DIVERSITÀ METABOLICA
i procarioti, nel loro insieme,
possono utilizzare:
quasi tutti i
composti naturali
Qualche composto
xenobiotico
Sono quindi importanti per i
cicli biogeochimici
Per il biorisanamento
Per acquisire nutrienti e trasformarli in macromolecole organiche, i
procarioti possono usare molte e diverse strategie
zuccheri
Proteine
minerali
vitamine
oligoelementi
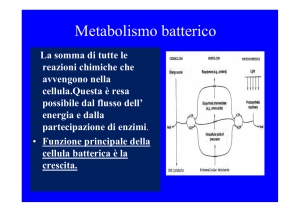
Il metabolismo energetico è l’insieme delle reazioni
biochimiche che si verificano in una cellula vivente
L’energia liberata in una reazione, che diventa disponibile per
svolgere un lavoro utile alla cellula batterica è “energia libera”: un
fattore determinante nel metabolismo microbico
∆“G”
substrato
prodotto
La variazione di energia libera (Δ G) in una reazione, dipende dalla
concentrazione di substrato e prodotto e dal pH
Una reazione, con Δ G negativo (che rilascia energia libera) è
spontanea e si definisce ESOERGONICA
ΔG-
substrato
prodotto
se il ΔG di una reazione è positivo, la reazione non è spontanea
(tenderà a verificarsi invece la reazione contraria) e si dice
ENDOERGONICA
ΔG+
substrato
prodotto
Gli enzimi accoppiano una reazione esoergonica e una reazione endoergonica
abbassano l’energia di attivazione di una
reazione e facilitano il metabolismo
substrato
prodotto
substrato
prodotto
L’energia di attivazione è l’energia libera richiesta da una molecola
per entrare in una determinata reazione
La velocità della reazione è inversamente proporzionale
all’energia di attivazione richiesta
VELOCE
LENTA
L’uso dell’energia chimica coinvolge reazioni redox
Si ossida
Si riduce
idrogeno
-
H
H
+
ossigeno
+
H
O
H
-
Libera energia
assorbe energia
Esotermica
(esoergonica)
endotermica
(endoergonica)
Si ossida
-
Donatore di elettroni
FONTE DI ENERGIA
elettroni
Si riduce
+
Accettore di elettroni
La tendenza a cedere o acquisire eè il potenziale redox (E0)
L’energia liberata in una reazione è proporzionale
alla differenza di E0 tra i composti coinvolti nella reazione
E0 positivo: ossidanti
TENDONO A RIDURSI
OSSIDANDO ALTRI COMPOSTI
E0 negativo: riducenti
TENDONO A OSSIDARSI E
RIDURRE ALTRI COMPOSTIV
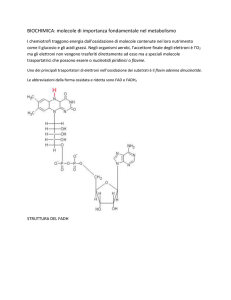
Nei processi di produzione di energia intervengono i trasportatori
intermedi di elettroni
NAD
NADP
Coenzimi
LIBERI
e-
FMN-FAD
CITOCROMI
CHINONI
FISSATI
CHE PERMETTONO IL TRASFERIMENTO DEGLI ELETTRONI DAL
DONATORE PRIMARIO ALL’ACCETTORE TERMINALE
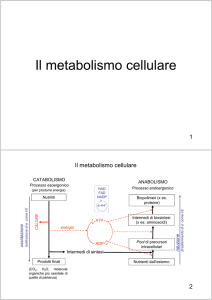
Il metabolismo ha due facce
anabolismo
catabolismo
Anabolismo
Nutrienti
Macromolecole e altri
composti cellulari
Energia per
le biosintesi
Energia
(luminosa o chimica)
Catabolismo
Energia per Mobilità,
trasporto nutrienti
Prodotti di scarto
Catabolismo
Fermentazione
(Anaerobia)
Respirazione
Aerobia-Anaerobia
Nello studiare le strategie metaboliche microbiche si tiene conto della fonte
di energia, del composto che cede gli elettroni e della fonte di carbonio
Foto-
energia : direttamente dalla luce
chemo-
energia : da reazioni chimiche, non
direttamente dalla luce
organo-
donatore di elettroni: organico
lito-
donatore di elettroni: inorganico
auto-
Carbonio (fonte di) = CO2
etero-
Carbonio (fonte di) composti organici
Le categorie si combinano definendo gruppi
microbici metabolicamente differenti
Tutte le combinazioni possono esistere, ma
le più comuni sono quattro
I MICRORGANISMI
Fonte di carbonio: CO2 inorganico
AUTOTROFI
Sintetizzano il proprio nutrimento
riducendo CO2
CO2
Fonte di carbonio: molecole organiche
ETEROTROFI
Usano come nutrienti
molecole organiche
AUTOTROFI
Fonte di energia: luce solare
FOTOAUTOTROFI
CO2
Fonte di energia: legami chimici
CHEMOAUTOTROFI
CO2
ETEROTROFI
Fonte di energia: luce solare
FOTOETEROTROFI
Fonte di energia: legami chimici
CHEMOETEROTROFI
microrganismi che possono vivere in assenza di composti organici
Fotoautotrofi
Cianobatteri
Chemoautotrofi.
batteri che ossidano lo zolfo
rossi
sulfurei
archaebatteri metanogeni
verdi
sulfurei
Fotoeterotrofi
Di questo gruppo fanno parte
gli archibatteri alofili
I batteri non sulfurei
verdi
rossi
hanno bisogno di composti organici come fonte di carbonio
cellulare ma non per l’energia
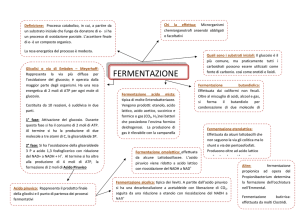
FERMENTAZIONE
NAD
NADP
reazione redox tra due molecole organiche
con stato di ossidazione simile
Composto organico
(donatore)
Composto organico ossidato
(accettore)
l’ossidazione del
substrato è incompleta
l’energia non si libera
completamente
Composto organico ridotto
(prodotto)
La resa energetica è bassa
L’energia liberata viene immagazzinata come ATP
prodotto durante il catabolismo
dei composti organici fermentati
Composto organico
(donatore)
Composto organico ossidato
(accettore)
energia
ADP
ATP
Composto organico ridotto
(prodotto)
Con un processo di fosforilazione a livello del substrato
~P
fermentazione del glucosio:
due passi
1) il glucosio viene
ossidato e si forma
piruvato (glicolisi)
Acido piruvico
Glucosio
elettroni
NAD
NADH
e‐
Gli elettroni vengono accettati dal coenzima NAD, che si riduce
I batteri impiegano vie diverse
per svolgere questo processo
Via di Embden-Meyerhoff (EM)
(eucarioti e batteri enterici)
Via dei pentoso-fosfati
(tipica dei batteri acidolattici)
Via di Entner-Douderoff (ED)
(Pseudomonas; Archibatteri)
fermentazione del glucosio:
Glucosio
GLICOLISI
ossidazione del glucosio
+ riduzione coenzimi
Acido piruvico
(INTERMEDIO)
NAD
NADH
elettroni
piruvato
Prodotti finali
Acido piruvico
(INTERMEDIO)
FERMENTAZIONI
riduzione del piruvato
+ ossidazione coenzimi
PRODOTTI
FINALI
I°
Glucosio
II°
G‐6P
3‐GADP
PIRUVATO
Omolattica
Acido lattico
III°
Eterolattica:
Acido lattico
Etanolo
CO2
Acidi misti:
Acido acetico
Acido succinico
Etanolo
CO2 , H2
alcolica:
Alcol etilico + CO2
propionica:
Acido propionico
Acido acetico
CO2
butanediolica:
Butanolo
CO2
Butirrico‐butilica:
Butanolo
Isopropanolo
Acetone
CO2
Fermentazione etanolica (Saccharomyces cerevisiae)
Fermentazioni acidolattiche
(Lactobacillus, Streptococcus, Lactococcus)
Omolattica (produzione di formaggio, yoghurt..):
ottiene acido lattico come unico prodotto
Eterolattica (produzione dei crauti): ottiene acido
lattico, etanolo e CO2
fermentazione propionica (substrato acido lattico)
tipica di Propionobacterium,
importante nella produzione di Emmenthal.
fermentazione ad acidi misti
caratteristica della famiglia delle Enterobacteriaceae, per la
quale può avere valore diagnostico
Il piruvato è convertito in molti prodotti, specialmente acido acetico, acido
lattico ed etanolo
(la composizione esatta dei vari prodotti è specie-specifica)
Fermentazione butanolica (acido butirrico) alcune specie di Clostridium.
I prodotti sono acido butirrico (odore di burro rancido), n-butanolo e acetone
(importanti prodotti industriali)
Fermentazione ad aminoacidi misti: vengono deaminati e decarbossilati
(ridotti e ossidati simultaneamente) diversi aminoacidi.
Molto importante per conferire aromi particolari ad alcuni vini e formaggi.
A differenza di quanto accade nella fermentazione, il processo di respirazione
può sfruttare un accettore di e- che non fa parte della catena (esterno)
e-
ACCETTORE
ESTERNO
DONATORE
O2
(aerobia)
Fe3-/NO3/SO4
(anaerobia)
la presenza di un accettore di elettroni “esterno” comporta notevoli
vantaggi
ACCETTORE
ESTERNO
L’ossidazione è completa CorgÆCO2
La netta differenza redox tra donatore iniziale e accettore terminale
permette la sintesi di un numero maggiore di molecole di ATP
IL METABOLISMO RESPIRATORIO SI SVOLGE IN TRE FASI:
GLICOLISI
Attraverso le stesse
vie usate dalla
fermentazione
CICLO DI KREBS
FOSFORILAZIONE
OSSIDATIVA
Ma con una resa
energetica superiore
Ciclo di Krebs o dell’acido tricarbossilico (TCA)
Ottiene 3 molecole di CO2 attraverso l’ ossidazione
completa dell’ acido piruvico
Trasportatori di
elettroni:NAD/FAD
FOSFORILAZIONE OSSIDATIVA
generazione di ATP durante l’ossidazione dei coenzimi ridotti
(NADH, FADH) attraverso una catena di trasporto di eIl trasporto genera un gradiente di protoni
attraverso la membrana
il gradiente di protoni permette la sintesi di ATP
Ciclo di Krebs o dell’acido tricarbossilico (TCA)
Ottiene 3 molecole di CO2 attraverso l’ ossidazione
completa dell’ acido piruvico
CO2
PIRUVATO
Ac-CoA
-
citrato
ossalacetato
CO2
CO2
Trasportatori di
elettroni:NAD/FAD
FOSFORILAZIONE OSSIDATIVA
generazione di ATP durante l’ossidazione dei coenzimi ridotti
(NADH, FADH) attraverso una catena di trasporto di e-
Il trasporto genera un gradiente di protoni
attraverso la membrana
il gradiente di protoni permette la sintesi di ATP
i trasportatori sono disposti secondo il potenziale redox
NADH
NAD
NDH
FP
H2O
FeS
QH2
CITs
CO
Gli elettroni sono trasferiti fino
all’accettore terminale, che si riduce
½O2
i trasportatori che trasferiscono anche protoni si alternano con
quelli che trasferiscono solo elettroni
Gli ioni ossidrile negativi restano all’interno
2H2O
2OH‐
2H2O
2OH‐
2H+
2H+
2H+
I protoni che non vengono trasferiti sono
espulsi e restano all’esterno della cellula
2H2O
½ O2
La separazione delle cariche sui due versanti
della membrana crea una forza
protomotrice che guida la sintesi di ATP
acido
‐
+
‐
alcalino
‐
+
‐
‐
+
‐ ‐ ‐ ‐ ‐ ‐ ‐ ‐ ‐ ‐ ‐ ‐
+
+ +
+
+
+
‐‐
‐
+
‐
+
+ ‐‐
+
‐
+
+
+
‐ ‐ ‐ ‐ ‐ ‐
‐ ‐
+
‐
+
+
+ ‐
+
+
+
+
+ + + + + + + + + +
Il trasporto di elettroni, con il gradiente di protoni
che si forma ENERGIZZA la membrana
H+
H+
H+
H+
H+
energia
I protoni vengono incanalati attraverso una proteina
F1F0 sintasi (ATPasi di membrana) che ne usa l’energia
per la sintesi di ATP
ATP
ADP
~P
F0 è una proteina idrofoba che attraversa la membrana e
porta gli ioni a F1, che aggetta sul versante citoplasmatico
F1 catalizza due tipi di reazione
ATPÆADP + Pi
ADP + Pi Æ ATP
[ATP] gradient
e
A seconda della [ATP] e dall’entità del
gradiente attraverso la membrana
Ingresso H+/
ADPÆATP
[ATP] gradient
e
uscita H+/ ATPÆADP
RESPIRAZIONE AEROBIA E ANAEROBIA
HANNO IN COMUNE ALCUNI COMPONENTI DEL TRASPORTO
e‐
e‐
NO3/SO4
Fe3MA ACCETTORI TERMINALI DIVERSI
O2
La produzione di energia è maggiore nella respirazione
La variazione di energia libera infatti è profondamente diversa
Glucosio Æ CO2 - H2O , ΔG = -686 kcal/mole (respirazione)
Glucosio Æ acido lattico, Δ G = -58 kcal/mole (fermentazione)
R
F
Nella respirazione vengono prodotte 38 molecole di ATP per molecola di glucosio
Nella fermentazione solo due
LA FERMENTAZIONE PERO’ GARANTISCE LA VITA
DI MOLTISSIMI MICRORGANISMI
? COME MAI ?
La Risposta è nel concetto di efficienza
quanta dell’energia rilasciata viene
effettivamente immagazzinata?
La respirazione ha un’efficienza del 40% circa; la
fermentazione ha un’efficienza di oltre il 50%
Chemoeterotrofi
dipendono completamente dai composti organici.
Saprofiti
(Materia morta)
e patogeni
(Materia organica viva)
si trovano in questo gruppo
i microrganismi fotosintetici convertono
l’energia luminosa in energia chimica
ENERGIA
luminosa
Molecole
complesse
Anabolismo
Energia chimica
Utilizzata per le sintesi della cellula batterica
batteri rossi sulfurei
(Chromatiaceae)
Cianobatteri
batteri verdi non sulfurei
(Chloroflexaceae)
batteri verdi sulfurei
(chlorobiaceae)
batteri rossi non sulfurei
(Rhodobiaceae)
PER UTILIZZARE L’ENERGIA LUMINOSA
SONO NECESSARIE LE CLOROFILLE
I Cianobatteri
Possiedono una
Clorofilla a
Come quella degli
eucarioti
I BATTERI ROSSI E VERDI
R2
R1
R3
hanno Batterioclorofille
R7
con picchi di assorbimento
diversi
R4
R5
R6
I PIGMENTI ACCESSORI
carotenoidi, ficobiliproteine;
ficoeritrina e ficocianina
catturano la luce da 470 a 630
(giallo-verde)
dove le clorofille non arrivano
Trasferiscono l’energia alla clorofilla
ampliano lo spettro utilizzabile e proteggono i batteri da una
eccessiva intensita’ luminosa
CLOROFILLE E PIGMENTI ACCESSORI
SONO RIUNITI IN “ ANTENNE”
antenna
(circa 300 molecole di clorofilla)
energia luminosa
centro di reazione
(trasporto fotosintetico di e-)
I BATTERI VERDI E ROSSI SULFUREI (ANAEROBI) SVOLGONO
UNA FOTOFOSFORILAZIONE ANOSSIGENICA E CICLICA
Chromatiaceae
Chlorobiaceae
le loro batterioclorofille assorbono luce
nell’intervallo 800-1100 nm
Il donatore di elettroni, per la fissazione di CO2 è
H2S (o altri composti ridotti dello zolfo)
La clorofilla, colpita dalla luce,
cambia stato
Emette un elettrone ad alta energia
(ECCITONE)
l’eccitone genera una forza
protomotrice passando per una
catena di trasportatori
‐1
840*
Fs 870*
Fd
Q
0
CIT
CIT
+0,5
870
CIT
+0,25
840
E0CIT
E torna alla batterioclorofilla che
riprende lo stato originale
Nelle Chromatiaceae, il potenziale dei chinoni (~0) è maggiore di quello dell’NAD+
(-0,32) che non può essere ridotto direttamente
‐1
Parte dell’energia ottenuta è quindi spesa
per rigenerare il potere riducente
indispensabile per fissare CO2
870*
NADH
0
Q CIT
+0,5
CIT
Solfuri,tiosolfati
Acido succinico (rossi non sulfurei)
E0 > NADP+/NADPH
Nelle Chlorobiaceae, P840* assume un valore di E0 più fortemente
negativo; la ferro-zolfo proteina, attraverso la ferredossina,
può ridurre direttamente NAD+.
‐1
L’energia si divide tra il pool di chinoni e la
ferredossina Æ ATP + NADH
840*
Fs Fd
Q
0
+0,25
CIT
E0CIT
NADH
Composti ridotti dello zolfo possono anche
donare e- direttamente al citocromo C553
H2S / S2O32-
TRASFERIMENTO DIRETTO
H2 ha E0 tanto basso (-0,42 volt) da ridurre direttamente NADP (-0,32)
le specie che possiedono idrogenasi
(catalizza l’ossidazione di H2 )
Possono trasferire elettroni direttamente a NAD+
Idrogenasi !
Microrganismi fotoetrotrofi
batteri rossi e verdi non sulfurei;
archeobatteri alofili (Halobacteraceae)
usano l’energia luminosa ma non fissano CO2 e
necessitano di fonti di carbonio organiche
O2
Luce
Assenza di O2
buio
Luce
fotosintesi
Respirazione
aerobia
buio
respirazione anaerobia/
fermentazione
nei batteri rossi non sulfurei (Rhodobiaceae)
l’ossigeno inibisce la sintesi delle ICM
In anaerobiosi è presente un sistema di membrane che si diramano dalla
IM e supportano i pigmenti fotosintetici neoformati
T0
LUCE + ANAEROBIOSI
T4h
La transizione da respirazione a fotosintesi è completa nel giro di circa quattro ore
le Halobacteraceae non usano clorofille
Sono archibatteri aerobi e ossidano il carbonio
organico usando O2 come accettore
Vivono in ambienti dove la concentrazione salina
ostacola la respirazione ma la luce è forte
L’energizzazione della membrana è operata dalla batteriorodopsina, connessa a
un retinale che ne separa due emicanali interni
Luce
retinale cambia conformazione
emicanali si uniscono
la batteriorodopsina
trasferisce H+ all’esterno
Fotofosforilazione non ciclica:
Cianobatteri, proclorofite
due fotosistemi in tandem: acqua come donatore di
elettroni e ossigeno come prodotto di scarto
‐1,2
0
PS-II con la
clorofilla “A” P680
P700
p680
+1,2
PS-I con la
clorofilla “A” (P700)
‐1,2
PS-I e PS-II producono energia e potere riducente usando acqua come
donatore di elettroni formando ossigeno come prodotto di scarto
0
FNR
2NADP+
P700
p680
+1,2
PS-II
genera ATP
PS-I, riduce
NADP+ a NADPH
2NADPH
Entrambe le clorofille emettono un eccitone
Quello emesso da
P680 reintegra P700
‐1,2
O2 + 4H+
0
2H2O
P680 (~1.3) può
sottrarre elettroni
all’acqua (+0,82)
+1,2
P700
p680
TRASPORTO INVERSO DI
ELETTRONI
(Rossi sulfurei e non sulfurei)
Termodinamicamente
SFAVOREVOLE
Attinge al potenziale di
membrana
batteriofeofitina
NAD+
Q
Cyt
c2
Cyt
bc1
e‐
Solfuri,tiosolfati
Acido succinico
(rossi non sulfurei)
PQ
P700
P870
E0 > NADP+/NADPH
MODIFICAZIONE DELLA
FOTOFOSFORILAZIONE CICLICA
(verdi sulfurei)
Fe-S
Q
Cyt
c553
PQ
Cyt
bc1
ATP
2e‐
P700
P840
NAD+
e‐ H2S / S2O32‐
La presenza/assenza di OSSIGENO nella produzione di
energia è il risultato di una lunga storia
L’ossigeno è divenuto parte integrante
dell’atmosfera nel paleo-medio proterozoico
O2
O2
O2
La fotosintesi ossigenica dei cianobatteri
produsse tanto O2 da causarne una presenza
stabile nell’atmosfera
La presenza di vaste stratificazioni di ossidi di ferro nei
sedimenti fossili ha permesso di datare questo avvenimento
Paleoproterozoico: O2 = 15%
O2 in
aumento
Anossico
Formazione microrganismi
della Terra anaerobi
4,5
4,0
3,5
Fotosintesi
ossigenica
3,0
2,5
Archeano:
O2 = 1%
Ricco di O2
Eucarioti
2,0
1,5
Animali
1,0
0,5
Presente
la respirazione aerobia si sviluppò in risposta all’aumentata tensione di O2
nell’atmosfera rappresentando un vantaggio come meccanismo di detossificazione e
di produzione di energia
I microrganismi “aerobi” che hanno
bisogno di ossigeno
Hanno dovuto anche evolvere strategie di difesa nei
confronti delle sue forme tossiche
3O
2
(ossigeno tripletto) è il normale stato di base dell’ossigeno
Durante molte reazioni foto o bio-chimiche, può
prodursi ossigeno singoletto (1O2 )
La forma singoletto ha una maggiore energia
e una grande reattività
Molto tossica
(provoca reazioni incontrollate)
1O
2
1O
2
1O
2
L’ossigeno singoletto è prodotto spesso in
ambienti fortemente illuminati
I microrganismi che vivono in ambienti dove sono esposti alla presenza di
ossigeno singoletto producono spesso pigmenti carotenoidi
ALTRE FORME TOSSICHE
Anione superossido
Perossido di idrogeno (H2O2 )
Radicale idrossile
O2-
H2O2
OH
.
Queste forme tossiche sono prodotte durante la riduzione
dell’ossigeno molecolare a H2O (respirazione)
O2
+
e-
O2-
+
e- +
H2O2
+
e-
.
+
e-
OH
O2-
H2O2
2H+
+
H+
+
OH
H+
.
H2O
H2O
La detossicazione è affidata a enzimi che eliminano
il perossido di idrogeno
catalasi
H2O2
H2O
O2
H2O2
H2O2
H2O
Le perossidasi hanno bisogno di cofattori riducenti
perossidasi
H2O2
H2O
NADH
H2O
H+
H2O2
NAD+
NADH
superossidodismutasi
O2-
O2-
H2O2
H+
O2
H+
O2-
Contro il prodotto più tossico, il radicale ossidrile (OH. ) non ci sono
enzimi ma la rimozione di H2O2 ne previene la formazione