TRASPORTI ATTRAVERSO LE MEMBRANE
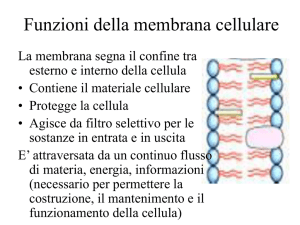
Ruolo delle membrane nel passaggio di soluti
Diffusione semplice
Diffusione facilitata: canali ionici e proteine “carrier”
Meccanismi di trasporto attivo diretto e indiretto
LA DIFFUSIONE SEMPLICE
Il
doppio
strato
fosfolipidico rappresenta
una barriera selettiva al
passaggio di soluti
Fattori che influenzano
la diffusione semplice:
Grandezza molecola
Temperatura
Carica molecola
Gradiente concentrazione
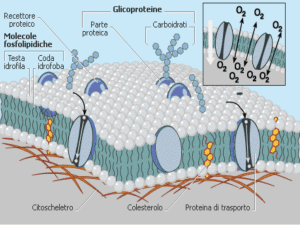
Le proteine di membrana si specializzano per le
funzioni di trasporto
LA DIFFUSIONE FACILITATA
Proteina canale
Proteina carrier
LE PROTEINE DI TRASPORTO
Le proteine trasportatrici (carrier) accelerano una reazione
che è già termodinamicamente favorita, legando il soluto da
trasportare: tale legame determina un cambiamento
conformazionale della proteina trasportatrice, che nella nuova
conformazione perde affinità per il soluto, “scaricandolo” dal
lato opposto della membrana.
Nella diffusione facilitata:
La velocità di diffusione è molto più alta rispetto alla
diffusione semplice
La proteina trasportatrice è specifica per il soluto da
trasportare
Il numero delle proteine trasportatrici è limitante per
l’efficienza e la velocità del trasporto
Tutti i carrier sono
occupati
SATURAZIONE
Modello di
funzionamento di
una proteina
trasportatrice
Il gradiente di glucosio viene
mantenuto poiché, una volta
dentro la cellula, esso è
rapidamente demolito
La concentrazione di glucosio nel sangue deve rimanere entro
certi limiti ed è regolata dagli ormoni (insulina, glucagone)
L’insulina segnala che la glicemia
è più alta del normale, quindi il
glucosio
in
eccesso
viene
assorbito
dal
sangue
e
convertito
in
glicogeno
e
trigliceridi.
Il glucagone segnala
che la glicemia è più
bassa del normale e
stimola la demolizione
del
glicogeno,
la
gluconeogenesi e la
degradazione
degli
acidi grassi
Il numero di proteine trasportatrici per un
determinato soluto può essere regolato
I CANALI IONICI
A differenza dei carrier, i canali ionici non si legano al
soluto di cui facilitano il passaggio secondo il gradiente
(di concentrazione ed elettrico)
I canali ionici non hanno sempre un’alta specificità per
un unico tipo di ione
La velocità di diffusione attraverso i canali ionici è molto
elevata (milioni di ioni al secondo per ogni canale)
I canali ionici non sono sempre “aperti”
Lo stato “aperto” o “chiuso” del canale dipende da fattori
diversi per ogni tipo di canale (canale voltaggio-dipendente,
canale ligando-dipendente) che non hanno nulla a che fare
con gli ioni in questione.
Canale ligando-dipendente
IL TRASPORTO ATTIVO
È direzionale
Contro gradiente di concentrazione (richiede energia)
Tre meccanismi possibili (uniporto, simporto, antiporto)
Diretto (o primario) e indiretto (o secondario) a seconda che ci
sia richiesta diretta di ATP
IL TRASPORTO ATTIVO DIRETTO
C’è consumo
diretto di ATP
I gradienti di concentrazione di alcuni ioni sono
generati e mantenuti da meccanismi di trasporto
attivo
Pompa Na+/K+: esempio di antiporto
Importante per mantenimento dell’equilibrio osmotico !
(Es. uabaina inibisce pompa scoppio cellula)
La fosforilazione fa
cambiare la
conformazione
Nelle cellule eucariotiche la concentrazione citosolica
del Ca2+ è mantenuta bassa grazie a pompe Ca2+-ATPsi
presenti sulla membrana plasmatica.
La Ca2+-ATPasi del muscolo pompa Ca2+ dal citosol nel
reticolo sarcoplasmatico (immagazzinamento Ca2+ che si
libera nel momento della contrazione muscolare).
La membrana plasmatica di cellule che secernono acido (stomaco)
contiene H+-ATPasi.
H+-ATPasi pompano protoni attraverso le membrane dei lisosomi
Le cellule della parete gastrica acidificano il contenuto del lume
dello stomaco mantenendo il pH del citosol neutro.
IL TRASPORTO ATTIVO
INDIRETTO
Simporti o antiporti
Energia per il trasporto recuperata dal
movimento di ioni secondo gradiente
generato da trasporto attivo diretto
Proteine di simporto legate a Na+ portano aminoacidi e
glucosio dentro le cellule contro gradiente sfruttando
proprio il passaggio del Na+
Trasporto attraverso gli epiteli: le cellule
dell’epitelio intestinale sono fortemente polarizzate
Il movimento del glucosio attraverso le cellule polarizzate di
un epitelio richiede diverse proteine di trasporto
ENDOCITOSI,
ESOCITOSI
e
TRAFFICO DI MEMBRANE
ENDOCITOSI - ESOCITOSI
ENDOCITOSI
FAGOCITOSI
FAGOCITOSI
ENDOCITOSI IN FASE FLUIDA O PINOCITOSI
La pinocitosi è costitutiva e aspecifica, ossia la cellula
introduce piccole gocce di matrice extracellulare in
maniera indifferenziata. Questo è possibile perché il
materiale in questione è presente disciolto in soluzione
acquosa.
ENDOCITOSI mediata da recettori è regolata e
specifica. La cellula riconosce il suo substrato
mediante proteine di membrana (recettori).
Questi recettori posti in lievi depressioni sulla
membrana (fossette rivestite) sono in grado di
legare
il
materiale
da
introdurre.
In questo modo la cellula riesce ad assumere anche
sostanze
poco
presenti
nell’ambiente
extracellulare.
L’endocitosi mediata da recettori generalmente
avviene mediante “fossette rivestite” di clatrina
Il
rivestimento
di
clatrina
conferisce
resistenza e stabilità
alle vescicole, ma viene
perso
appena
la
vescicola entra nella
cellula.
Una fossetta rivestita di clatrina che sporge dalla
faccia citosolica della membrana plasmatica
Il recettore delle LDL lega
e internalizza particelle
contenenti
colesterolo
(complessato
nelle
lipoproteine LDL poichè è
insolubile in acqua)
Ipercolesterolemia
familiare: deficit di
recettori per LDL
livello ematico di
colesterolo elevato
ESOCITOSI
Endocitosi ed
esocitosi
sono due vie opposte nel
traffico di membrane dal
Golgi trans alla membrana
e dalla membrana ai
lisosomi