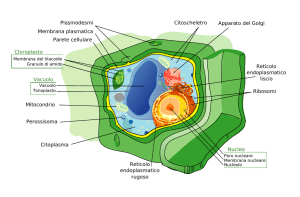
Molti organuli cellulari sono in comunicazione tramite un sistema di membrane interne Il sistema di membrane interne è un insieme di organuli circondati da membrane che lavorano insieme nel sintetizzare, immagazzinare e distribuire i prodotti cellulari (molecole importanti quali, per esempio, lipidi e proteine). I vari organuli del sistema di membrane interne sono interconnessi strutturalmente e funzionalmente. Reticolo endoplasmatico
ruvido
Vescicola di trasporto
proveniente
dal reticolo endoplasmatico
Vescicola di trasporto proveniente
dall’apparato di Golgi
Membrana plasmatica
Nucleo
Vacuolo
Lisososma
Reticolo endoplasmatico liscio
Membrana nucleare
Apparato di Golgi
Il nucleo è il centro di controllo della cellula Il nucleo è separato dal citoplasma tramite la membrana nucleare. Il nucleo è il centro di Cromatina
controllo genetico della Nucleolo
cellula eucariotica Poro
perché contiene il DNA che dirige tutte le attività cellulari. Nucleo
Membrana nucleare
a doppio strato
Reticolo
endoplasmatico
ruvido
Ribosomi
Il reticolo endoplasmatico ruvido dà origine a
membrane e proteine
Il reticolo endoplasmatico ruvido (RER):
- Presenta ribosomi adesi alla membrana
- Serve ad ampliare l’estensione del sistema di
membrane
- Serve per la sintesi delle proteine e assemblare le
proteine destinate a essere secrete dalla cellula.
I ribosomi sulla superficie del reticolo endoplasmatico
ruvido producono proteine che sono secrete dalla
cellula, inserite nelle membrane o trasportate in
vescicole ad altri organuli.
Vescicola di trasporto
4
che si stacca
Ribosoma
Vescicola di trasporto
con all’interno una
glicoproteina
3
Catena
glucidica
1
2
Polipeptide
Figura 4.8
Glicoproteina
Reticolo
endoplasmatico
Il reticolo endoplasmatico liscio svolge molteplici funzioni Il reticolo endoplasmatico liscio (REL):
• demolisce le tossine e i farmaci nelle cellule del fegato; • immagazzina e rilascia ioni calcio nelle cellule muscolari. Reticolo
endoplasmatico
liscio
Reticolo
endoplasmatico
ruvido
Involucro
nucleare
Ribosomi
Reticolo endoplasmatico liscio Reticolo endoplasmatico ruvido
TEM 45 000×
• sintetizza i lipidi (acidi grassi, fosfolipidi, steroidi); L’apparato di Golgi rielabora, seleziona e trasporta i prodotti cellulari L’apparato di Golgi è composto da sacchetti appiattiti impilati uno sull’altro che ricevono e modiGicano i prodotti del reticolo endoplasmatico e li trasportano ad altri organuli o sulla superGicie della cellula (per essere espulsi). Vescicola di trasporto
proveniente dal
reticolo
Apparato
di Golgi
Apparato di Golgi
Nuova vescicola
in formazione
Lato «di uscita»
dell’apparato
di Golgi
Vescicola
di trasporto
prodotta
dall’apparato di Golgi
TEM 130 000×
Lato «d’ingresso»
dell’apparato di Golgi
MITOCONDRI Mitos = Gilamento Chondros = granulo Forma a bastoncello, granulare. Diametro = 0.5 m Lunghezza = 1-­‐4 m Gino a 7 m. molto permeabile
(porine)
Sono degli apparati biochimici che recuperano l’energia contenuta negli alimenti (attraverso il ciclo di Krebs e la catena respiratoria) e mediante la fosforilazione la trasformano nel legame fosforico ad alta energia dell’ATP. Sono presenti in tutte le cellule a respirazione aereobica eccettuati i batteri nei quali gli enzimi respiratori sono localizzati nella membrana plasmatica. Delimitati da due membrane Le membrana interna si i n t r o G l e t t e f o r m a n d o numerose invaginazioni: le creste I mitocondri sono organuli semiautonomi e la maggior p a r t e d e l l e p r o t e i n e m i t o c o n d r i a l i s o n o codiGicate da geni nucleari e sintetizzate da ribosomi citoplasmatici. DNA mitocondriale Circolare a due catene polinucleotidiche La quantità di DNA contenuta in un mitocondrio varia a seconda della specie. I ribosomi sono del tipo batterico 70S. Sicuramente codiGicano la porzione idrofobica dell’ATPasi e la citocromo-­‐ossidasi. Respirazione:
Demolizione delle molecole organiche
all’interno delle cellule accompagnata
dal rilascio di energia utilizzabile
Glucosio (C6H12O6) Il catabolismo di molte sostanze ricche di energia inizia con la loro conversione in qualcuno degli intermedi della via di demolizione del glucosio. Negli uomini Dieta: saccarosio, lattosio, amido (saccarasi, lattasi, amilasi) Demolizione del glicogeno Nelle piante Amido e saccarosio La degradazione dell’amido è catalizzata da tre enzimi: 1) α-­‐amilasi: enzima idrolitico. Attacca i legami 1,4 sia d e l l ’ a m i l o s i o c h e d e l l ’ a m i l o p e c t i n a : s i l i b e r a n o progressivamente frammenti di circa 10 unità di glucosio che sono le destrine. Per ultimo dall’amilosio si genera il maltosio (disaccaride con 2 molecole di glucosio). Non attacca i legami 1,6 dell’amilopectina. 2) β-­‐amilasi: enzima idrolitico. Attacca l’amilosio solo nelle terminazioni non riducenti per cui verranno prodotte molecole di b maltosio (idrolizzato dalla maltasi intestinale). 3) Amido fosforilasi: enzima fosforolitico (amido + H2PO4-­‐ glucosio-­‐1-­‐P). La scissione dell’amido avviene per incorporazione del fosfato. La respirazione è un processo di ossido-riduzione perchè
comporta il trasferimento di elettroni da molecole donatrici verso
molecole che li accettano
RESPIRAZIONE
C6H12O6 + 6 O2
6 CO2 + 6 H2O + Energia E’ una reazione esoergonica (ΔG=-­‐686 kcal/mol, cioè si liberano circa 686.000 calorie per mole di glucosio ossidato) e si formano 36 molecole di ATP. Il glucosio viene completamente ossidato a CO2 mentre l’ossigeno viene utilizzato come accettore Ginale di elettroni e viene ridotto ad H2O. La respirazione è un processo suddiviso in tre tappe: 1) Glicolisi 2) Ciclo di Krebs 3) Catena di trasporto degli elettroni GLICOLISI (rottura dello zucchero): via di Embden-­‐Meyerhof (1930). Per ogni molecola di glucosio demolita e ossidata si producono due molecole di piruvato 6 C6H12O6
2 CH3COCOOH + 4 H+ + 4 e-­‐ (glucosio) (acido piruvico) Avviene nel citoplasma e non necessita di O2; è una via completamente anaerobica. Durante la glicolisi la molecola di glucosio ricca di energia viene scissa in due molecole di acido piruvico più povere di energia: Una mole di glucosio contiene 686.000 calorie, due moli di acido piruvico 547.000 calorie. Pertanto durante la glicolisi l’energia liberata è di circa 139.000 calorie per mole di glucosio. ➫ il glucosio viene ossidato senza la partecipazione di ossigeno libero. ➫ i protoni e gli elettroni ceduti dal glucosio vengono assunti dal NAD
NADH2 (2 molecole). ➫ durante la glicolisi avviene la fosforilazione a livello di substrato (2 molecole di ATP). GLICOLISI 1) Prima fase o fase di “investimento energetico” comprende reazioni endoergoniche che necessitano di ATP. Il glucosio è una molecola relativamente stabile non facile da rompere; in due reazioni di fosforilazione due gruppi fosfato sono trasferiti dall’ATP allo zucchero. Lo zucchero fosforilato che si forma (fruttosio 1,6 difosfato) è meno stabile del glucosio e si può scindere per via enzimatica. 2) La seconda fase comprende reazioni esoergoniche che permettono di produrre ATP (fosforilazione a livello del substrato) e NADH La fase preparatoria, che richiede l’impiego di 2 molecole di ATP termina con
la scissione di 1 molecola di zucchero a 6 atomi di carbonio in 2 molecole a 3
atomi di carbonio
La fase conclusiva dà luogo ad un guadagno energetico di 4 molecole di ATP
e 2 molecole di NADH
Virtualmente tutte le cellule possiedono la capacità di estrarre l’energia dal glucosio ossidandolo a piruvato generando due molecole di ATP per molecola di glucosio. Ciò che succede dopo, dipende dalla disponibilità di ossigeno; il catabolismo a valle del piruvato in condizioni aereobie è diverso da quello in condizioni anaerobie. In presenza di ossigeno il piruvato è ossidato completamente ad anidride carbonica nel processo di respirazione Respirazione: Glusso di elettroni attraverso una membrana a partire da coenzimi ridotti Gino ad un accettore di elettroni, di solito accompagnato dalla produzione di ATP. Per molti organismi (uomini e animali) l’accettore ultimo di e-­‐ è l’O2, la forma ridotta dell’accettore è l’H 2 O ed il processo nel suo insieme è detto respirazione aerobia. Matrice
Matrice
mitocondriale mitocondriale Membrana interna
Tutti gli enzimi del ciclo di krebs sono localizzati in soluzione nella matrice,
tranne uno associato alla membrana interna insieme ai componenti della
catena di trasporto degli elettroni
La membrana esterna del mitocondrio è permeabile a molte piccole molecole
La membrana interna permette il passaggio di alcune molecole (piruvato, l’ATP,
l’ADP) e ostacola il passaggio di altre molecole (ioni H+)
I tappa: decarbossilazione ossidativa (in presenza di Coenzima A e di NAD+) dell’acido piruvico: CH3COCOO-­‐ + CoA-­‐SH + NAD+
CH3-­‐CO-­‐SCoA + CO2 + NADH + H+ Il piruvato proveniente dalla glicolisi passa nella matrice mitocondriale e viene ossidato in acetato attivo o acetil-­‐CoA. Questa reazione è catalizzata dalla piruvato deidrogenasi 3
2
2
1
1
CICLO DI KREBS o ciclo degli acidi tricarbossilici o ciclo dell’acido citrico. I due atomi di carbonio del gruppo acetilico che entrano nel ciclo di krebs si combinano con un composto a 4 atomi di carbonio (ossalacetato) dando luogo ad un composto a 6 atomi di carbonio (citrato). Nel corso del ciclo due dei sei atomi di carbonio del citrato vengono ossidati ad anidride carbonica rigenerando l’ossalacetato Bilancio globale del ciclo di Krebs: I due atomi di carbonio che entrano come gruppo acetilico dell’acetil CoA, sono ossidati ad anidride carbonica Gli atomi di idrogeno sono trasferiti ai coenzimi NAD+ e FAD. L’ossidazione dell’acetil-­‐CoA consente la formazione di un legame energetico: l’ATP (fosforilazione a livello di substrato). NADH e CO2 proveniente dal piruvato
COMPLESSIVAMENTE: Glucosio + 10 NAD+ + 2 FAD + 4 ADP + 4 Pi
6 CO2 + 10 NADH + 2 FADH2 + 4 ATP NADH: 2 dalla glicolisi + 2 dalla decarbossilazione ossidativa del piruvato + 6 dal ciclo di Krebs per due CoA che entrano Gli elettroni del NADH e del
FADH2 si trovano ad alto un
livello energetico
Questi elettroni ad alta energia
vengono trasportati gradualmente
a un livello energetico più basso
lungo la catena di trasporto degli
elettroni
Ogni trasportatore di elettroni si
trova ad un livello energetico
leggermente più basso
Alla fine della catena gli
elettroni vengono accettati
dall’ossigeno che
combinandosi con i protoni
(ioni idrogeno) forma acqua
TRASPORTATORI: Flavoproteine Ferro-­‐zolfo proteine (Fe+2/Fe+3) Citocromi (che acquistano elettroni con il loro atomo di ferro) Citocromi contenenti rame (Cu2+/Cu+) Chinone (coenzima Q). Tutti i trasportatori sono capaci di accettare e donare uno o due elettroni e sono contenuti nella membrana interna del mitocondrio. Il coenzima Q non fa parte di alcun complesso respiratorio e non è associato a proteine, ma esiste sotto forma di “pool” nella porzione interna, apolare, della membrana mitocondriale interna (o plasmatica nei procarioti). Può accettare o donare uno o due elettroni. Prende un protone insieme ad ogni elettrone, quindi trasporta l’equivalente di un atomo di idrogeno Le proteine trasportatrici di elettroni sono organizzate in quattro complessi multiproteici in serie (I, II, III e IV) che contengono i trasportatori di elettroni e gli enzimi necessari a catalizzare il trasferimento di elettroni da un trasportatore al successivo. Durante il Glusso degli elettroni in discesa lungo la catena di t r a s p o r t o d e g l i elettroni, i protoni vengono pompati a t t r a v e r s o i t r e complessi proteici dalla matrice allo spazio intermembrane Modello dell’accoppiamento chemiosmotico: la differenza di concentrazione in ioni H+ che si stabilisce da una parte e dall’altra della membrana interna provoca la comparsa di un gradiente elettrochimico (un gradiente di pH in quanto lo spazio intermembranoso diventa più acido della matrice e un gradiente di cariche elettriche corrispondente ad un potenziale di membrana di circa 150 mV) I protoni si muovono secondo
il gradiente elettrochimico
dallo spazio intermembrana
verso la matrice e fluiscono
attraverso l’ATP sintasi.
Il rientro dei protoni attraverso
il complesso ATP sintasi libera
l’energia necessaria per la
sintesi di ATP da ADP e
fosfato
(infatti gli elettroni del FAD passano solo
attraverso due complessi)
Efficienza del processo globale di
respirazione= circa 40%
(denitrificazione) (Paracoccus denitrificans)
Solo nel caso che l’ossigeno operi da accettore terminale di elettroni durante l’ossidazione di un substrato si attua una vera respirazione aereobia. Nel caso che l’ossigeno sia sostituito da altri composti (per es. NO3-­‐, N2O, SO42-­‐) si parla di respirazione anaerobia (solo alcune specie batteriche come Paracoccus denitri/icans). Se una cellula eucariote e procariote si trova nell’impossibilità di effettuare la respirazione aerobia o anaerobia, la glicolisi sfocia nella fermentazione: in questo processo la degradazione degli zuccheri non sarà completa ma si arresterà a livello di composti organici ancora ricchi di energia (alcol etilico o acido lattico). Inoltre non vi è alcun accettore di e-­‐ esterno. Durante la glicolisi una molecola di NAD+ viene ridotta a NADH per ogni piruvato prodotto; per evitare che NAD+ si esaurisca la cellula deve trovare il modo per rigenerarlo. In presenza di O2 il NADH è riossidato attraverso il trasferimento dei suoi elettroni e protoni all’ossigeno. In condizioni anaerobie i due elettroni (e un protone) del NADH sono trasferiti al carbonio 2 del piruvato Le due vie di fermentazione più comuni usano il piruvato c o m e a c c e t t o r e d i e -­‐ t r a s f o r m a n d o l o i n l a t t a t o (FERMENTAZIONE LATTICA) o in CO 2 ed etanolo (FERMENTAZIONE ALCOLICA). IN ASSENZA DI OSSIGENO:
Perché non vi è un’ossidazione totale!
FERMENTAZIONE ALCOLICA: via anaerobica operata da Saccaromiceti, funghi e batteri. Le cellule di lievito presenti sull’uva sembrano polvere e si mescolano con il mosto quando l’uva viene pigiata. I lieviti presenti in questa miscela, in condizioni anaerobiche, trasformano il glucosio presente nel succo d’uva in etanolo. Nella fermentazione alcolica viene liberata CO 2 che nei liquidi in fermentazione (es. mosto che si trasforma in vino) produce un caratteristico ribollio. 2 ADP + 2 Pi
2 ATP
O—
C O
Glucose
C O
Glycolysis
CH3
2 pyruvate
2 NAD+ + 2 H+
H
2 NADH
2 CO2
H
H
C OH
C O
CH3
CH3
2 H+
2 ethanol (secreted from the cell)
(b) Production of ethanol
2 acetaldehyde
Saccharomyces ellipsoidens (per il vino) Saccharomyces cerevisiae (per la birra). Anche le piante superiori in assenza di O2 fermentano (es. radici immerse in acqua). I lieviti hanno una tolleranza limitata per l’alcol e, quando questo raggiunge la concentrazione del 12%, le cellule di lievito muoiono e la fermentazione cessa. Risvolti applicativi ed economici della fermentazione:
Tutte le bevande alcoliche si ottengono mediante processi fermentativi: Il rhum dallo zucchero di canna Il vino dallo zucchero d’uva La birra e il whisky dal malto d’orzo Il sakè dal riso. Nella paniGicazione, la fase della lievitazione si avvale della fermentazione operata da un Saccharomyces (CO 2 resta nell’impasto e etanolo si allontana durante la cottura). La produzione dello yogurt impiega un batterio: Lactobacillus bulgaricus che opera la fermentazione lattica.