta nel tampone addizionato di MgCl 2 2 mM,
dNTP 200 mM, dell’opportuna coppia di primers 0,3 μM (le sequenze dei primers
sono riportate in tabella 3), delle opportune quantità di DNA stampo (plasmide
pcHUMCAR) e di Pfu-polimerasi High Fidelity (Biotools) secondo le indicazioni
della casa produttrice. Alla miscela di reazione è stato aggiunto anche DMSO 10%
a causa dell’elevato contenuto in GC del templato. L’amplificazione è stata
affettuata in thermal cycler (GeneAmp PCR system 2400 Perkin Elmer Life
sciences), impostando 35 cicli termici e una temperatura di annealing di 50°C.
Gli amplificati sono stati caricati su un gel d’agarosio 1% e purificati con il kit High
pure Product
PCR Purification Kit (Roche) secondo le istruzioni della ditta
produttrice.
Enzima di restrizione
associato
Fw (5’
3’)
5’-CCTAGAATTCATGTGGCTCCGTGCCTTTATCCTGGC-3’
EcoRI_FW
Rev (5’ 3’)
5’-TTTGCTCGAGTTATTCTGTCTGGGGTGGCT-3’
XhoI_REV
Tabella 3: Sequenze dei primers Fw e Rev usati per il clonaggio in pcDNA
2.10 Clonaggio di hCE1b in PCDNA3.1/MYC-HIS A
I siti di restrizione inseriti nella coppia di primers utilizzata per l’amplificazione del
cDNA hanno permesso il clonaggio direzionale nel vettore d’espressione
pcDNA3.1/myc-His A.
L’inserto e il vettore sono stati digeriti con gli enzimi di restrizione negli opportuni
tamponi (Fermentas) secondo le indicazioni fornite dalla ditta produttrice. Dopo
ogni digestione preparativa, inserto e vettore sono stati caricati su gel di agarosio
1% e purificati con il kit di purificazione High Pure Product PCR Purification Kit
(Roche).
26
Vettore ed inserto, preparati come descritto e recanti le opportune estremità sticky
complementari, sono stati tra loro saldati mediante T4 DNA ligasi (FERMENTAS)
secondo le istruzioni della ditta produttrice ed utilizzando nella miscela di ligazione
2 U di enzima, un rapporto molare vettore:inserto 1:5 ed incubazione a 22°C per 1
ora. L’intera miscela di ligazione (20 μl) è stata utilizzata per trasformare cellule di
E.coli TOP10 F’. Brevemente, il protocollo di trasformazione prevede, dopo
l’aggiunta della miscela di ligazione ad un’ aliquota di 200 μl di cellule competenti,
l’incubazione in ghiaccio per 30 minuti seguita da shock termico a 37°C per 5
minuti. L’aliquota di cellule, previa aggiunta di 500 μl di terreno LB (Luria-Bertani;
triptone 1%, estratto di lievito 0,5%, NaCl 1%, pH= 7.0), è stata incubata per 40
minuti a 37°C in agitazione. La miscela è stata piastrata su piastre di LB-agar
(terreno LB addizionato di agar alla concentrazione di 15 g/l) addizionate di
ampicillina ad una concentrazione finale di 100 μg/ml. Le piastre sono state
lasciate in incubazione a 37°C overnight e le colonie cresciute sono state saggiate
per la presenza dei plasmidi ricombinanti mediante colony-PCR e caricamento
degli amplificati su gel d’agarosio 1%. Una generica miscela di saggio per colonyPCR contiene dNTP 200 μM, l’opportuna coppia di primers specifici per il cDNA
d’interesse (0,5 μM ciascuno), il templato rappresentato da una porzione di colonia
prelevata con un puntale sterile, Taq-polimerasi (Biotools) 1U e il relativo tampone
in presenza di MgCl2 1,5 mM. Le colonie positive alla colony-PCR vengono fatte
crescere in terreno LB addizionato di ampicillina alla concentrazione finale di 100
μg/ml. I plasmidi ricombinati vengono estratti dalle colture con il kit Nucleo Spin
Plasmid (Macherey-Nagel) secondo le istruzioni della ditta produttrice ed in
seguito digeriti con gli opportuni enzimi di restrizione per verificare il rilascio
dell’inserto. Il corretto inserimento e l’assenza di mutazioni nell’inserto viene infine
stabilito mediante sequenziamento nucleotidico.
27
2.11 Estrazione del DNA codificante hCE1b
Da glicerinati di Top10 F’ contenenti il pcDNA3.1 che possiede la sequenza della
carbossilesterasi umana hCE1b è stato estratto il DNA plasmidico con il kit:
NucleoSpin Plasmid (MN); con questo kit, il pellet batterico, contenente il DNA
plasmidico viene risospeso in un buffer di lisi alcalino in modo da liberare il DNA
plasmidico dalle cellule ospiti di E.coli. Un’ulteriore buffer neutralizza il lisato e
crea le condizioni appropriate per il legame del DNA plasmidico alla membrana del
NucleoSpin Plasmid QuickPure Column. Le proteine precipitate, il DNA genomico,
ed i detriti cellulari, sono pellettati mediante centrifugazione. Il DNA plasmidico
puro viene in fine eluito con un buffer contenente 5 mM Tris/HCl, pH= 8.5.
Una volta estratto il DNA plasmidico, questo dapprima viene tagliato con l’enzima
di restrizione EcoRI al fine di linearizzare il plasmide, successivamente viene
tagliato con altri due enzimi di restrizione (EcoRI e XhoI) per vedere se è presente
l’inserto. Gli enzimi utilizzati sono entrambi della Fermentas.
2.12 Mutagenesi sito-specifica
La mutagenesi sito-specifica è stata effettuata sul vettore di espressione pcDNA
3.1/myc-His A(Invitrogen) contenente l’intero cDNA codificante per la hCE1b. In
particolare è stato mutato il nucleotide in posizione 1522 presente sul cDNA
codificante per la hCE1b utilizzando il kit “QuickChange XL II site-directed
mutagenesis kit” (Stratagene), seguendo il protocollo fornito dalla ditta produttrice.
È stata eseguita una reazione di mutagenesi utilizzando la seguente coppia di
primers:
Fw (5’ 3’)
5’-GGAACTGGGGTCACCTGGACCAGGTGG-3’ hCE1_MUT_FW
Rev (5’ 3’)
5’-CCACCTGGTCCAGGTGACCCCAGTTCC-3’ hCE1_MUT_REV
Tabella 4: Sequenze dei primers Fw e Rev usati per la mutagenesi
28
La presenza effettiva della mutazione nel plasmide risultante è stata verificata
attraverso il sequenziamento del vettore.
2.13 Trasfezione delle cellule COS-7
Il costrutto ricombinante pcDNA-hCE1b viene inserito nelle cellule di mammifero
COS-7 per l’espressione della proteina ricombinante, attraverso la tecnica della
trasfezione.
A seconda del destino a cui il DNA inserito va incontro si può distinguere tra
trasfezione stabile e trasfezione transiente.
La trasfezione è stabile quando il DNA esogeno si integra stabilmente nel genoma
cellulare e viene quindi trasmesso anche alle cellule figlie. La selezione delle
cellule che hanno ricevuto il DNA esogeno avviene ponendo le stesse in un mezzo
selettivo per circa 2-3 settimane, fino alla selezione delle singole colonie che
vengono poi propagate nel normale terreno di crescita. La trasfezione è transiente
quando il vettore resta nella cellula come elemento extracromosomico, cioè non
integrato nel genoma cellulare. In questo caso l’espressione della proteina ha una
durata limitata nel tempo. Le cellule infatti, dopo 48-72 ore, vengono raccolte e
lisate. L’espressione della proteina ricombinante nella frazione solubile dei lisati
cellulari è valutata mediante immunoblotting e medinate saggi di attività
enzimatica. La trasfezione del costrutto ricombinante in cellule COS-7 è stata da
noi effettuata in maniera transiente mediante l’impiego di Lipofectamina
(Invitrogen). La lipofectamina è una formulazione di liposomi cationica, la quale
forma dei complessi con le cariche negative delle molecole degli acidi nucleici per
permettergli di attraversare la membrana cellulare (fig. 12). Il DNA contenente i
liposomi (con la carica positiva sulla loro superficie) possono fondersi la carica
negativa della membrana plasmatica delle cellule vive, e permettere l’introduzione
del DNA all’interno della cellula.
29
Figura 12: Trasfezione transiente mediante l'utilizzo di liposomi
Il terreno utilizzato per la crescita è il D-MEM (Dulbecco’s modified Eagle’s
medium) High glucose 4,5 g/l, supplementato con L-glutammina 4 mM,
anfotericina B [250 μg/ml], penicillina-streptomicina [10,000 U/ml penicillina –
10,000 μg/ml streptomicina] e FBS 10% (siero bovino fetale). La riproduzione delle
cellule avviene in incubatore a 37°C, in atmosfera controllata di CO2 5%. Le cellule
sono state fatte crescere nel terreno di crescita, in fiasche con area di 75 cm2 , fino
a raggiungere una confluenza dell’80%. Il plasmide è stato prorogato in cellule di
E.coli e successivamente estratto con kit endotoxin-free Pure Link HiPure Plasmid
Filter Purification Kit (Invitrogen). Alle cellule nei pozzetti è stata aggiunta una
soluzione DNA-Lipofectamina, in terreno OPTIMEM (Gibco) nel rapporto 1:2,5 e
lasciata per 2 ore a 37°C con 5% di CO2. Il terreno OPTIMEM è stato sostituito
con il terreno di crescita delle COS-7 e le cellule sono state incubate per altre 48
ore. Successivamente sono state staccate con tripsina, risospese nel terreno DMEM e centrifugate a 1000 g per 5 minuti a 4°C. Il pellet cellulare così ottenuto è
stato lavato con PBS, quindi le cellule sono state rotte per sonicazione nel
tampone di lisi così costituito:
100 mM Tris/HCl pH= 7.5
Il sonicato è stato centrifugato a 13000 rpm per 5 minuti e il supernatante ottenuto
è stato utilizzato per la determinazione della concentrazione proteica, la
determinazione dell’attività carbossilesterasica, purificazione dell’estratto grezzo
mediante cromatografia di affinità Ni-NTA e per esperimenti di immunoblotting al
fine di verificare l’espressione della proteina ricombinante.
30
2.14 La resina di affinità Ni-NTA
L’espressione e la purificazione di proteine ricombinanti facilita la produzione e la
caratterizzazione dettagliata di teoricamente tutte le proteine. Sebbene una
grande varietà di sistemi di espressione eterologhi siano stati sviluppati nel corso
degli anni e sono ancora oggi usati per produrre proteine ricombinanti, la
purificazione delle proteine di interesse può essere ancora uno step problematico.
Le procedure di purificazione classiche possono essere sempre impiegate, ma in
molti casi le tecniche del DNA ricombinante hanno permesso la costruzione di
proteine di fusione in cui specifici tag di affinità sono aggiunti alla sequenza
proteica d’interesse; l’utilizzo di questi tag di affinità semplifica la purificazione
delle proteine di fusione ricombinanti, attraverso l’utilizzo di metodi cromatografici
di affinità.
La resina Ni-NTA agarose è composta da acido nitrilotriacetico (NTA)
immobilizzato su una matrice di Sepharose CL-6B. Il gruppo funzionale della
resina è l’acido nitrilotriacetico, un chelante di metalli tetradentato che occupa
quattro dei sei siti di legame della sfera di coordinazione dello ione nickel; gli altri
due siti possono interagire con gli anelli imidazolici dei 6 residui istidinici
consecutivi presenti nella coda con cui viene funzionalizzata la proteina
ricombinante. Questa coda (6xHis tag) può essere posta sia all’N- che al Cterminale della proteina di interesse. La resina Ni-NTA lega saldamente le proteine
con code istidiniche permettendo la purificazione ad omogeneità anche di proteine
ricombinanti che costituiscono una bassissima percentuale (<<1%) del contenuto
proteico totale [59]. L’eluizione viene condotta utilizzando concentrazioni elevate di
imidazolo o istidina che competono con i residui di istidina per il legame alla
resina, oppure attraverso la protonazione, abbassamento dal pH, dei residui di
istidina che vengono respinti dalla carica positiva dello ione metallico. La Ni-NTA
agarose offre una elevata capacità di legame (5-10 mg di proteina con 6 residui di
istidina all’estremità di fusione per millilitro di resina) con interazioni non
aspecifiche molto ridotte.
31
2.15 Immunoblotting
Allo scopo di visualizzare la proteina hCE1b ricombinante, espressa in cellule
COS-7, sono stati messi a punto degli esperimenti di immunoblotting. Pertanto
un’opportuna quantità di estratto grezzo delle cellule trasfettate è stata caricata su
un gel SDS-PAGE di Tricina al 10%, quindi trasferita su membrana di PVDF. La
membrana è stata successivamente incubata con l’anticorpo primario anti-myc e
lasciata a 4°C overnight. L’incubazione con l’anticorpo secondario anti-mouse IgGHRP e la rivelazione, effettuata in chemiluminescenza, sono state condotte
utilizzando il kit Super Signal West Femto Maximum Sensivity Substrate (Pierce),
secondo le istruzioni della ditta produttrice.
2.16 Estrazione dei nucleotidi con etanolo bollente da pellet di
cellule Hep G2
I pellet vengono risospesi in 600 μl di tampone Hepes 10 mM, 75% etanolo, pH
7.1 bollente e poi incubati a 100°C per 3 minuti. Dopo centrifugazione a 13000
rpm per 10 minuti, i campioni sono stati portati a secco tramite la speed-vac e
risospesi in 600 μl di acqua [60].
2.17 Estrazione acida dei nucleotidi da pellet di cellule HepG2
I pellet vengono risospesi in 550 μl di HClO4 0,4 M. Dopo 10 minuti in ghiaccio, le
cellule vengono centrifugate a 13000 rpm per 10 minuti e il sovranatante è stato
neutralizzato con K2CO3 1 M fino al raggiungimento di un pH pari a 6.0.
Un’ulteriore centrifugazione a 13000 rpm per un minuto serve poi a separare il
precipitato salino dal sovranatante.
32
2.18 Preparazione degli enzimi ancillari NadD, PncC, NadE e
determinazione delle loro attività.
2.18.1 NaMN adeniltrasferasi (NadD)
L’enzima ancillare NaMN adeniltrasferasi (NadD) è stato purificato da pellet di
cellule E.coli BL21 precedentemente trasformate con il plasmide pOC29
contenente il gene corrispondente. Il pellet è stato risospeso in un tampone di lisi
(Tris-HCl 20 mM, pH 8.0, MgCl2 1 mM, EDTA 0,5 mM, PMSF 2 mM) in un volume
pari a 1/20 del volume originale della coltura. Dopo rottura meccanica delle cellule
per sonicazione (3 cicli da 1 minuto con pause da 1 minuto), la sospensione
cellulare è stata centrifugata a 13000 rpm per 10 minuti. Il sovranatante
rappresenta l’estratto grezzo. Dopo aver determinato la concentrazione proteica
dell’estratto grezzo attraverso il metodo Bradford [61], è stata determinata l’attività
enzimatica mediante un saggio spettrofotometrico continuo.
L’estratto grezzo è stato caricato su una colonnina contenente la resina Ni-NTA,
collegata ad un sistema FPLC (AKTA, Biotech Pharmacia). Tale resina viene
utilizzata nella cromatografia per chelatura di metalli, che sfrutta la capacità di certi
ioni metallici, specialmente Ni2+, Cu2+, Zn2+, Hg2+ e Cd2+, di legare proteine
formando complessi di coordinazione con i gruppi imidazolici dell’istidina, i gruppi
indolici del triptofano o i gruppi tiolici di residui di cisteina.
La resina Ni-NTA agarose utilizza lo ione Ni2+ ed è composta da acido nitrilacetico
(NTA) immobilizzato su una matrice di SEPHAROSE CL-6B. Il gruppo funzionale
della resina è proprio l’NTA che grazie alle sue proprietà di chelante dei metalli,
occupa quattro siti di legame della sfera di coordinazione dello ione Ni 2+ presente
nella resina. I restanti due siti di coordinazione restano quindi liberi per
l’interazione con gli anelli imidazolici dei residui istidinici presenti sulle proteine
ricombinanti. La resina è stata equilibrata in un tampone Tris-HCl 50 mM pH 7.5,
NaCl 0,15 M (tampone A), Imidazolo 10 mM, ad un flusso costante di 1 ml/min.
Dopo un lavaggio con lo stesso tampone di equilibrazione (10 volumi di colonna),
l’eluizione è stata eseguita in 45 minuti applicando un gradiente discontinuo di
33
imidazolo da 10 mM a 350 mM, nel tampone A. Le caratteristiche del gradiente
sono riportate in Tabella 5:
Tempo (min) % (A + Imidazolo 10mM) % (A+ Imidazolo 350mM) Durata (min)
0,0
10,0
20,0
45,0
100
94
0
0
6
100
1,0
15,0
END
Tabella 5: Caratteristiche del gradiente utilizzato per la cromatografia in FPLC
L’eluato è stato raccolto in frazioni che sono state saggiate per verificare la
presenza dell’attività enzimatica. Le frazioni attive sono state riunite in un unico
pool conservato a -20°C.
Il saggio spettrofotometrico continuo per la determinazione dell’attività enzimatica
prevede l’allestimento di una miscela di reazione in cui sono presenti sia il
campione di cui si vuole saggiare l’attività NaMN adeniltrasferasica, sia tutti gli
enzimi coinvolti nella formazione del NADH a partire dal NaAD.
Figura 13: Il NADH viene sintetizzato a partire dall’NaMN prodotto nella miscela di reazione in seguito
ad una serie di reazioni ancillari catalizzate dagli enzimi NadD, NadE e alcol deidrogenasi
In particolare, un’opportuna quantità di campione viene aggiunto ad una miscela
costituita da Hepes 56 mM pH 7.5, etanolo assoluto 0,5% v/v, semicarbazide 14
mM, MgCl2 11 mM, NH4Cl 4,5 mM, ATP 1 mM, NaMN 0,1 mM, ADH 0,03 mg/mL,
BSA 0,56 mg/mL e NadE (45 μg di proteine di un estratto grezzo di E. coli BL21
che iperesprimono l’enzima ricombinante NadE).
La velocità di formazione del NaAD, direttamente proporzionale alla velocità di
formazione del NADH, viene determinata andando a misurare nel tempo
l’aumento di assorbanza a 340 nm, alla temperatura di 37°C. L’attività enzimatica
(U/mL) è calcolata sulla base dell’aumento lineare di assorbanza a 340 nm della
miscela nel tempo (ΔE/Δt) rispetto ad un bianco preparato con la miscela di saggio
sopra descritta nella quale al posto del campione viene aggiunto un identico
34
volume di acqua. Infatti, nel calcolo dell’attività enzimatica è necessario tenere in
considerazione
l’aumento
di
assorbanza
causato
dall’attività
NaMN
adeniltrasferasica endogena, presente negli estratti ricombinanti di E.coli.
L’attività enzimatica, espressa in U/mL, viene calcolata con la formula:
Ae (U/mL) = (ΔE/Δt) ∙ Vmix ∙ 1000/ ε ∙ Venz
Dove:
ΔE = aumento dell’assorbanza a 340 nm nell’intervallo di tempo Δt
Δt = intervallo di tempo nel quale avviene l’incremento lineare di assorbanza ΔE
Vmix = volume totale della miscela di saggio, espressa in mL
ε = coefficiente di estinzione millimolare del NADH (6,2 mM-1 ∙ cm-1)
Venz = volume di preparazione enzimatica nella miscela di saggio, espresso in μL.
2.18.2 NMN deamidasi (PncC)
L’espressione della proteina ricombinante PncC è stata ottenuta facendo crescere
il clone di E. coli proveniente dalla libreria ASKA [62]iperesprimente la proteina di
interesse. Le cellule sono state fatte crescere a 37°C in agitazione nel terreno
Luria Bertani addizionato di Cloramfenicolo 34 μg/ml fino ad una OD600 pari a 0.4,
dopodiché le colture sono state incubate a 20°C e addizionate di IPTG 1 mM per
indurre l’espressione delle proteine. Dopo 12 ore dall’induzione, le cellule sono
state raccolte mediante centrifugazione a 5000 g per 10 minuti e i pellet congelati
a -20°C. Per la preparazione degli estratti grezzi, i pellet cellulari sono stati
opportunamente risospesi in tampone di lisi (Tris-HCl 50 mM, NaCl 0,15 M, PMSF
1 mM, DTT 1 mM e 2 μg/ml di ciascuno dei seguenti inibitori di proteasi:
chimostatina, antipaina, leupeptina, pepstatina) in un volume pari a 1/20 del
volume originale della coltura. Dopo rottura meccanica delle cellule per
sonicazione (3 cicli da 1 minuto con pause da 1 minuto), le sospensioni cellulari
sono state centrifugate a 13000 g per 10 minuti. I sovranatanti, che rappresentano
gli estratti grezzi, sono stati sottoposti ad analisi elettroforetica per verificare
l’avvenuta espressione delle proteine.
35
Anche per la determinazione dell’attività di questo enzima si è utilizzato il metodo
spettrofotometrico continuo utilizzato per NadD. La quantità di NaMN formata
viene determinata misurando allo spettrofotometro la formazione di NADH in
seguito a una serie di reazioni ancillari che prevedono l’utilizzo degli enzimi NadD,
NadE e alcol deidrogenasi.
Figura 14: Il NADH viene sintetizzato a partire dall’NMN prodotto nella miscela di reazione in seguito
ad una serie di reazioni ancillari catalizzate dagli enzimi PncC, NadD, NadE e alcol deidrogenasi
Come per la precedente attività, un’opportuna quantità della preparazione
enzimatica viene aggiunta ad una miscela costituita da Hepes 56 mM pH 7.5,
etanolo assoluto 0,5% v/v, semicarbazide 14 mM, MgCl2 11 mM, NH4Cl 4,5 mM,
ATP 1 mM, NMN 1 mM, ADH 0,03 mg/mL, BSA 0,56 mg/mL, NadD (45 μg di
proteine di un estratto grezzo di E.coli BL21 che iperesprimono l’enzima
ricombinante NadD), NadE (45 μg di proteine di un estratto grezzo di E. coli BL21
che iperesprimono l’enzima ricombinante NadE). La velocità di formazione
dell’NaMN, direttamente proporzionale alla velocità di formazione del NADH, viene
determinata andando a misurare nel tempo l’aumento di assorbanza a 340 nm,
alla temperatura di 37°C. L’attività enzimatica (U/mL) è calcolata sulla base
dell’aumento lineare di assorbanza a 340 nm della miscela nel tempo (ΔE/Δt)
rispetto ad un bianco preparato con la miscela di saggio sopra descritta nella
quale al posto della preparazione enzimatica viene aggiunto un identico volume di
acqua. Infatti, nel calcolo dell’attività enzimatica è necessario tenere in
considerazione l’aumento di assorbanza causato dall’attività NMN deamidasica
endogena, presente negli estratti ricombinanti di E.coli.
Per il calcolo dell’attività enzimatica, viene utilizzata la stessa formula descritta nel
paragrafo precedente.
36
2.18.3 NAD sintetasi (NadE)
Glicerinati di cellule di E.coli BL21 trasformate con i costrutti d’interesse vengono
inoculati in terreno liquido LB addizionato di ampicillina (concentrazione finale 0,1
mg/ml). Dopo crescita overnight a 37°C, le precolture vengono inoculate in terreno
LB addizionato di ampicillina in modo tale da ottenere una OD600 compresa tra
0,05 e 0,1. Le colture vengono cresciute a 37°C fino al raggiungimento di una
OD600 compresa tra 0,5 e 0,8; vengono quindi poste a 25°C e dopo 30 minuti, si
procede
all’induzione
dell’espressione
delle
proteine
aggiungendo
IPTG
(concentrazione finale 0,4mM). La crescita viene condotta per tutta la notte a
25°C. Le cellule sono quindi raccolte mediante centrifugazione a 5000 g per 10
minuti e i pellet vengono conservati a -20°C.
Il pellet viene risospeso nello stesso tampone di lisi usato per la purificazione di
PncC e NadD. Dopo sonicazione e centrifugazione, l’estratto grezzo viene caricato
su una colonna contenente 1 ml di resina Ni-NTA equilibrata con Tris-HCl 50 mM,
pH 7.5, NaCl 0,15 M, imidazolo 10 mM. Dopo un lavaggio con Tris-HCl 50 mM, pH
7.5, NaCl 0,15 M, imidazolo 40 mM, la proteina viene eluita con Tris-HCl 50 mM,
pH 7,5, NaCl 0,15 M, imidazolo 250 mM. Il pool costituito dalle frazioni attive viene
sottoposto a gel filtrazione su colonna contenente la resina PD10, equilibrata ed
eluita con HEPES/KOH 50 mM, pH 7.5, NaCl 0,3 M. Anche in questo caso, ad
ogni passaggio delle procedure di purificazione descritte vengono determinate la
concentrazione proteica, con il metodo Bradford [61]e l’attività enzimatica
mediante il saggio spettrofotometrico continuo.
Come per le precedenti attività, un’opportuna quantità della preparazione
enzimatica viene aggiunta ad una miscela costituita da Hepes 56 mM pH 7.5,
etanolo assoluto 0,5% v/v, semicarbazide 14 mM, MgCl2 11 mM, NH4Cl 4,5 mM,
ATP 1 mM, NaAD 1 mM, e ADH. Anche in questo caso andiamo a valutare il
NADH che si forma dal NAD andando a misurare nel tempo l’aumento di
assorbanza a 340 nm, alla temperatura di 37°C. L’attività enzimatica (U/mL) è
calcolata sulla base dell’aumento lineare di assorbanza a 340 nm della miscela nel
tempo (ΔE/Δt) rispetto ad un bianco preparato con la miscela di saggio sopra
descritta nella quale al posto della preparazione enzimatica viene aggiunto un
37
identico volume di acqua. Infatti, nel calcolo dell’attività enzimatica è necessario
tenere in considerazione l’aumento di assorbanza causato dall’attività endogena,
presente negli estratti ricombinanti di E.coli.
2.19 Rilevazione e quantificazione dei mononucleotidi mediante
saggio in HPLC
I livelli intracellulari dei mononucleotidi sono stati determinati mediante la loro
conversione a NaAD, sfruttando l’attività degli enzimi NMN deamidasi (PncC) e
NaMN adeniltrasferasi (NadD): il primo è responsabile della trasformazione di
NMN a NaMN, il secondo della conversione di NaMN a NaAD. A tale scopo 100 μl
degli estratti nucleotidici preparati con l’estrazione acida sono stati addizionati a
tre miscele di reazione (mix 1, 2, 3) contenente in 150 μl finali tampone fosfato di
potassio 100 mM pH 8.0, NaF 10 mM, MgCl2 1 mM, ATP 1 mM, 4 mU di PncC e 4
mU di NadD ricombinanti purificati. In una prima miscela l’estratto nucleotidico è
incubato in assenza degli enzimi ancillari e questo ci darà la quantità del NaAD
endogeno; nella seconda miscela l’estratto è incubato in presenza di NadD e il
NaAD rivelato in questa miscela corrisponderà al NaAD endogeno più quello
derivante dall’NaMN. Nella terza miscela l’estratto è incubato con entrambi gli
enzimi ancillari, NadD e PncC e qui il NaAD sarà la somma del NaAD endogeno,
quello formato dal NaMN e quello formato dall’NMN. Dopo 20 minuti di
incubazione a 37°C, le reazioni vengono bloccate aggiungendo un volume di
HClO4 1,2 M pari a ½ della miscela di reazione (concentrazione finale HClO 4 0,4
M). Dopo 10 minuti in ghiaccio, le miscele acidificate vengono centrifugate per 5
minuti a 13000 rpm (per allontanare le proteine precipitate) e neutralizzate con
K2CO3 0,8 M fino al raggiungimento di un pH pari a 6.0. Un’ulteriore
centrifugazione a 13000 rpm per un minuto serve poi a separare i precipitati salini
dai sovranatanti che sono stati direttamente sottoposti a cromatografia in HPLC.
Per la separazione cromatografica è stata utilizzata una colonna contenente la
resina Supelcosil LC-18-S (25 cm x 4 mm), equilibrata in tampone A (fosfato di
potassio 0,1 M pH 6.0). L’eluizione è stata effettuata applicando un gradiente
38
discontinuo di metanolo nel tampone A. Il gradiente ha una durata di 35 minuti e il
flusso viene mantenuto a 1,3 ml/min. La temperatura alla quale sono state
effettuate le corse è 18°C. Il sistema HPLC utilizzato è dotato di un rilevatore
diode-array che consente l’analisi del campione nell’intervallo di lunghezze d’onda
compreso tra 200 e 350 nm. Le caratteristiche del gradiente, realizzato
mescolando in proporzioni diverse il tampone A con il tampone B (tampone A
contenente il 20% di metanolo) sono riportate in Tabella 6:
Tempo (min)
%A
%B
Durata (min)
0
100
0
-
9,0
88
12
6,0
15,0
55
45
2,5
17,5
0
100
2,5
25,5
100
0
5,0
35,0
END
Tabella 6: Caratteristiche del gradiente utilizzato per l'analisi in HPLC. per durata s'intende l'intervallo
di tempo che il sistema impiega a raggiungere la composizione della fase mobile indicata nella
seconda e terza colonna, a partire dal tempo della corsa cromatografica indicato nella prima colonna.
Confrontando l’area del picco dell’ NaAD del campione con l’area del dinucleotide
a concentrazione nota (standard), è possibile risalire alle μmoli di NaAD formate
nella miscela di reazione. Tenendo conto delle diluizioni a cui viene sottoposta la
miscela durante i passaggi di deproteinizzazione e neutralizzazione, si risale
all’attività enzimatica espressa in U/mg utilizzando la seguente formula:
Ae (U/mg) = N ∙ D / mg proteine ∙ T
Dove:
N = μmoli di NaAD iniettate
D = fattore di diluizione della miscela di reazione
T = tempo d’incubazione della miscela di reazione, espresso in minuti
mg proteine = quantità di proteine presenti nel campione, espresse in mg.
39
2.20 Saggio ciclico fluorimetrico
Al fine di aumentare sia la sensibilità sia la velocità di esecuzione del saggio, il
NaAD che si forma a seguito delle reazioni enzimatiche precedentemente
descritte, è dapprima convertito a NAD tramite l’enzima NadE, e poi rilevato
mediante un saggio ciclico fluorimetrico sviluppato da Graeff and Lee [63].
Brevemente la procedura prevede l’allestimento di tre miscele di reazione (75 μL
finali) costituite da estratto nucleotidico del nostro campione, tampone fosfato di
potassio 100 mM pH 6.5, NaF 10 mM, MgCl2 1 mM, ATP 2,5 mM, 4 mU di PncC
ricombinante che trasforma l’NMN in NaMN e 4 mU di enzima NadD ricombinante
che trasforma l’NaMN in NaAD, Hepes 56mM, pH7.5, etanolo assoluto 0,5% (v/v),
semicarbazide 14 mM, MgCl2 11 mM ed un eccesso di ADH da S. cerevisiae tale
da ridurre tutto il NAD endogeno a NADH. Le miscele di reazione sono incubate a
37°C per 10 minuti e successivamente addizionate con 25 μL di HCl 0,8 M che
oltre a stoppare le reazioni, distrugge il NADH, sia quello endogeno sia quello
prodotto
dall’ADH,
che
altrimenti
contribuirebbe
alla
fluorescenza.
Successivamente, le miscele sono neutralizzate con 25 μL di NaOH 0,8 M per
raggiungere
un pH pari a 6.0. A 125 μL di queste miscele, si aggiunge un
tampone costituito da Hepes 56 mM pH 7.5, etanolo assoluto 0,5% v/v,
semicarbazide 14 mM, MgCl2 11 mM, NH4Cl 4,5 mM e l’enzima NAD sintetasi
(NadE) al fine di ottenere una stechiometrica conversione di tutto il NaAD a NAD.
Viene poi aggiunto il reagente ciclico che contiene: tampone fosfato di sodio 100
mM pH 8.0, etanolo 2%, resazurina 30 μM, FMN 10 μM, ADH da S. cerevisiae
0,1mg/ml in BSA 0,1 mg/ml, diaphorase 0,01 mg/ml. Durante il ciclo si ha la
conversione della resazurina nel composto altamente fluorescente resorufina. La
reazione di fluorescenza è stata monitorata in un lettore di piastre con eccitazione
a 530 nm ed emissione a 590 nm.
40
3 RISULTATI (parte I)
3.1 CE e NDasi di fegato di coniglio e maiale
In questa prima parte del lavoro di tesi, abbiamo acquistato due preparazioni
commerciali di CE provenienti da fegato di coniglio e maiale; e su questi due
enzimi, abbiamo testato sia l’attività CE, che NDasica. Per verificare se ambedue
le attività sono a carico dello stesso enzima abbiamo proceduto con prove
cromatografiche, in modo osservare la coeluizione o separazione delle due
attività.
Per la determinazione di entrambe le attività enzimatiche, sono stati allestiti due
saggi già descritti nella sezione 2.2, 2.3.
Da quest’analisi sono emersi i seguenti risultati riportati in tabella:
ESTERASI DA FEGATO DI CONIGLIO
(0,83 mg/ml)
Nam deamidase
6,6 mU/ml
7,9 mU/mg
Carbossilesterase 32,21 U/ml
38,8 U/mg
Entrambe le attività sono effettivamente presenti nella preparazione commerciale
con rapporto carbossilesterasi/deamidasi stimato pari a ~5000.
Con una separazione elettroforetica mediante SDS-PAGE si è osservato come la
preparazione commerciale non fosse omogenea elettroforeticamente (fig.15); per
tale motivo si è proceduto con una cromatografia a scambio anionico per
osservare la coeluizione o separazione delle due attività.
RABBIT
liver (3ug)
RABBIT
liver (12ug)
STD
97,4 KDa
66,2 KDa
60,0 KDa
45,0 KDa
31 KDa
14,4 KDa
Figura 15: SDS PAGE 12% della preparazione commerciale della RLE
41
Dalla cromatografia a scambio anionico, abbiamo raccolto tutte le frazioni eluite, e
su ognuna è stata determinata sia l’attività CE che NDasica.
Il grafico che segue, mostra come la preparazione commerciale della esterasi da
fegato di coniglio, eluisca in un unico picco. Il grosso picco alla fine del gradiente è
presente anche nel bianco e quindi ininfluente.
mAu
λ=280
λ=260
200
180
160
140
120
100
80
60
40
20
0
0
5
10 15 20 25 30 35 40 45 50 55 60 65 70
ml
Figura 16: Profilo di assorbanza della cromatografia TSK-DEAE
Entrambe le attività coeluiscono nel picco di assorbanza osservato nel profilo.
Mentre nelle altre frazioni non è statto osservato nulla.
Effettuando un SDS-PAGE delle frazioni raccolte nel picco attivo, si osserva come
la preparazione commerciale sia elettroforeticamente omogenea.
STD
97,4 KDa
66,2 KDa
45,0 KDa
31 KDa
14,4 KDa
Figura 17: SDS PAGE 12% del picco attivo
42
Per la esterasi da fegato di maiale abbiamo seguito lo stesso protocollo della
esterasi da fegato di coniglio appena descritto ottenendo i seguenti risultati:
ESTERASI DA FEGATO DI MAIALE
(13,5 mg/ml)
Nam deamidase
104 U/ml
8 mU/mg
Carbossilesterase 1659 U/ml 122,9 U/mg
Anche in questo caso, entrambe le attività sono effettivamente presenti nella
preparazione commerciale ma con rapporto carbossilesterasi/deamidasi molto più
alto pari a ~15000.
PORCINE LIVER
(16.5ug/ul)
STD
97,4 KDa
66,2 KDa
60,0 KDa
45,0 KDa
31 KDa
14,4 KDa
Figura 18: SDS PAGE 12% della preparazione commerciale della PLE
Anche la preparazione enzimatica da fegato di maiale non è omogenea in
elettroforesi. Cerchiata, la banda attesa di ~60000 Da come per la precedente.
Anche per questa preparazione commerciale, abbiamo effettuato una TSK-DEAE
ed anche in questo caso, abbiamo osservato come per la esterasi di coniglio,
anche in quella di maiale entrame le attività coeluiscano in un unico picco di
assorbanza.
43
30
280nm
260nm
25
ml
20
15
10
5
0
-5
0
20
40
60
80
mAu
Figura 19: Profilo di assorbanza della cromatografia TSK-DEAE
3.2 Determinazione della Km su RLE e PLE
Entrambe le preparazioni enzimatiche commerciali di RLE e PLE, sono state
utilizzate per la determinazione del parametro cinetico Km, nei confronti del p-NPA
e Nam. Il valore di tale parametro può essere utile per comprendere meglio se tale
attività enzimatica ha un significato fisiologico.
Per l’analisi cinetica della reazione catalizzata dalla RLE, l’attività è stata
determinata con il saggio spettrofotometrico continuo variando la concentrazione
del substrato p-NPA. Sono state utilizzate le seguenti concentrazioni: 13,2 μM,
20,4 μM, 30 μM, 30,72 μM, 45,6 μM, 60,48 μM, 80,4 μM, 80,64 μM, 100,8 μM, 120
μM, 149,76 μM.
In figura è riportato il grafico della velocità della reazione in funzione della
concentrazione del substrato e il grafico dei doppi reciproci.
44
A
B
Grafico 1: A. Grafico lineare diretto di Eisental e Cornish-Bowden. B. Grafico di Lineweaver-Burk per la
RLE
Il grafico 1 mette in evidenza come l’enzima carbossilesterasi segua la cinetica di
Michaelis-Menten, in quanto la curva rappresentata un ramo di iperbole. Tale
comportamento è confermato dal grafico dei doppi reciproci, in cui i dati
sperimentali danno origine ad una retta intersecante l’asse delle ascisse (grafico
1). Dai dati grafici è stata calcolata una Km di 28,67 μM. I risultati da noi ottenuti
sono in accordo con i valori riportati in letteratura [33] [30].
Per la determinazione della Km per le PLE, come per la RLE, è stato utilizzato il
saggio spettrofotometrico in continuo e le concentrazioni di substrato utilizzate per
il saggio, vanno da una concentrazione di 38.6 μM fino a ~1800 μM. In figura è
45
riportato il grafico della velocità della reazione in funzione della concentrazione del
substrato e il grafico dei doppi reciproci.
A
B
Grafico 2: A Grafico lineare diretto di Eisental e Cornish-Bowden. B. Grafico di Lineweaver-Burk per la
PLE
Anche in questo caso, il grafico 2 mette in evidenza come l’enzima
carbossilesterasi segua la cinetica di Michaelis-Menten, in quanto la curva
rappresentata un ramo di iperbole. Dai dati grafici è stata calcolata una K m di
259,72 μM. I risultati da noi ottenuti sono in accordo con i valori riportati in
letteratura [64] [65].
46
Per l’analisi cinetica della reazione catalizzata dalla NDasi di coniglio e di maiale,
l’attività è stata determinata con il saggio HPLC discontinuo, come descritto nella
sezione 2, variando la concentrazione del substrato Nam.
Per la determinazione della Km di coniglio le concentrazioni di substrato vanno da
2.352 mM fino a 142.36 mM.
Nam Deamidase from Rabbit Liver
0.05
0.04
v
0.03
0.02
0.01
0.00
0
25
50
75
100
125
150
[Nam]
A
Lineweaver-Burk Nam Deamidase from Rabbit Liver
500
1/v
400
300
200
100
0
0.0
B
0.1
0.2
0.3
0.4
0.5
1/[Nam]
Grafico 3: A Grafico lineare diretto di Eisental e Cornish-Bowden. B. Grafico di Lineweaver-Burk per la
RLE
Il grafico 3 mette in evidenza come l’enzima segua la cinetica di Michaelis-Menten,
in quanto la curva rappresentata un ramo di iperbole. Dai dati grafici è stata
calcolata una Km di 51,91 mM. Da questa analisi emerge che mentre per la
47
Carbossilesterasi (from Rabbit Liver), che ha una Km di ~30 μM, lavoriamo a Vmax
nei saggi di reazione, per la Nam Deamidase, non lavoriamo a Vmax, ma lavoriamo
intorno alla Km.
Per la determinazione della Km di maiale, le concentrazioni di substrato vanno da
4,47 mM fino a 272,86 mM.
Nam Deamidase from PORCINE LIVER
0.07
0.06
0.05
v
0.04
0.03
0.02
0.01
0.00
0
100
200
300
[Nam]
A
Lineweaver-Burk Nam Deamidase from Porcine Liver
600
500
1/v
400
300
200
100
0
0.0
B
0.1
0.2
0.3
1/[Nam]
Grafico 4: Grafico lineare diretto di Eisental e Cornish-Bowden. B. Grafico di Lineweaver-Burk
Anche in questo caso l’enzima segue la cinetica di Michaelis-Menten, come
mostrato dai grafici. Dalle prove è stato calcolato un valore di Km di 225,7 mM.
Da questi valori di Km emerge essendo questi valori così alti, probabilmente
potrebbero non avere un significativo ruolo fisiologico all’interno della cellula.
48
3.3 Mutagenesi pcDNA 3.1/hCE-1b
Dopo aver dimostrato che entrambe le attività enzimatiche della CE e NDasi sono
a carico dallo stesso enzima, si è deciso di passare allo studio dell’enzima umano.
Il glicerinato contenente la sequenza della carbossilesterasi umana isoforma b è
stato rimesso in crescita e successivamente mandato a sequenziare. Dall’analisi
di sequenza effettuata sul plasmide in nostro possesso, ci siamo accorti della
presenza di una mutazione sul nucleotide 1522: è sostituita la citosina in posizione
1522, che invece è presente in tutte le numerose sequenze in banca dati.
Figura 20: Sequenza nucleotidica della hCES1b allineata con la sequenza nucleotidica della rCES
Confrontando la sequenza della carbossilesterasi umana con quella di coniglio,
della quale si conosce anche la struttura cristallografica [60], ci si è accorti che:
1.
Questa citosina sostituita determina lo scambio di un aminoacido con uno
completamente diverso.
2.
Inoltre, questo aminoacido sostituito è in una posizione vicina al sito attivo e
potrebbe essere importante per l’attività.
49
Dopo aver effettuato una mutagenesi per introdurre il nucleotide corretto, ed
esserci accertati che la mutazione ha avuto esito positivo, decidiamo di procedere
con la trasfezione transiente del costrutto ricombinante pcDNA/hCE-1b in cellule di
COS-7.
Al termine della trasfezione transiente, il lisato cellulare delle cellule trasfettate, è
stato caricato su una colonnina Ni-NTA, che lega saldamente le proteine con code
istidiniche permettendo la purificazione della proteina ricombinante. L’avvenuta
espressione della proteina nelle cellule è stata determinata mediante rivelazione in
immunitaria della membrana PVDF sulla quale sono state immobilizzate le
proteine di cellule trasfettate e di cellule di controllo, cioè non trasfettate, e mostra
la presenza di un’unica banda, del peso molecolare atteso, solo in corrispondenza
delle cellule trasfettate.
CP
CT
STD
Figura 21: Immunoblotting di lisato di cellule COS7 non trasfettate (CT) e di lisato di cellule COS7
trasfettate con hCES1b (CP)
L’espressione della proteina è stata confermata anche mediante rivelazione
dell’attività CE che è risultata essere:
COS7 trasfettate
COS7 non trasfettate
CE
0,9415 U/ml 0,2482 U/mg
NDasi
ND
ND
CE
0,010 U/ml
-
NDasi
ND
ND
Nello stesso campione non è stata rilevata alcuna attività NDasica a causa della
scarsità del materiale di partenza. I saggi di attività sono stati allestiti come
descritti nella sezione 2.2, 2.3.
50
Il lisato di cellule trasfettate, purificato attraverso la Ni-NTA, è stato
successivamente dializzato con la colonna PD-10 con lo scopo di allontanare
l’imidazolo utilizzato per staccare la proteina ricombinante dalla resina di affinità
Ni-NTA, in quanto si è visto che l’imidazolo determina l’autoidrolisi del p-NPA [61],
impedendoci in questo modo di determinare correttamente l’attività della CE.
Gli stessi campioni sono stati caricati su gel per vedere l’eventuale presenza di
proteina ricombinante:
CP
STD
1,6 μg di CAMPIONE CARICATO
97,4 KDa
60,0 KDa
66,2 KDa
45,0 KDa
31 KDa
14,4 KDa
Figura 22: SDS-PAGE al 12% di lisato di cellule COS7 trasfettate
Come ulteriore prova che la proteina purificata era effettivamente la hCE1b,
abbiamo effettuato un esperimento di proteolisi limitata e sequenziati alcuni
frammenti ottenuti.
Figura 23: SDS-PAGE 12% della hCE1b sottoposta a proteolisi limitata
51
La sequenza dei frammenti cerchiati ha dato il seguente risultato: HPSSPPVVD
Abbiamo cercato questa sequenza all’interno di quella della Carboxylesterase 1
isoform b precursor trovando un riscontro positivo:
Carboxylesterase 1 isoform b precursor [Homo sapiens]
>gi|68508965|ref|NP_001020365.1| carboxylesterase 1 isoform b precursor [Homo
sapiens]
MWLRAFILATLSASAAWGHPSSPPVVDTVHGKVLGKFVSLEGFAQPVAIFLGIPF
AKPPLGPLRFTPPQPAEPWSFVKNATSYPPMCTQDPKAGQLLSELFTNRKENIP
LKLSEDCLYLNIYTPADLTKKNRLPVMVWIHGGGLMVGAASTYDGLALAAHENVV
VVTIQYRLGIWGFFSTGDEHSRGNWGHLDQVAALRWVQDNIASFGGNPGSVTIF
GESAGGESVSVLVLSPLAKNLFHRAISESGVALTSVLVKKGDVKPLAEQIAITAGC
KTTTSAVMVHCLRQKTEEELLETTLKMKFLSLDLQGDPRESQPLLGTVIDGMLLL
KTPEELQAERNFHTVPYMVGINKQEFGWLIPMQLMSYPLSEGQLDQKTAMSLLW
KSYPLVCIAKELIPEATEKYLGGTDDTVKKKDLFLDLIADVMFGVPSVIVARNHRD
AGAPTYMYEFQYRPSFSSDMKPKTVIGDHGDELFSVFGAPFLKEGASEEEIRLSK
MVMKFWANFARNGNPNGEGLPHWPEYNQKEGYLQIGANTQAAQKLKDKEVAF
WTNLFAKKAVEKPPQTEHIEL
La sequenza: MWLRAFILATLSASAAWG costituisce il peptide segnale che
probabilmente la cellula produce e taglia dopo che il peptide è stato sintetizzato.
Il peptide segnale è una breve sequenza amminoacidica, costituita da circa 20-40
residui, in massima parte idrofobici, che permette il corretto smistamento delle
proteine sintetizzate dalla cellula. In genere è localizzato all'estremità N-terminale
della proteina, anche se, in alcune proteine, è stato riscontrato a livello del Cterminale.
Rappresenta, dunque, la prima porzione della proteina ad essere sintetizzata (la
sintesi proteica procede infatti in direzione N-ter/C-ter) e quindi a fuoriuscire dal
ribosoma.
Esso viene riconosciuto da una particolare nucleo-proteina, la particella di
riconoscimento del segnale (RSP) la quale presenta, inoltre, domini di
riconoscimento per specifiche proteine di superficie del RE. In tal modo, non
appena il peptide segnale fuoriesce dal ribosoma, viene legato dalla RSP che,
52
interagendo con le proteine del RE, trascina con sè, sulla superficie del RE, il
complesso proteina-ribosoma, dando in tal modo origine al RE ruvido.
Una volta sulla membrana del RE, la proteina nascente viene fatta entrare nel
lume del reticolo, grazie a canali proteici ottenuti dall'assemblaggio di più sub-unità
proprio a seguito dell'arrivo del complesso proteina-ribosoma. La superficie interna
di questi canali è costituita in massima parte da residui amminoacidici idrofobici
che possono interagire con i residui idrofobici del peptide segnale. In tal modo la
proteina penetra, man mano che viene sintetizzata, nel lume del RE, mentre la
sua estremità N-terminale rimane adesa al canale. Terminata la sintesi della
proteina, specifiche peptidasi, localizzate sul versante luminale della membrana
del RE, scindono il peptide segnale permettendo il rilascio completo della proteina
nel lume del RE, dove subirà diverse trasformazioni post-traduzionali (es. la
glicosilazione).
Purtroppo a causa della scarsità di materiale a disposizione, nonostante numerosi
tentativi per aumentare la quantià di materiale prodotto dalle cellule, non si è
riusciti a determinare l’attività NDasica della proteina ricombinante in cellule di
COS7.
53
4 RISULTATI (parte II)
4.1 Studio sulle vie biosintetiche del NAD
Nella seconda parte del lavoro di tesi abbiamo iniziato la messa a punto di un
saggio per la determinazione delle attività degli enzimi chiave delle vie
biosintetiche amidate e deamidate del NAD. Il principio del saggio è schematizzato
in figura:
I prodotti delle reazioni catalizzate da questi enzimi sono NMN e NaMN. L’NaMN è
convertito in NaAD usando l’ NaMN adeniltrasferasi ricombinante (NadD), il quale
mostra una stretta specificità per il mononucleotide deamidato e catalizza la sua
adenilazione in modo ATP-dipendente. L’ NMN è convertito a NaAD in due
reazioni consecutive: la prima prevede la sua deamidazione a NaMN ad opera
dell’ NMN deamidasi ricombinante (PncC), la seconda consiste nell’adenilazione
dell’ NaMN a NaAD catalizzata da NadD. La completa conversione dei
mononucleotidi a NaAD è assicurata dall’irreversibilità della reazione catalizzata
da PncC, dall’utilizzo di un eccesso di ATP che sposta l’equilibrio della reazione
NadD-dipendente verso la formazione dell’ NaAD e dalla stretta specificità di
substrato dell’enzima
NadD. Una volta formato, il NaAD viene quantificato
mediante analisi HPLC o convertito a NAD mediante la NAD sintetasi
ricombinante da E.coli (NadE). Il NAD così formato viene rilevato attraverso il
saggio ciclico fluorimetrico sviluppato da Graeff e Lee [63] che prevede due
reazioni enzimatiche: nella prima reazione l’alcol deidrogenasi da S.cerevisiae
54
riduce il NAD a NADH, nella seconda reazione la diaforasi da C.kluyveri riossida il
NADH a NAD con la simultanea conversione della resazurina nel composto
altamente fluorescente resorufina.
4.2 Conversione dell’NaMN e NMN in NaAD e quantificazione
mediante HPLC
L’analisi diretta dei mononucleotidi mediante HPLC è ostacolata dalla loro scarsa
capacità di legarsi alla fase stazionaria apolare delle colonne cromatografiche che
operano in fase inversa e quindi dalla loro tendenza ad eluire insieme ad altre
molecole nel volume escluso. Al contrario, l’ NaAD si lega fortemente alla resina
ed eluisce con la fase mobile polare, consentendo di utilizzare un gradiente e
quindi di trovare le condizioni migliori per una risoluzione ottimale. In aggiunta
l’NaAD ha un coefficiente di estinzione molare 3,8 volte maggiore di quello dei
mononucleotidi.
Il metodo è stato inizialmente sviluppato per valutare la concentrazione
intracellulare dei mononucleotidi NaMN e NMN. I livelli dei mononucleotidi sono
stati analizzati come descritto in Materiali e Metodi. L’intera procedura è
schematizzata in Figura 24.
55
Figura 24: Schema della procedura utilizzata per la determinazione dell’NMN e dell’NaMN descritta
nella sezione Materiali e Metodi. In basso è rappresentata parte del cromatogramma dell'analisi in
HPLC dove viene mostrato il picco dell’ NaAD formato nelle tre miscele di reazione.
Al fine di verificare la stechiometrica conversione dei mononucleotidi in NaAD,
quantità note di NMN e NaMN, in un range da 0,2 a 1,0 nmoli, sono state
convertite a NaAD e il dinucleotide formato è stato quantizzato in HPLC.
56
Come mostrato in figura i mononucleotidi sono stati completamente convertiti in
NaAD in 5 minuti, usando 4 mU di PncC e NadD.
È stata inizialmente valutata l’accuratezza del metodo per l’estrazione dei
mononucleotidi utilizzando cellule Hep G2 e C6. A tale scopo quantità note di
NaMN ed NMN sono state aggiunte agli estratti nucleotidici delle cellule preparati
mediante estrazione acida o etanolo bollente, come descritto nella sezione 2. Il
recupero è stato determinato calcolando la differenza tra i campioni con lo spike e
quelli senza spike. Entrambi i metodi estrattivi danno risultati simili, con un
recupero in un intervallo compreso tra il 90% e 96%. L’estrazione acida è più
rapida rispetto all’estrazione con etanolo; quest’ultima però rappresenta anche
uno step di concentrazione e può essere conveniente usarla quando si trattano
campioni diluiti. Il limite di rilevazione del saggio in HPLC è 3 pmoli di NaAD
iniettate, che corrispondono a 4,5 pmoli di NMN o NaMN nella miscela di reazione.
La sensibilità del metodo è 0,4 pmol mononucleotidi/mg di pellet cellulare.
L’analisi è stata applicata ad estratti nucleotidici acidi di Hep G2 e C6 e i risultati
sono mostrati in tabella.
57
C6 Rattus Hep-G2
pmoli/mg
NaAD
0,4
≤ 0,4
NaMN
1,44
0,52
NMN
1,12
8,2
Si osserva che negli astrociti i livelli di NaMN e NMN sono molto simili. Nelle
cellule epatiche invece i livelli di NMN sono significativamente più alti di quelli dell’
NaMN. Questi risultati suggeriscono che negli epatociti la biosintesi del NAD
avvenga preferenzialmente attraverso la via amidata Nam -> NMN -> NAD,
mentre negli astrociti anche la via deamidata Na -> NaMN -> NaAD -> NAD
potrebbe contribuire in maniera significativa alla sintesi del NAD.
4.3 Conversione dell’NaAD a NAD e quantificazione mediante
saggio ciclico fluorimetrico
Per aumentare la sensibilità ed esecuzione del saggio al fine di usarlo per la
determinazione dell’attività enzimatica degli enzimi responsabili della biosintesi
dell’ NMN e NaMN, il NaAD che si forma nella miscela di reazione viene misurato
dopo la sua conversione a NAD mediante il saggio ciclico fluorimetrico, come
descritto in materiali e metodi.
La linearità del saggio è stata valutata usando differenti concentrazioni di NMN,
NaAD e NAD in un intervallo compreso tra 0,5 e 3 pmol. Come mostrato in figura
sia la fluorescenza che la linearità del saggio sono simili per i tre nucleotidi: ciò
indica che la conversione dell’NMN in NaAD e NAD è stechiometrica e la
presenza degli enzimi ancillari non influisce sulla fluorescenza.
58
59
5 CONCLUSIONI (parte I)
La Nam, conosciuta anche come niacinamide è l’amide dell’acido nicotinico. La
Nam è una vitamina idrosolubile che fa parte del gruppo delle vitamine B. La Na,
conosciuto anche come niacina, è convertito a Nam, in vivo, e, se le due sono
identiche per la funzione vitaminica, la Nam non ha gli stessi effetti farmacologici
della niacina. Nelle cellule la niacina è annessa nel NAD e NADP. NAD+ e NADP+
sono coenzimi in un’ampia varietà di reazioni enzimatiche di ossidoriduzioni.
L’importanza di tale vitamina nella dieta è stata messa in evidenza 70 anni fa dalla
scoperta che la sua carenza nel regime alimentare era responsabile dello sviluppo
della pellagra umana. Il NAD+, infatti, può essere descritto come un anello
nicotinamidico, legato attraverso un legame -N-glicosidico al C-1 del ribosio di un
residuo adenosindifosforibosilico. Tale legame viene scisso, in reazioni non-redox
NAD dipendenti, da enzimi implicati in importanti processi cellulari quali il CD38, le
PARPs e le sirtuine, con la conseguente liberazione di Nam e metaboliti vari, quali
rispettivamente
cADPR,
ADP-riboso,
e
O-acetil-ADP-riboso,
implicati
nei
meccanismi di trasduzione del segnale.
Questi enzimi, utilizzando il NAD come substrato e quindi producendo un
consumo effettivo della molecola, contribuiscono alla modulazione della maggior
parte del pool cellulare del dinucleotide piridinico; tale attività degradativa è a sua
volta regolata dalla Nam, il prodotto comune di tutte le reazioni sopra citate. E’
stato dimostrato, infatti, che la Nam è un inibitore potente degli enzimi che
degradano il NAD. Poiché le costanti d’inibizione (Ki) della Nam per tali enzimi è
dell’ordine micromolare, le variazioni anche minime della concentrazione di tale
molecola possono influire su processi cellulari significativi.
Inoltre, la Nam svolge importanti funzioni anche dal punto di vista farmacologico.
La somministrazione di dosi anche 100 volte più elevate rispetto al fabbisogno
giornaliero come vitamina (15-20 mg/giorno) è utilizzata, o si trova in fase di studio
preclinico, per la cura di diverse patologie quali tubercolosi, AIDS, diabete,
malattie neurodegenerative. Sebbene i meccanismi molecolari dell’azione della
Nam in queste patologie devono ancora essere delucidati in dettaglio, l’ effetto
60
benefico di questo nucleotide piridinico, in alcune di queste patologie, sembra
essere consistente con un meccanismo nel quale la Nam inibisce gli enzimi
implicati nel catabolismo del NAD.
Uno degli enzimi oggetto del presente lavoro è la nicotinamide deamidasi. Il gene
per tale enzima è stato identificato nei procarioti e negli eucarioti mono e
pluricellulari (S. Cerevisiae e C. Elegans). Mentre i dati bioinformatici supportano
la tesi che tale gene non esiste nell’uomo in quanto non è stato trovato un
omologo umano, alcuni lavori in letteratura riportano la presenza dell’attività
nicotinamide deamidasica in alcuni tessuti di mammiferi quali il ratto, il coniglio e
l’uomo.
In un precedente lavoro è stato purificato ad omogeneità l’enzima dotato di attività
NDasica da fegato di coniglio; la sequenza di tale enzima ha rilevato che si
trattava in realtà della CES. Le carbossilesterasi sono enzimi che svolgono un
ruolo importante nel metabolismo e nella detossificazione di numerosi composti
endogeni e xenobiotici. Tali enzimi, idrolizzano il legame di un’ampia varietà di
esteri carbossilici, tra cui numerosi composti analgesici e narcotici quali aspirina,
cocaina, eroina ecc. E’ stato rilevato inoltre che possono svolgere anche un’attività
deamidasica nei confronti di ammidi aromatiche.
In questo progetto di ricerca, abbiamo confermato che la CES da mammifero è in
grado di deamidare la Nam ad Na, usando preparazioni commerciali di esterasi di
fegato di coniglio e di maiale.
Attraverso dei saggi enzimatici si è osservato come tali enzimi siano in grado di
catalizzare sia la reazione di rottura dell’estere carbossilico nel corrispondente
alcol ed acido carbossilico, sia la reazione di deamidazione della Nam ad Na. Per
essere certi che è lo stesso enzima a catalizzare entrambe le reazioni, attraverso
delle prove cromatografiche, abbiamo osservato la coeluizione di ambedue le
attività (carbossilesterasica e deamidasica) in un unico picco di assorbanza.
Lo studio della cinetica di queste due preparazioni enzimatiche nei confronti della
Nam, ha mostrato dei valori di Km molto alti, il che ci farebbe escludere un
possibile ruolo fisiologico dell’enzima.
L’obiettivo successivo è stato quello di valutare se anche la CES umanana fosse
dotata di attività nam deamidasica.
61
Nell’uomo la proteina omologa alla CE di coniglio, hCE1 (identità di sequenza con
rCE = 81%) è codificata dal gene CES-1 che può dare origine a 3 isoforme (a,b,c).
Per dimostrare che tale enzima è responsabile dell’attività deamidasica è stata
tentata l’espressione della proteina in sistemi di espressione eucariotici quali le
cellule di mammifero COS7 allo scopo di produrre la proteina con le necessarie
modifiche post-traduzionali, quali la glicosilazione, indispensabile per una corretta
funzionalità enzimatica. Tuttavia, non è stata ottenuta una quantità sufficiente di
proteina idonea agli studi necessari.
Va sottolineato che per rilevare l’attività deamidasica della CE umana è stata
utilizzata un’alta concentrazione di Nam(100 mM) in accordo con il valore della Km
per la Nam riportato in letteratura pari a 40 mM. L’alto valore di K m indica
un’attività deamidasica non significativa da un punto di vista fisiologico, a sostegno
dell’ ipotesi che nei mammiferi, la via di deamidazione della Nam a Na è assente.
L’unica eccezione può essere rappresentata da una certa attività deamidasica
svolta nell’intestino dai batteri commensali che potrebbe contribuire al salvage
della Nam dell’ospite. Tuttavia, non si può escludere che l’attività enzimatica sia
sottoposta a regolazione. Nel tentativo di individuare un eventuale effettore di tale
attività, potrebbe essere interessante continuare ad indagare sull’ipotesi che la
carbossilesterasi umana possa avere anche attività Nam deamidasica e svolgere
quindi un ruolo importante anche nel metabolismo di questa vitamina.
I risultati ottenuti nella prima parte della tesi, suggeriscono l’assenza di una
significativa attività Nam deamidasica in cellule di mammifero. Nei mammiferi è
anche assente l’omologo dell’enzima NMN deamidasi che è stato recentemente
identificato nei batteri. Sembra quindi che non esista nessuna connessione tra la
via amidata verso quella deamidata per la sintesi del NaAD.
62
6 CONCLUSIONI (parte II)
Le due forme mononucleotidiche della vitamina B 3, il nicotinato mononucleotide
(NaMN) e la sua forma amidata, nicotinamide mononucleotide (NMN), vengono
sintetizzate nella cellula a partire dalle tre forme della vitamina B3: acido nicotinico
(Na),
nicotinammide
(Nam)
e
nicotinammide
riboside
(NamR).
I
due
mononucleotidi sono intermedi della biosintesi del coenzima NAD; mentre l’NMN
viene direttamente convertito nel coenzima NAD; l’NaMN viene dapprima
adenilato nel corrispondente dinucleotide NaAD che successivamente viene
amidato a NAD.
Negli ultimi anni, l’NMN è stato oggetto di particolari studi in quanto si è osservato
che la sua somministrazione ripristina significativamente i livelli di NAD nel fegato
e nel tessuto adiposo di topi diabetici dove si sono ridotti, migliorando la tolleranza
al glucosio [66] e ripristina la secrezione d’insulina in topi alimentati con una dieta
ad alto contenuto di fruttosio [67]. Inoltre, è stato dimostrato che stimola la
proliferazione delle cellule della muscolatura lisca vascolare, contribuendo al
rimodellamento della parete del vaso sanguigno in seguito a danneggiamento [68].
In un altro lavoro è riportato che l’NMN svolge un ruolo protettivo anche nel
sistema nervoso; è stato infatti dimostrato che è in grado di ridurre i danni
cerebrali indotti da ischemia [69].
L’attenzione nei confronti di questo mononucleotide, porta con sè la necessità di
avere a disposizione un saggio semplice e sensibile per la determinazione dei suoi
livelli in campioni di varia natura.
I metodi riportati in letteratura per la rivelazione dei mononucleotidi sono basati
sulla separazione in HPLC e rivelazione in UV [70], separazione in HPLC seguita
da analisi in spettrometria di massa (LC/MS/MS ANALYSIS) [71] e cromatografia
in HPLC dopo derivatizzazione dell’NMN in un composto fluorescente [72].
In alcuni casi, questi metodi hanno dato risultati contrastanti, ad esempio, per la
determinazione dell’NMN nel siero, il quale è risultato essere 80-90 μM quando
determinato mediante cromatografia HPLC e rivelazione in UV, e inferiore a 50 nM
con la LC/MS/MS ANALYSIS.
63
L’obiettivo di questo lavoro, è stato quello di mettere a punto un saggio sensibile,
ma anche accurato, rapido e possibilmente meno costoso di quelli sopra citati, che
potesse essere applicato anche alla determinazione degli enzimi della via
biosintetica del NAD che catalizzano la formazione dell’NMN e dell’ NaMN.
Il nostro saggio prevede la trasformazione dei mononucleotidi a NaAD e la
successiva quantificazione del dinucleotide mediante HPLC.
Gli enzimi utilizzati per le reazioni sono coinvolti nella biosintesi del NAD nei
batteri e tutti vengono utilizzati in forma ricombinante ed hanno delle peculiari
proprietà catalitiche.
PncC è stata identificata e caratterizzata per la prima volta nel nostro laboratorio.
NadD è specifico per l’NaMN.
Per aumentare la sensibilità e velocità di esecuzione del saggio, il NaAD viene
trasformato a NAD e quest’ultimo viene rivelato mediante un saggio ciclico
fluorimetrico già descritto in letteratura [63].
In conclusione: utilizzando gli enzimi della biosintesi del NAD nei batteri, abbiamo
messo a punto un saggio per la contemporanea rivelazione dei mononucleotidi
piridinici.
Nella seconda parte del mio lavoro di tesi, è stato iniziato lo sviluppo di un saggio
per la determinazione delle attività degli enzimi chiave delle diverse vie (amidata e
deamidata). Si tratta di un saggio che fa uso di enzimi ancillari di origine batterica
che convertono i prodotti delle reazioni catalizzate dagli enzimi di interesse (NMN
e NaMN) in NaAD. Quest’ultimo viene poi valutato mediante un saggio ciclico
fluorimetrico. La messa a punto del saggio ha previsto inizialmente l’ottimizzazione
della procedura per la quantizzazione dei mononucleotidi NaMN e NMN.
64
Bibliografia
[1]
F. Li, Z. Z. Chong and K. Maiese, “Cell life versus cell longevity: the mysteries surrounding
the NAD+ precursor nicotinamide,” Curr. Med. Chem., no. 13, pp. 883-895, 2006.
[2]
H. D. Turner JB, «THE ABSORPTION OF SOME B-GROUP VITAMINS BY SURVIVING
RAT INTESTINE PREPARATIONS,» Q.J. EXP. PHYSIOL. COGN. MED. SCI., vol. 47, pp.
107-33, 1962.
[3]
C. Bernofsky, "Physiology aspects of pyridine nucleotide regulation in mammals," Mol. Cell
Biochem., no. 33, pp. 135-143, 1980.
[4]
I. A. H. O. IJICHI H, «STUDIES ON THE BIOSYNTHESIS OF NICOTINAMIDE
ADENINE DINUCLEOTIDE. COMPARATIVE IN VIVO STUDIES ON NICOTINIC
ACID, NICOTINAMIDE, AND QUINOLINC ACID AS PRECURSOR OF
NICOTINAMIDE ADENIN DINUCLEOTIDE,» J. BIOL. CHEM., vol. 241, pp. 3071-7,
1966.
[5]
Y. Tanigawa, M. Shimoyama, R. Murashima, T. Ito and K. Yamaguchi, “The role of
microrganisms as a function of nicotinamide deamidation in rat stomach,” Biochem. Biophys.
Acta, no. 201, pp. 394-397, 1970.
[6]
C. Streffer and J. Benes, “Nicotinamide mononucleotide. Determination of its enzymatic
formation in vitro and its physiological role for the biosynthesis of nicotinamide-adenine
dinucleotide in mice,” Eur. J. Biochem, no. 21, pp. 357-362, 1971.
[7]
C. A. Benavente, M. K. Jacobson and E. L. Jacobson, "NAD in skin: therapeutic approaches
for niacin," Current Pharmaceutical Design, vol. 15, no. 1, pp. 29-38, 2009.
[8]
R. A. Jacob and M. E. Swendseid, “Niacin,” Present Knowledge in Nutrition, pp. 184-190,
1996.
[9]
K. Karthikeyan and D. M. Thappa, “Pellagra and skin,” International Journal of
Dermatology, vol. 41, no. 8, pp. 476-481, 2002.
[10] "Nutrient reference values for Australian and New Zealand, including recommended dietary
intake," in National Health and Medical Research Council publication, Canberra, Australia,
2006, pp. 1-329.
[11] S. Kitamura, K. Nitta, Y. Tayama, C. Tanoue, K. Sughara, T. Inoue, T. Horie and S. Ohta,
“Aldehyde oxidase-catalyzed metabolism of N1-methylnicotinamide in vivo and in vitro in
chimeric mice with humanized liver,” Drug Metab Dispos, no. 36, pp. 1202-1205, 2008.
[12] J. Goldberger, "The etiology of Pellagra," Public Health Rep., no. 29, pp. 1683-1686, 1914.
65
[13] C. A. Elvehjem, R. J. Madden, F. M. Strong and D. W. Wolley, "Relation of nicotinic acid
and nicotinic acid amide to canine black tongue," J. Am. Chem. Soc., no. 59, p. 1767, 1937.
[14] A. E. Axerold, R. J. Madden and C. A. Elvehjem, "The effort of nicotinic acid deficiency
upon the coenzyme I content of animal tissue," J. Biol. Chem., no. 131, pp. 85-93, 1939.
[15] G. G. Graham, "Starvation in the modern world," N Eng J Med, no. 328, pp. 1058-1061,
1993.
[16] W. T. Penberthy, “Pharmacological targeting of IDO-mediated tolerance for treating
autoimmune disease,” Curr Drug Metab., vol. 3, no. 8, pp. 245-266, 2007.
[17] W. M. Hendricks, "Pellagra and pellagralike dermatoses: etiology, differential diagnosis,
dermatopathology, and treatment," Seminars in Dermatology, vol. 10, no. 4, pp. 282-292,
1991.
[18] P. Chambon, J. D. Weill and P. Mandel, "Nicotinamide mononucleotide activation of a new
DNA-dependent polyadenylic acid synthesizing nuclear enzyme," Biochemical and
Biophysical Research Communications, vol. 11, no. 1, pp. 39-43, 1963.
[19] F. J. Oliver, J. Menissier-de Murcia and G. De Murcia, “Ploy(ADP-Ribose) polymerase in the
cellular response to DNA damage, apoptosis, and disease,” American Journal of Human
Genetics, vol. 64, no. 5, pp. 1282-1288, 1999.
[20] D. Surjana, G. M. Halliday and D. L. Damian, “Role of nicotinamide in DNA damage,
mutagenesis and DNA repair,” J. Nucleic Acids, p. doi: 10.4061/2010/157591, 2010.
[21] A. Ieraci and D. G. Herrera, "Nicotinamide protects against ethanol-induced apoptotic
neurodegeneration in the developing mouse brain," PLoS Med, p. 3:e101, 2006.
[22] D. M. Capuzzi, J. M. Morgan, O. A. Brusco and C. M. Intenzo, "Niacin dosing: relationship
to benefits and adverse effects," Curr. Atheroscl. Rep., no. 2, pp. 64-71, 2000.
[23] V. S. Kamanna and M. L. Kashyap, “Mechanism of action of niacin on lipoprotein
metabolism,” Curr. Atheroscl. Rep., no. 2, pp. 36-46, 2000.
[24] G. Magni, A. Amici, M. Emanuelli, G. Orsomando, N. Raffaelli and S. Ruggieri,
“Enzymology of NAD+ homeostasis in man,” Cell. Mol. Life Sci., no. 60, pp. 1-16, 2003.
[25] G. Magni, “Enzymologi of NAD+ synthesis,” Adv Enzymol Relat Areas Mol Biol, no. 73, pp.
135-182, 1999.
[26] T. Penfound and J. W. Foster, “Biosynthesis and recycling of NAD in Escherichia coli and
Salmonella Typhimurium,” Cellular and Molecular Biology, vol. 1, pp. 721-730, 1996.
66
[27] O. Kursanov, “NAD biosynthesis: identification of the tryptophan to quinolinate pathway in
bacteria,” Chem biol, vol. 10, no. 12, pp. 1195-1204, 2003.
[28] J. W. Foster, "Regulation of NAD metabolism in Salmonella typhimurium: molecular
sequence analysis of the bifunctional nadR regulator and the nadA-pnuC operon," J Bacteriol,
no. 172, pp. 4187-4196, 1990.
[29] J. W. Foster and A. G. Moat, "Nicotinamide adenine dinucleotide biosynthesis and pyridine
nucleotide cycle metabolism in microbial systems," Microbiol Rev, vol. 44, pp. 83-105, 1980.
[30] S. Su, L. Albizati and S. Chaykin, “Nicotinamide deamidase from rabbit liver. II. Purification
and properties,” J. Biol. Chem., no. 244, pp. 2956-2965, 1969.
[31] S. S. Gillam, J. G. Watson and S. Chaykin, "Nicotinamide deamidase from rabbit liver 3.
Inhibition and sedimentation studies," Arch. Biochem. Biophys., no. 157, pp. 268-284, 1973.
[32] J. Kirchner, J. G. Watson and S. Chaykin, “Nicotinamide deamidase from rabbit liver,” J.
Biol. Chem., no. 241, pp. 953-960, 1966.
[33] L. D. Albizati and J. L. Hedrick, “Active-site studies on rabbit liver nicotinamide deamidase,”
Biochemistry, vol. 11, no. 8, pp. 1508-1517, 1972.
[34] M. Wintzerith, A. Dierich and P. Mandel, “Nicotinamide deamidase released by
neuroblastoma cell cultures,” J. Neurochem., no. 33, pp. 677-685, 1979.
[35] A. B. Pardee, E. J. J. Benz, D. A. St Peter, J. N. Krieger, M. Meuth and H. W. Trieshmann,
“Hyperproduction and purification of nicotinamide deamidase, a microconstitutive enzyme of
Escherichia coli,” J. Biol. Chem., no. 246, pp. 6792-6796, 1971.
[36] E. S. Dickinson and T. K. Sundaram, “Chromosomal location of a gene defining nicotinamide
deamidase in Escherichia Coli,” J. Bacteriol, no. 101, pp. 1090-1091, 1970.
[37] C. Yan and D. L. Sloan, “Purification and characterization of nicotinamide deamidase from
yeast,” J. Biol. Chem., no. 262, pp. 9082-9087, 1987.
[38] J. CasHman, “Pharmacokinetics and molecular detoxification,” Environ. Health Perspect, no.
104, pp. 23-40, 1996.
[39] T. Satoh and M. Hosokawa, “The mammalian carboxylesterases: from molecules to
function,” Annu. Rev. Pharmacol. Toxicol., no. 38, pp. 257-288, 1998.
[40] F. M. Williams, Clin. Pharmacokinet, no. 10, pp. 392-403, 1985.
[41] N. Bodor and P. Buchwald, P. Med. Res. Rev., no. 20, pp. 392-403, 2000.
67
[42] J. M. Joly and T. M. Brown, Toxicol. Appl. Pharmacol., no. 84, pp. 523- 532, 1986.
[43] M. R. Brzezinski, Drug Metab. Dispos., no. 25, pp. 1089-1096, 1997.
[44] L. M. Kamendulis, M. R. Brzezinski, E. V. Pindel, W. F. Bosron and R. A. Dean, J.
Pharmacol. Exp. Ther., no. 32, pp. 3735-3738, 1996.
[45] S. Bencharit, C. C. Edwards, C. L. Morton, E. L. Howard-Williams, P. Kuhn, P. M. Potter
and M. R. Redinbo, “Multisite Promiscuity in the Processing of Endogenous Substrates by
Human Carboxylesterase 1,” J Mol Biol., vol. 363(1), pp. 201-214, 2006.
[46] J. S. Munger, “A serine esterase released by humanalveolar macrophages is closely related to
liver microsomal carboxylesterases,” J. Biol. Chem., no. 266, pp. 18832-18838, 1991.
[47] H. Schwer, “Molecular cloning and characterization of a novel putative carboxylesterase,
present in human intestine and liver,” Biochem. Biophys. Res. Commun., no. 233, pp. 117120, 1997.
[48] M. Mori, “cDNA cloning, characterization and stable expression of novel human brain
carboxylesterase,” FEBS Lett., no. 458, pp. 17-22, 1999.
[49] M. P. Potter, J. S. Wolverton, C. L. Morton, M. Wierdl and M. K. Danks, “Cellular
localization domains of a rabbit and human carboxylesterase: influence on ironetecan (CPT11) metabolism by the rabbit enzyme,” Cancer Research, no. 58, pp. 3627-3632, 1998.
[50] U. K. Laemmli, “cleavage of structural proteins during the assembly of the head of
bacteriophage T4,” Nature, vol. 227, no. 5259, pp. 680-685, 1970.
[51] H. Neurath and W. K.A., Proc. Natl. Acad. Sci., no. 73:3825-3832, 1976.
[52] O. M., Ann. Rev. Biochem., no. 36:55-76, 1967.
[53] R. Beynon and J. Bond, Mol. Aspects Med., no. 9: 175-287, 1987.
[54] H. Neurath, “In protein folding,” Jaenicke R. ed., no. 501-504, 1980.
[55] H. Neurath, Chem. Scripta, no. 27B: 221-229, 1986.
[56] S. Hubbard, “The structural aspects of limited proteolysis of native proteins,” Biochimica et
Biophysica Acta, pp. 1-15.
[57] A. Fontana, P. Polverino de Laureto, E. Scaramella, M. Zambonin and V. De Filippis, Fold.
Des., no. 2:17-26, 1997.
[58] Y. Hu, C. Fenwick and A. English, Inorg. Chim. Acta, no. 242:261-69, 1996.
68
[59] J. Crowe, B. Masone and J. Ribbe, “One-step purification of recombinant proteins with the
6xHis tag and Ni-NTA resin,” Methods Mol. Biol., vol. 58, pp. 491-510, 1996.
[60] S. Bencharit, C. L. Morton, E. L. Howard-Williams, M. K. Danks, P. M. Potter and M. R.
Redinbo, “Structural insight into CPT-11 activation by mammalian carboxylesterases,”
Nature Structural Biology, vol. 9, no. 5, pp. 337-342, 2002.
[61] B. MM., “A rapid and sensitive method for the quantification of microgram quantities of
protein utilizing the principle of protein-dye binding.,” Anal. Biochem., vol. 72, pp. 248-254,
1976.
[62] J. A. Khan, S. Xiang and L. Tong, “Crystal Structure of Human Nicotinamide Riboside
Kinase,” Structure, vol. 15, pp. 1005-1013, 2007.
[63] R. Graeff and H. C. Lee, “A novel cycling assay for nicotinic acid-adenine dinucleotide
phosphate with nanomolar sensitivity,” Biochem. J., no. 367, pp. 163-168, 2002.
[64] W. Junge and E. Heymann, “Characterization of the isoenzymes of pig-liver esterase,” Eur. J.
biochem, no. 95, pp. 519-525, 1979.
[65] C. P. Landowski, P. L. Lorenzi, S. Xuequin and G. L. Amidon, “Nucleoside Ester Prodrug
Substrate Specificity of Liver Carboxylesterase,” The Journal of Pharmacology and
Experimental Therapeutics, no. 316, pp. 572-580, 2006.
[66] J. Yoshino, K. Mills, M. Yoon and S. Imai, “NICOTINAMIDE MONONUCLEOTIDE, A
KEY NAD+ INTERMEDIATE, TREATS THE PATHOPHYSIOLOGY OF DIET- AND
AGE-INDUCED DIABETES IN MICE,” CELL METABOL, vol. 14, pp. 528-36, 2011.
[67] P. Caton, J. Kieswich, M. Yagoob, M. Holness and M. Sugden, “Nicotinamide
mononucleotide protects against pro-inflammatory cytokine-mediated impairment of mouse
islet function.,” Diabetologia, vol. 54, no. 12, pp. 3083-92, 2011.
[68] P. Wang, T. Xu, Y. Guan, D. Su, G. Fan and C. Miao, “PERIVASCULAR ADIPOSE
TISSUE-DERIVED VISFATIN IS A VASCULAR SMOOTH MUSCLE CELL GROWTH
FACTOR: ROLE OF NICOTINAMIDE MONONUCLEOTIDE,” CARDIOVASCULAR
RESEARCH, vol. 81, pp. 370-80, 2009.
[69] P. Wang, T. Xu, Y. Guan, W. Tian, B. Viollet, Y. Rui, Q. Zhai, D. Su and C. Miao,
“NICOTINAMIDE PHOSPHORIBOSYLTRANSFERASE PROTECTS AGAINST
ISCHEMIC STROKE THROUGH SIRT1-DEPENDENT ADENOSINE
MONOPHOSPHATE-ACTIVATED KINASE PATHWAY,” ANN. NEUROL., vol. 69, no. 2,
pp. 360-74, 2011.
[70] J. Revollo, A. Korner, K. Mills, A. Satoh, T. Wang, A. Garten, B. Dasqupta, Y. Sasaki, C.
Wolberger, R. Townsend, J. Milbrandt, W. Kiess and S. Imai, “Nampt/PBEF/Visfatin
regulates insulin secretion in beta cells as a systemic NAD biosynthetic enzyme.,” Cell
69
Metab., vol. 6, no. 5, pp. 363-75, 2007.
[71] K. Yamada, N. Hara, T. Shibata, H. Osago and M. Tsuchiya, “The simultaneous measurement
of nicotinamide adenine dinucleotide and related compounds by liquid
chromatography/electrospray ionization tandem mass spectrometry.,” Anal. Biochem., vol.
352, no. 2, pp. 282-5, 2006.
[72] L. Formentini, F. Moroni and A. Chiarugi, “Detection and pharmacological modulation of
nicotinamide mononucleotide (NMN) in vitro and in vivo.,” Biochem. Pharmacol., vol. 77,
no. 10, pp. 1612-20, 2009.
70