TRASPORTO DI MEMBRANA
Movimento di soluti dentro e fuori dalla
cellula (attraverso la membrana cellulare o
plasmalemma) per l’acquisizione di nutrienti.
Movimento dei soluti all’interno della cellula
tra i vari compartimenti (attraverso le
varie membrane degli organelli) per lo
svolgimento dei processi metabolici.
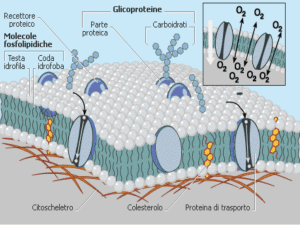
Le membrane cellulari
semipermeabili.
sono
membrane
Il trasporto passivo è un fenomeno spontaneo
che si compie con diminuzione di energia
libera (∆
∆G < 0).
Il trasporto attivo si compie con un aumento
di energia libera (∆
∆G > 0) e quindi per
verificarsi deve essere collegato a processi
esoergonici che forniscono energia.
Termodinamica del trasporto
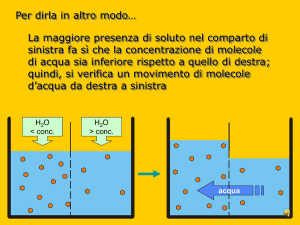
Soluto non carico. L’energia che si libera nel
trasferimento di una mole di soluto dallo
scomparto A allo scomparto B è data da:
∆G = RT ln [S]B/[S]A
se SA>SB il ∆G è negativo ed il movimento del
soluto da A verso B avviene spontaneamente
senza consumo di energia.
se SA<SB il ∆G è positivo e per trasferire il
soluto da A verso B occorre compiere un lavoro
con consumo di energia.
Soluto carico. Il trasferimento del soluto da A a
B determina, oltre ad una differenza di
concentrazione, anche lo sviluppo di una differenza
di carica attraverso la membrana. Poiché cariche
simili si respingono, il lavoro per spostare il soluto
da A verso B sarà anche funzione della differenza
di carica elettrica attraverso la membrana
(potenziale di membrana, ∆E).
L’energia libera implicata nel movimento (legata alla
differenza di carica) sarà data da:
∆G = nF ∆E
L’energia libera legata al movimento dei soluti
con carica è quindi determinata sia dal
gradiente di concentrazione (potenziale chimico)
che da quello elettrico (potenziale elettrico) ed
è data dalla somma delle due energie
(potenziale o gradiente elettrochimico):
∆G = RT ln [S]B/[S]A + nF ∆E
Il movimento dei soluti nella cellula sarà quindi
determinato dai versi e dall’entità dei due
gradienti.
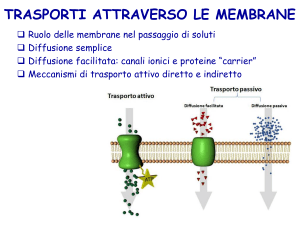
TRASPORTO PASSIVO
Qualsiasi membrana attraverso cui
possono passare alcuni soluti mentre
altri ne sono impediti viene detta
membrana semipermeabile.
La permeabilità dei soluti attraverso la
membrana è legata al
coefficiente di
ripartizione tra i lipidi e l’acqua del
soluto:
> coefficiente > permeabilità
Le membrane (doppi strati lipidici) sono in
genere impermeabili alle molecole polari e
neutre di elevate dimensioni e agli ioni.
Gas come CO2 ed O2 permeano le membrane
molto rapidamente.
Acqua: trasporto tramite cambi di conformazione
delle code idrofobiche degli acidi grassi, carriers
(trasportatori di natura proteica), ma soprattutto
canali selettivi di costituiti da proteine integrali di
membrana (acquaporine).
Tipi di trasporto passivo
Permeazione
Diffusione semplice (permeazione)
E’ un processo spontaneo che avviene
secondo un gradiente di concentrazione (da
zone a più alta concentrazione a zone a più
bassa concentrazione) mediante il quale un
soluto tende a distribuirsi uniformemente
nella soluzione.
Diffusione semplice (canali)
Proteine transmembrana che funzionano da canali o
pori selettivi nella membrana.
Velocità di trasporto circa 106 volte maggiore che
nella diffusione facilitata.
Presenza di “cancelli” che aprono e chiudono il
canale in risposta a segnali esterni.
Il trasporto attraverso i canali è sempre passivo
ed è limitato principalmente agli ioni.
Diffusione facilitata
Presenza di proteine transmembrana specifiche
il cui compito è quello di facilitare il trasporto
frazionando l’energia di attivazione necessaria
tramite
una
via
alternativa
di
attraversamento.
facilitano
il
I
trasportatori
(carriers)
movimento solo ed esclusivamente nella
direzione termodinamicamente favorevole e
NON richiedono energia per la loro attività.
Come gli enzimi, i carriers presentano:
• incremento della velocità di trasporto;
• selettività
e
specificità
inibizione competitiva;
di
trasporto;
• saturazione cinetica.
Cinetica simile a quella descritta dalla relazione di
Michaelis-Menten.
KT (costante di trasporto) = concentrazione dello
ione o della molecola quando la velocità di trasporto
è la metà della velocità massima. Maggiore è la KT e
minore è l’affinità del carrier per il soluto.
Antagonismo = competizione degli ioni per lo
stesso sito di trasporto (carrier). L’aumento
della concentrazione di uno ione nella soluzione
deprime l’assorbimento di altri.
Sinergismo = la presenza di uno ione stimola
l’assorbimento di un altro ione tramite azione
induttiva
sull’attività
del
trasportatore
(attivazione) oppure accelerazione della velocità
di combinazione tra ione e trasportatore.
TRASPORTO ATTIVO
Le piante accumulano molecole neutre e ioni
contro un gradiente di concentrazione e/o di
carica elettrica consumando quindi energia.
Il
trasporto
attivo
cosiddetto
primario
(endoergonico) è accoppiato direttamente ad una
fonte di energia metabolica come l’idrolisi
dell’ATP, e le proteine di membrana responsabili
del trasporto attivo primario sono definite
‘pompe’.
Nella membrana plasmatica e nel tonoplasto il
principale ione trasportato elettrogenicamente
attraverso la membrana è lo ione H+ (H+-ATPasi).
Il
gradiente
protonico
(gradiente
elettrochimico) rappresenta l’energia libera
accumulata in seguito al movimento degli H+
effettuato dalla pompa H+-ATPasi.
Questa energia potenziale generata dalla
traslocazione dei protoni è utilizzata per
effettuare il trasporto di numerose altri
soluti contro i loro gradienti elettrochimici
nel trasporto attivo secondario (anch’esso
endoergonico).
trasporto attivo primario
trasporto attivo secondario
∆G = RT ln [S]B/[S]A + nF ∆E
Per ristabilire l’equilibrio di concentrazione e di carica, gli H+
devono rientrare nel compartimento di origine. Se il risultato
del trasporto è un cambiamento nel numero di cariche si parla
di trasporto elettrogenico. In caso contrario, di trasporto
elettroneutro.
Il potenziale di membrana ∆E (valore negativo, circa -200 mV) è
generato principalmente dall’efflusso di ioni K+ ([K+]est < [K+]int) e
dall’estrusione di H+ (attività delle pompe H+-ATPasi), oltre che
dalla dissociazione di soluti cellulari.
cariche nette negative dovute
alla dissociazione di proteine
e acidi organici
canale del K+
H+-ATPasi