METABOLISMO DEL
GLICOGENO
1
2
DESTINI DEL GLUCOSIO 6-FOSFATO
3
RAPPRESENTAZIONE SCHEMATICA DEL METABOLISMO DEL GLUCOSIO
4
Glucosio-6-fosfatasi
Omeostasi del glicogeno
•
Nell'organismo animale, il glicogeno è una riserva di carboidrati
•
Non è possibile conservare all'interno delle cellule il glucosio in forma libera.
•
Glicogeno epatico: serve a mantenere costante il livello ematico di glucosio
•
Glicogeno dei muscoli: è una riserva energetica (le cellule del muscolo non
5
possiedono l'enzima glucosio-6-fosfatasi)
Struttura del glicogeno
Formula molecolare
M.E.
Rappresentazione
schematica
6
DEMOLIZIONE DEL
GLICOGENO
7
•La
degradazione
del
glicogeno avviene, ad opera
della glicogeno fosforilasi
•L’enzima rimuove residui di
glucosio dalle estremità non
riducenti del polisaccaride
mediante fosforolisi.
•Parte
dell’energia
del
legame
glicosidico
viene
conservata
nell’estere
fosforico del glucosio 1fosfato
•L’enzima
lavora sino a
raggiungere la quarta unità
di glucosio prima di una
ramificazione.
8
L’enzima deramificante
•È
una α (1,4) glicosiltransferasi
e una α (1,6) glicosidasi; le 2
attività sono localizzate in 2 siti
separati della stessa proteina
•In una prima fase, l’enzima stacca
(attività transferasica) un blocco
di tre residui dalla ramificazione ad
una estremità non riducente vicina,
con formazione di un legame (a 1->
4) glicosidico.
Il residuo coinvolto nella formazione della ramificazione (legame a 1 -> 6)
viene rilasciato direttamente come glucosio libero (attività
glucosidasica).
9
La fosfoglucomutasi
•La
demolizione
del
glicogeno ad opera della
glicogeno
fosforilasi
produce G1P, che viene
trasformato in G6P dalla
fosfoglucomutasi
•La
fosfoglucomutasi
richiede glucosio 1,6bisfosfato come cofattore
•Il G6P può entrare nella
glicolisi oppure nella via
dei pentosi fosfati
•Nel fegato la glucosio 6 fosfatasi idrolizza il G6P in glucosio che
può così essere esportato ad altri organi
10
Rimodellamento del glicogeno
• I legami α 1,4-glicosidici
vengono scissi su ciascun
ramo dalla fosforilasi
• La transferasi sposta
un blocco di tre residui
glicosidici da un ramo
etremo all’ altro
• L’ α-1,6-glucosidasi
rimuove i residui in
modo da lasciare una
molecola lineare con
tutti i legami α-1,4glicosidici suscettibili
ad ulteriori scissioni
11
•Catalizza la fosforolisi del glicogeno a
La glicogeno fosforilasi
glucosio-1-Pi
•È regolata sia da interazioni allosteriche
che da modificazioni covalenti
•Utilizza il piridossal-5’-fosfato come
cofattore
•Non può staccare residui oltre 5 unità da
un punto di ramificazione
Disegno schematico
Struttura ai raggi X della glicogeno fosforilasi
12
1) Glicogeno fosforilasi : catalizza la fosforolisi del
glicogeno
Glicogeno + Pi
(n residui)
glicogeno + G 1P
(n - 1 residui)
2) Enzima deramificante del glicogeno
3) Fosfoglucomutasi
13
GLICOGENO FOSFORILASI
• è un omodimero
• ciascun sito catalitico
comprende un gruppo
piridossalfosfato ( PLP)
• il PLP è legato alla
lisina 680 dell’enzima
14
La fosforilasi a è fosforilata sulla serina 14 di ciascuna subunità che favorisce la
struttura dello stato R più attivo
15
• Sia la fosforilasi a sia la fosforilasi b
esistono come equilibri tra uno stato R
più attivo e uno stato T meno attivo
• La fosforilasi b di solito è inattiva
poiché l’equlibrio favorisce lo stato T
• La fosforilasi a è di solito attiva
poiché l’equilibrio favorisce lo stato R
16
Regolazione allosterica della fosforilasi muscolare
• Una bassa, carica
energetica, rappresentata da
elevate concentrazioni di
AMP favorisce la transizione
allo stato R
17
REGOLAZIONE ALLOSTERICA DELLA FOSFORILASI
EPATICA
• Il legame del glucosio alla
fosforilasi a sposta l’ equilibrio
verso lo stato T e inattiva
l’enzima
• Il glicogeno non viene
mobilizzato quando il
glucosio è già abbondante
18
Sintesi del glicogeno
19
Sintesi del glicogeno
La catena del polisaccaride
viene allungata (a partire da
una catena preformata di
almeno 8 residui) dall’enzima
glicogeno sintasi.
Il glucosio attivato sotto forma
di UDP-glucosio viene trasferito
all’estremità non riducente di
una catena polisaccaridica, con
formazione di un legame α(1-4)
glicosidico.
20
Sintesi dell’UDP-glucosio
•La
formazione di uno
zucchero attivato legato a
nucleotidi avviene mediante
una reazione di condensazione di un NTP con uno
zucchero fosforilato.
•La
carica
negativa
dell’ossigeno
serve
da
nucleofilo per un attacco sul
gruppo fosforico interno del
nucleoside trifosfato e libera
PPi.
•L’equilibrio
della reazione
viene
complessivamente
spinto
verso
destra
dall’idrolisi del pirofosfato
da parte della pirofosfatasi
inorganica.
• L’ UDP-glucosio è una forma attivata di
glucosio così come l’ATP e l’acetil CoA sono
forme attivate dell’ortofosfato e dell’acetato21
La glicogeno sintasi
•Catalizza l’aggiunta di unità di glucosio (a partire dall’UDPG) ad una
catena di glicogeno di almeno 8 residui
•Il primo step è catalizzato dalla tirosina glicosiltransferasi, che
attacca un residuo di glucosio all’OH di una Tyr della glicogenina
22
•La
proteina glicogenina (332 amminoacidi )
innesca la sintesi del glicogeno legando al
gruppo –OH di un residuo di Tyr del
polipeptide un residuo di glucosio.
AUTOGLICOSILA
•La
reazione utilizza UDP-glucosio e avviene
grazie all’attività autocatalitica della proteintirosina glicosiltransferasica.
•All’estremità
non riducente di questa prima
unità glucosidica legata alla glicogenina
vengono aggiunti altri 7 residui di glucosio ad
opera
della glicogenina associata
alla
glicogeno sintasi
•A
questo punto, è possibile l’azione diretta
della glicogeno sintasi che si dissocia dalla
glicogenina e sintetizza una catena lineare.
•Non
appena il numero di residui aggiunti lo
permette, abbiamo l’azione dell’enzima
ramificante che aggiunge una seconda
estremità non riducente sulla quale la
23
glicogeno sintasi può lavorare e così via.
Sezione trasversale di una molecola del glicogeno
GLICOGENINA
24
La reazione catalizzata della glicogeno sintasi.
25
L’enzima ramificante
(amilo (1,4
1,6 transglicosilasi)
1
•Le
ramificazioni
si
formano grazie all’azione
dell’enzima ramificante.
• L’enzima
lavora
trasferendo un segmento
terminale di 6-7 residui
glucosidici dall’e-stremità
non riducente di una
catena
poliglucosidica
composta da almeno 11
residui, al gruppo –OH del
carbonio C-6 di un residuo
di glucosio della stessa
catena o di un’altra catena
ma localizzato in un punto
più interno.
26
Ramificazione del glicogeno ad opera dell’enzima ramificante
L’enzima ramificante
(amilo (1,4
1,6 transglicosilasi)
2
•L’azione
combinata
di
glicogeno
sintasi
e
dell’enzima
ramificante
permette la formazione di
un polisaccaride a ramificazione crescente che
permette una crescita (e
una demolizione) molto più
rapida di un polisaccaride
lineare.
27
Ramificazione del glicogeno ad opera dell’enzima ramificante
Vie opposte di sintesi e di degradazione del glicogeno
28
Controllo del metabolismo
del glicogeno
29
Regolazione della sintesi e della degradazione
del glicogeno
•
Sintesi e degradazione del glicogeno sono coregolate in modo da
non funzionare simultaneamente
•
La regolazione comporta sia un controllo allosterico diretto sugli
enzimi
•
Sia un controllo ormonale mediato da una modificazione
covalente degli enzimi coinvolti
•
La glicogeno fosforilasi è attivata da AMP e inibita da ATP e G6P
•
La glicogeno sintasi è attivata dal G6P
30
A) Controllo allosterico diretto
della glicogeno fosforilasi e della glicogeno sintasi
B) Modificazione covalente
della glicogeno fosforilasi e della glicogeno sintasi
C) Effetti ormonali sul metabolismo del glicogeno
31
Controllo allosterico
[ATP]
[G6P]
[AMP]
Glicogeno fosforilasi +
Glicogeno sintasi -
[ATP]
[G6P]
Glicogeno fosforilasi –
Glicogeno sintasi +
32
Il sistema enzimatico di
interconversione della
glicogeno fosforilasi
33
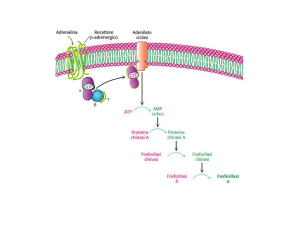
Protein chinasi A dipendente da AMPc
.Attivazione
della proteina chinasi dipendente dal cAMP.
• Le due subunità regolatrici
(subunità R) del complesso
R2C2 sono legate da due ponti
disolfuro.
• Il legame di due molecole di
AMPc a ogni subunità R
provoca la flessione di ogni
subunità R nella regione
cerniera e la liberazione delle
due subunità catalitiche (C).
34
•La fosforilasi b ( inattiva)
si trova nella
configurazione T
La fosforilasi a (attiva) è
principalmente nello stato
R
Controllo dell’attività della glicogeno fosforilasi
35
36
La calmodulina è una proteina
ubiquitaria che lega il Ca 2+ e che
partecipa a numerosi processi
cellulari di tipo regolatorio
37
STRUTTURA DELLA CALMODULINA
• E’ una proteina di 148 amminoacidi , altamente ben conservata
• Possiede due domini globulari strutturalmente simili collegati da
una struttura ad α-elica che descrive sette giri
• Ognuno dei due domini globulari contiene due siti di legame ad
alta affinità per il Ca 2+
• Ognuno di questi siti di legame per il Ca 2+ è formato da un
motivo strutturale elica-ansa-elica conosciuto come mano EF
• Il legame del Ca 2+ ad uno dei due domini della CaM
induce in quel dominio un cambiamento conformazionale
tale, da esporre una superficie idrofobica ricca di
Metionina. Questa zona , a sua volta, si lega con alta
affinità al dominio della CaM che lega la subunità γ della
fosforilasi chinasi
38
Subuità G che lega il
glicogeno
MECCANISMO DI
REGOLAZIONE DELLA
FOSFOPROTEINA
FOSFATASI
• Sono specifiche per la
rimozione del fosfato da
residui di serina
• La fosforilazione della
subunità G da parte della
proteina chinasi A causa il
rilascio della subunità
catalitica libera, che in
questa forma è meno attiva
1
4
2
3
Inibitore-1
5
39
Regolazione nel muscolo della fosfoproteina fosfatasi
Effetti antagonistici dell’insulina e dell’adrenalina
40
R2C2
proteina chinasi
cAMP-dipendente
(inattiva)
2C
proteina chinasi
cAMP-dipendente
(attiva)
cAMP
ADP
ATP
P P
(α β γ δ)4
(α β γ δ)4
fosforilasi
chinasi b
fosforilasi
chinasi a
Pi
SISTEMA DI
FOSFORILAZIONE
+ R2 (cAMP)4
altre
chinasi
H2O
ATP
ADP
glicogeno
fosforilasi b
glicogeno
fosforilasi a
Pi
ATP
P
H2O
ADP
glicogeno
sintasi a
P
glicogeno
sintasi b
Pi
H2O
P
inibitore della
fosfoproteina
fosfatasi-1 a
fosfoproteina
fosfatasi-1
(attiva)
fosfoproteina
fosfatasi-1
(inattiva)
ATP
ADP
inibitore della
fosfoproteina
fosfatasi-1 b
inibitore della
fosfoproteina
fosfatasi-1 a
Pi
SISTEMA DI
DEFOSFORILAZIONE
P
H2O
41
GLUCOSIO
42
GLUCAGONE
43
ADRENALINA
44
ACETILCOLINA
45
INSULINA
46
Il
bilancio netto tra la sintesi del glicogeno e la sua demolizione
viene controllato dai livelli ormonali di insulina e glucagone.
Questi,
regolando i livelli di cAMP (il secondo messaggero
intracellulare), determinano i rapporti tra le forma attive di glicogeno
sintasi e glicogeno fosforilasi.
Gli stessi ormoni regolano anche i livelli di F2,6BP e dunque il bilancio
tra glicolisi e gluconeogenesi.
L’adrenalina o epinefrina ha effetti simili a quelli del glucagone ma il
tessuto bersaglio di questo ormone è tipicamente il muscolo, mentre il
glucagone agisce essenzialmente a livello del fegato.
47
SCHEMA RIASSUNTIVO
Glucagone
Stimola la gluconeogenesi e la demolizione del glicogeno
a livello epatico
Insulina
Stimola la glicolisi e la glicogenosintesi a livello epatico
Adrenalina
Stimola la demolizione del glicogeno (fegato e muscolo) e
la gluconeogenesi (fegato)
48
Controllo ormonale del metabolismo del glicogeno
Glucosio 6-fosfato
49
Malattie del metabolismo del glicogeno
oTipo I: deficienza di glucosio 6 fosfatasi (Malattia di Von Gierke)
oTipo II: deficienza di a-1,4 glucosidasi (Malattia di Pompe)
oTipo III: deficienza di amilo-1,6-glucosidasi (Enz. deramificante; Malattia di Cori)
oTipo IV: deficienza di amilo-1.4---1,6-transglicosilasi (Enzima ramificante)
oTipo V: deficienza della fosforilasi muscolare (Malattia di Mc Ardle)
oTipo VI: deficienza della fosforilasi epatica (Malattia di Hers)
oTipo VII: deficienza di fosfofruttochinasi nel muscolo (Malattia di Tarui)
oTipo VIII: deficienza di fosforilasi chinasi legata al cromosoma X
oTipo IX: deficienza di fosforilasi chinasi
oTipo “0”: deficienza di glicogeno sintasi epatica
50
Tipo I : Morbo di Von Gierke
•La deficienza congenita dell’enzima glucosio 6 fosfatasi, che
idrolizza il glucosio 6 fosfato in glucosio) provoca:
• Abnorme accumulo di glicogeno nel fegato e nei reni.
•Grave ipoglicemia per mancato rilascio del glucosio dal fegato
(ipoglicemia insensibile alla somministrazione di glucagone e
adrenalina, ormoni che stimolano la glicogenolisi).
•Diminuito rilascio di insulina in seguito all’ipoglicemia.
•Elevata mobilizzazione degli acidi grassi per alterazione ormonale;
aumento dei NEFA e dei corpi chetonici.
•Incapacità del fegato di convertire il lattato in glucosio
(gluconeogenesi)
•Accumulo di acido lattico nel sangue che induce decalcificazione
delle ossa (osteoporosi).
51
Tipo II: deficienza di a-1,4 glucosidasi (Malattia di Pompe)
•Gli organi maggiormente colpiti sono cuore, fegato, muscolo
scheletrico (cardiomegalia e debolezza muscolare).
•Accumulo del glicogeno nei lisosomi dove la fosforilasi non ha accesso.
•La morte sopravviene dopo circa 6 mesi di vita.
52
Tipo III: deficienza di amilo-1,6-glucosidasi (Enz. deramificante; Malattia di Cori)
•Accumulo di glicogeno nel fegato, muscolo, cuore, eritrociti e leucociti.
•Epatosplenomegalia. Debolezza muscolare.
•Ipoglicemia a digiuno e mancata risposta al glucagone (risposta glicemica).
•Alcuni muoino nei primi anni di vita, altri raggiungono l’età adulta.
53
Tipo IV: deficienza di amilo-1.4---1,6-transglicosilasi (Enzima ramificante)
•La deficienza dell’enzima ramificante determina l’accumulo di glicogeno
anomalo, più solubile del normale che induce lesioni strutturali e funzionali.
•Epatosplenomegalia seguita da cirrosi.
•Alterazioni al cuore e ai muscoli scheletrici.
•La sopravvivenza è limitata ai primi anni di vita.
54
Glicogenosi tipo V: deficienza della fosforilasi muscolare (Malattia di Mc Ardle)
•Elevato contenuto di glicogeno nei muscoli, incapaci di utilizzarlo.
•La fosforilasi epatica è normale ed i pazienti non sono ipoglicemici.
•Presenti crampi muscolari e debolezza.
55