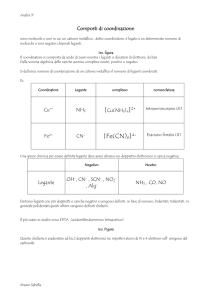
Struttura Quaternaria
Due o più subunità unite in modo non covalente
Esempi
-2 subunità identiche:
-alcool deidrogenasi
-2 o più subunità diverse:
-Hb (a2b2)
-RNA polimerasi (a2bb’s)
-piruvato deidrogenasi (t24(p2)12(f2)12)
-Strutture polimeriche a dimensioni finali variabili
-microtubuli (anbn)
1. Come si dispongono le subunità
• Simmetria: nessun legame apparente con la funzionalità
•
Il numero di subunità di un aggregato proteico è (quasi sempre –
vedi punto 3) indipendente dal ruolo funzionale
2. Quali forze stabilizzano la struttura
quaternaria
•
•
•
forze di non-legame
legami H
interazioni elettrostatiche
3. Quale ruolo assolve
Mioglobina
Emoglobina
Singola subunità Hb ~ Mioglobina
F8: His prossimale legata al Fe
Eme
E7: His distale
Fe
Prevalentemente a-elica 8 tratti A-H connessi da tratti di random coil
MGLSDGEWQL
LEKFDKFKHL
GHHEAEIKPL
PGDFGADAQG
VLNVWGKVEA
KSEDEMKASE
AQSHATKHKI
AMNKALELFR
DIPGHGQEVL
DLKKHGATVL
PVKYLEFISE
KDMASNYKEL
IRLFKGHPET
TALGGILKKK
CIIQVLQSKH
GFQG
Solo Ca
Blow-up eme
Eme: gruppo prostetico
Fe - Protoporfirina IX
Coordinazione Fe
•
4 N della porfirina
•
1 N istidina prossimale (F8)
•
sito libero per il legame con O2
Fe n.a. 26
Due stati di ossidazione
•
Ferroso Fe2+ può legare O2
• Ferrico Fe3+ non può legare O2 (meta-emoglobina denaturazione)
Fe n.a. 26
1s
2
2
2s
2
2
2p
6
6
3s
2
2
3p
6
6
3d
6
5
Shell incomplete
Fe2+
Fe3+
Il concetto di cooperatività
L'affinità per il legante (O2 nell'emoglobina) modificata da un
cambiamento conformazionale
modulato dal legame di una (o più di una) subunità con il legante
Hb scatto conformazionale (della struttura quaternaria) quando da due a
tre delle quattro subunità legano legante
aumento (~ 300 volte) dell'affinità delle subunità non ancora legate
curva di saturazione con l’ O2
Confronto tra Hb (a più subunità) e Mb (singola subunità)
Y: saturazione frazionaria = rapporto tra concentrazione di proteina (o
subunità, nel caso dell’Hb) legata con O2 e concentrazione totale.
pO2: pressione parziale di ossigeno
Mb + O2
Y
MbO2
MbO2 Mb
k
Mb 1
1
1
MbO2 kpO2
Y
MbO2
MbO2
MbO2
MbO2
kpO2
Mb
kpO2
Y
1 kpO2
Iperbole equilatera
Y
k pO2 n
1 k pO2 n
Forma generale n = 1 non cooperativo (singola subunità)
n > 1 cooperatività
X
Hb: la pendenza della curva
di saturazione cambia al
variare di pO2
Y
k pO2 n
1 k pO2 n
Il modello di Monod-Wymann-Changeux (MWC)
Ipotesi:
1. promoteri identici occupano posizioni strutturalmente equivalenti nella
proteina, che equivale a dire che la proteina ha almeno un asse di simmetria
2. ciascun promotero ha almeno un sito per ciascuno dei diversi ligandi
(definizione di promotero);
3. Esistono almeno due stati conformazionali (strutture) reversibilmente
accessibili alla proteina per i quali deve valere:
i. in ciascuno degli stati la simmetria di cui al punto 1 è conservata;
ii. l’affinità per una dato legante può essere anche molto diversa nei due
stati
4.
l’affinità per uno specifico legante dipende dallo stato conformazionale
dell’intero enzima.
Nota: Interazione omotropica: l’affinità per un legante è influenzata da quanto legante
dello stesso tipo è già legato ( cooperatività).
Interazione etrotropica: il legame di un legante (al suo specifico sito di legame) ha
effetto sull’affinità di legame di un altro (es: effetto Bohr , affinità e pH)
Il caso dell’Hb.
# promoteri=n=4
2 stati: R (relaxed) e T (tense)
.
Per rispettare la condizione (3i) solo alcune combinazioni di
stati dei singoli promoteri sono permesse.
… e analoghe per lo stato tense
Dove kR
(kT) è la costante di dissociazione microscopica (reazione di
dissociazione del singolo legante dal singolo sito di legame) nello stato
relaxed (tense)
Nota: la costante di dissociazione microscopica è sempre la stessa indipendentemente
dallo stato di occupazione degli altri siti i 4 siti sono implicitamente ipotizzati
essere indipendenti.
Si possono quindi scrivere le seguenti reazioni di equilibrio
R0 + L
R1 + L
R2 + L
R3 + L
R1
R2
R3
R4
T0 + L
T1 + L
T2 + L
T3 + L
T1
T2
T3
T4
Ri (Ti ): specie microscopica con i leganti L legati nello stato R (T )
K1
M 0 L
M 1
K2
M 1 L
M 2
Kn
M n1 L
M n
Per una generica molecola, M, con n siti.
Per il generico i-esimo sito
e
M i 1
M i 2 L
K i 1
M i 1 L
Ki
M i
ovvero
che sostituita nella precedente
M i M i1 L
Ki
M i M i2 LL
K i K i 1
i
M i M 0 L K j
e infine
i
j 1
Ci sono
n,i
n!
i!(n i)!
modi distinti di disporre i leganti in n siti
da cui la seguente generica relazione tra costanti macroscopiche e microscopiche
Ki
n,i 1
n,i
n!
i!(n i)!
i!(n i)!
k
k
k
(i 1)!(n 1 i)! n!
(i 1)!(n 1 i)!
L’esempio dell’Hb
R e T siano i due stati conformazionali
kR kT (ipotesi 3.ii)
kR
kT
C
con C = costante >1
la costante di dissociazione microscopica è maggiore nello stato T ( il
legante preferisce stare legato alla forma R)
Dalla precedente relazione tra costanti macroscopiche e microscopiche
K1
n,i 1
n,i
kR
i!(n i)!
1!(4 1)!
kR
kR
kR
(i 1)!(n 1 i)!
0!(4 1 1)!
4
ed equivalentemente
2k R
3k R
K2
; K3
;
3
2
K 4 4k R
Dalle reazioni di equilibrio
[ R0 ][ L] k R
[ R1 ]
4
[ R ][ L] 2k R
K2 1
[ R2 ]
3
[ R ][ L] 3k R
K3 2
[ R3 ]
2
[ R ][ L]
K4 3
4k R
[ R4 ]
K1
E analoghe per lo stato T
Che utilizzando le definizioni
a
[T ]
[ L]
; Q 0
kR
[R 0 ]
diventano
[ R1 ] 4[ R0 ]a
[ R2 ] 6[ R0 ]a 2
dove [T0 ] e[R 0 ] sono le concentrazioni delle
[ R3 ] 4[ R0 ]a 3
subunità prive di leganti rispettivamente in T e R
[ R4 ] [ R0 ]a 4
Utilizzando la
Q
[ L]
4 4Q[ R0 ]Ca
kT
[T2 ] 6Q[ R0 ]C 2a 2
[T3 ] 4Q[ R0 ]C 3a 3
[T4 ] Q[ R0 ]C 4a 4
[T1 ] Q[ R0 ]
[T0 ]
[R 0 ]
si ottengono le relazioni equivalenti per T
La saturazione frazionaria sia data da
i[ Ri ] i i[Ti ]
i
YL
4i [ Ri ] i [Ti ]
Dove, al numeratore sono sommate le concentrazione totali di siti occupati con
un numero qualsiasi, i, di leganti in uno dei due stati conformazionali e al
numeratore la concentrazione totale.
Riscrivendo le varie sommatorie in funzione di a, C, [R0] e Q
i 4
[ R ] [ R ](1 4a 6a
i 0
i
0
2
4a 3 a 4 ) [ R0 ](1 a ) 4
i 4
4
[
T
]
Q
[
R
]
(
1
C
a
)
i
0
i 0
i 4
2
3
4
3
i
[
R
]
0
[
R
]
1
4
[
R
]
a
2
6
[
R
]
a
3
4
[
R
]
a
4
[
R
]
a
)
4
[
R
]
a
(
1
a
)
i
0
0
0
0
0
0
i 0
i4
3
i
[
T
]
4
Q
[
R
]
C
a
(
1
C
a
)
i
0
i 0
La saturazione frazionaria si può
scrivere per n=4
Se
a (1 a ) 3 QCa (1 Ca ) 3
YL
(1 a ) 4 Q(1 Ca ) 4
[T0 ] 0 (subunità indipendenti o singola subunità), anche Q 0 , quindi
a (1 a ) 3
a
YL
1a
(1 a ) 4
E per n qualsiasi
la curva di saturazione è un iperbole.
a (1 a ) n1 QCa (1 Ca ) n1
YL
(1 a ) n Q(1 Ca ) n
Andamento di YL in funzione della concentrazione di legante
• bassa concentrazione di legante libero
a
[ L]
1
kR
a (1 a )3
a
YL
(1 a ) 4 Q Q
YL
Retta di pendenza
[R ]
Q 1 0
[T0 ]
a
• dove
YL
1 a Q 1
a (1 a ) 3
Q
[T ]
[ L]
0
kR
[ R0 ]
a
3a 2 3a 3 a 4
Q
Q
Q
Q
YL varia rapidamente per piccole variazioni di a
[ L]
e quindi di [L]
kR
YL
a
• dove
[ L] [T0 ]
a Q
1
k R [ R0 ]
[L] è grande tutti i siti sono saturati con il legante
YL
YL 1
Si noti che la sigmoicità
a
[T0 ]
• cresce al crescere di Q
[ R0 ]
• decresce mentre
C
kR
1
kT
Le frecce in verde indicano il modello descritto da Perutz per l’Hb
M.F.Perutz “Mechanism of cooperativity and allosteric regulation in
proteins”, (1989) Quart.Rev.Biophys., 22, 139-236.
Effetto Bohr